纤毛鹅观草原变种和竖立鹅观草变种居群间核型变异研究
2023-12-25方忠艳朱明昆包俊浩庞菁璐张亚洲康厚扬吴丹丹
方忠艳 朱明昆 包俊浩 庞菁璐 张亚洲 康厚扬 吴丹丹
(四川农业大学小麦研究所,成都611130)
鹅 观 草 属(Roegneria)是 小 麦 族(Triticeae Dumortier)中最大属,该属包含异源四倍体和异源六倍体,基因组符号为StY 或StStY。现知全世界约有130 余种,广泛分布于北半球的温寒地带。我国约有70 余种,主要分布于西北、西南和华北地区[1]。纤 毛 鹅 观 草(Roegneria ciliaris,StY,2n=4x=28)是鹅观草属多年生异源四倍体物种,有纤毛鹅观草原变种(R.ciliarisvar.ciliaris)、阿穆尔鹅观草变种(R.ciliarisvar.amurensi)和竖立鹅观草变种(R.ciliarisvar.japonensis)3 个变种,在我国有广泛的分布[1-2]。纤毛鹅观草是优良的牧草植物,赣饲一号纤毛鹅观草是我国第一个选育并登记的纤毛鹅观草牧草新品种[3]。纤毛鹅观草是栽培小麦的三级基因资源库,在小麦遗传改良中具有重要价值,它具有抗赤霉病、白粉病、条锈病及抗非生物逆境等优良性状。前人通过纤毛鹅观草与普通小麦杂交,将含有抗病抗逆基因的染色体转入普通小麦,并创制鉴定出普通小麦—纤毛鹅观草二体异附加系等异染色体系种质资源[4-7]。然而,纤毛鹅观草存在3 个变种,变种在自然中存在不同来源的居群,这些变种之间和居群之间的遗传多样性和染色体水平的变异也尚不明了,使纤毛鹅观草的有效利用受到限制。
植物染色体核型分析是对植物染色体数量及结构变异、形态结构特征进行定量和定性的描述,是对物种亲缘关系与进化程度的表达的重要方法[8]。卢宝荣等[9]对纤毛鹅观草和竖立鹅观草的形态学比较、核型分析和染色体组进行分析,认为纤毛鹅观草和竖立鹅观草应为一个物种的两个变种,以纤毛鹅观草为原变种,而竖立鹅观草为纤毛鹅观草的一个变种。魏秀华等[10]对纤毛鹅观草和竖立鹅观草不同居群的染色体核型进行研究,表明两个材料核型特征有很大的相似性且两个材料的不同居群间存在丰富的遗传多样性。基因组荧光原位杂交(GISH,Genomic in situ hybridization)和荧光原位杂交(FISH,Fluorescence in situ hybridization)技术是识别小麦族物种染色体组和单条染色体的重要手段。Wang等[11]利用质粒pCbTaq4.14 DNA,在纤毛鹅观草的1St、2St、3St、7St 染色体上检测到FISH 信号。利用pTa794、pTa71、RcAfa 和(GAA)10,Kong 等[6]在一整套小麦-纤毛鹅观草异附加系中构建了纤毛鹅观草的14 条染色体的FISH 核型。程梦豪等[12]、Cheng 等[13]利用前期开发的St 基因组特异性探针oligo-2ScL-163,结合oligo-pAs1、(GAA)10、黎巴嫩拟鹅观草(St 基因组)DNA 进行FISH 和GISH 对纤毛鹅观草品系建立核型,通过统计不同品系FISH核型分布特征和分子标记分析,揭示了不同纤毛鹅观草品系染色体水平具有丰富的遗传多样性以及染色体的结构重排。但是,目前关于纤毛鹅观草种下水平的染色体FISH 核型研究尚未见报道。
鹅观草属的St 和Y 基因组亲缘关系较近,双色GISH 在区分仅含有鹅观草属物种中的St 和Y 染色体上仍有难度。Wang 等[14]筛选了新的St 基因组标记St2-80,用于区分小麦族中的St 和其他基因组,尤其是区分鹅观草属物种中的St 和Y 基因组。目前鹅观草属中Y 亚基因组的起源尚不清楚,因此专门针对鹅观草属的St 和Y 亚基因组开发的探针数量极为有限。Wu 等[15]利用纤毛鹅观草(StY)和St 基因组同源四倍体糙缘拟鹅观草进行比较基因组学分析,筛选开发了St 和Y 基因组富集的重复序列探针StY_34、StY_90、StY_93、StY_107,鉴定大鹅观草(R.grandis)的核型。因此可以利用这4个St 和Y 基因组重复序列探针结合St2-80 和45S rDNA 对纤毛鹅观草种下水平的染色体结构变异开展进一步研究。
本研究对纤毛鹅观草的纤毛鹅观草原变种和竖立鹅观草变种共15 份居群材料的染色体长短臂进行测量,计算染色体核型参数;同时利用StY_34、StY_90、StY_93、StY_107、45S rDNA、St2-80 6个重复序列探针,对15 种不同来源材料进行FISH核型分析,建立FISH 核型图和模式图,比较分析不同来源的纤毛鹅观草原变种和竖立鹅观草变种的染色体核型差异,为了解纤毛鹅观草以及鹅观草属野生材料染色体特征提供依据。
1 材料与方法
1.1 植物材料本试验包含纤毛鹅观草的2 个变种R.ciliarisvar.ciliaris和R.ciliarisvar.japonensis共15 份居群材料,物种名、材料编号、染色体数目和基因组组成、采集地列于表1。所有材料来自于四川农业大学小麦研究所,材料均种植于四川农业大学小麦研究所种质资源圃,凭证标本保存于四川农业大学小麦研究所标本室(SAUTI)。
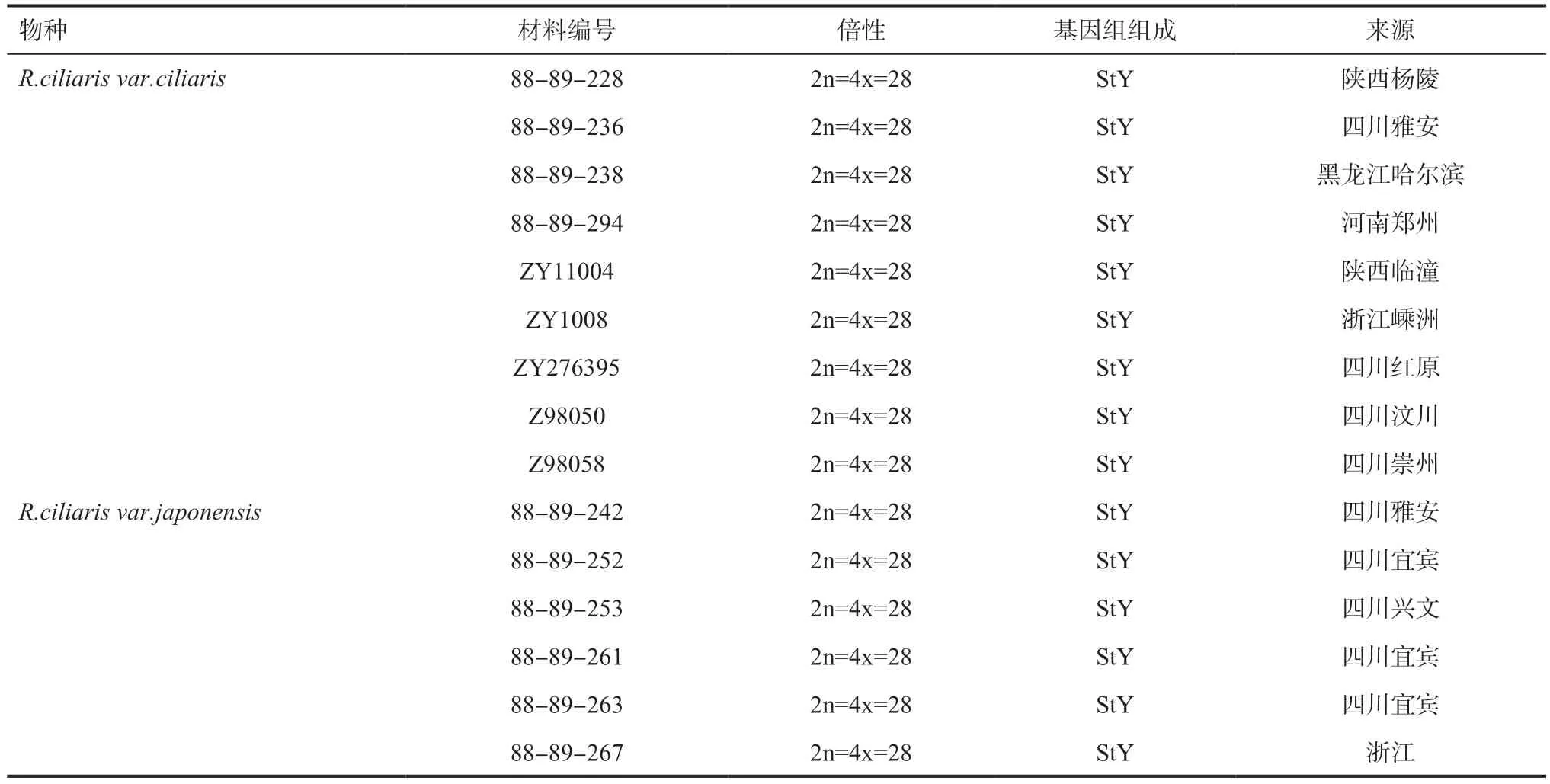
表1 供试材料
1.2 有丝分裂中期染色体制备和染色体核型分析染色体制片和染色体长度测量参照Wu 等[16]方法进行。其中核型公式、平均臂比、染色体长度比、臂比大于2 的比例参照李懋学等[8]的方法计算,核型不对称系数按Arano[17]的方法计算,分类标准依照Stebbins[18]的方法进行分类。
1.3 探针标记、FISH 和显微镜检查St2-80[14]、45S rDNA[19]、StY_90、StY_34、StY_107 和 StY_93[15]6 个探针用于鉴定纤毛鹅观草和竖立鹅观草染色体。以普通小麦(中国春)和纤毛鹅观草的基因组DNA 为模板,通过聚合酶链式反应分别扩增 45S rDNA 和St、Y 基因组富集型探针。 StY_93 探针用6-FAM(6-carboxy - fluorescein)标记(TAMRA;Sangon Biotech,中国上海),其余探针均由DNA 使用Atto 488 NT Labeling Kit(PP-305L-488) 和Atto 550 NT Labeling Kit(PP-305L-550)(Jena Bioscience,Jena,Germany)试剂盒将重复序列标记成探针。FISH 探针的杂交按照 Wu 等[15]的方法进行,略有改动。在18μL 杂交缓冲液中加不同色素荧光标记探针各1μL,制备总共20μL 的双色FISH杂交溶液。将20μL 杂交混合溶液置于90℃变性炉上变性10min。杂交混合液添加到载玻片上,用25mm×25mm 盖玻片覆盖,于80℃变性炉上变性90s,将处理好的载玻片在37℃的黑暗潮湿环境中培养6~8h。杂交后的载玻片在55℃的2×SSC 中洗涤20min,用75%、85%和100%梯度酒精脱水各1min,干燥后在黑暗环境中滴加10~15μL 的4’,6-二脒基-2-苯基吲哚(DAPI),盖上盖玻片,即可在BX63 荧光显微镜下观察。荧光观察拍照后去除盖玻片,将标本载玻片在2× SSC 中煮沸20min,75%、80%和100%梯度酒精中脱水10min,风干,然后将载玻片暴露在强烈的阳光下至少48h 后即可进行连续FISH 杂交,方法与上述杂交方法相同。
2 结果与分析
2.1 纤毛鹅观草原变种与竖立鹅观草变种核型数据15 份居群材料染色体数目都为28 条;核型类型均为2A;染色体主要由中部着丝粒染色体(11~12 对)和近中部着丝粒染色体(2~3 对)组成;含2 对随体染色体。纤毛鹅观草和竖立鹅观草居群间核型差异主要在核型公式、平均臂比、染色体长度比、臂比>2 的比例、核不对称系数等方面,如表2 所示。纤毛鹅观草原变种居群间平均臂比的变化范围为1.29~1.40;染色体长度比的变化范围为1.65~1.80;臂比>2 的比例为0.07 或0.14;核不对称系数为0.51~0.57。竖立鹅观草变种居群间的平均臂比变化范围为1.21~1.38,染色体长度比变化范围为1.59~1.75;臂比>2 的比例为0.07 或0.14;核不对称系数为0.54~0.62。其中,同一采集地的不同居群竖立鹅观草变种材料的核型数据也存在差异,如采集地是四川宜宾编号为88-89-252、88-89-263、88-89-261 的3 份材料,其核型公式分别为2n=4x=28=24m(2sat)+4sm(2sat)、2n=4x=28=22m(4sat)+6sm、2n=4x=28=24m(4sat)+4sm;平均臂比分别为1.21、1.36、1.38;染色体长度比分别为1.59、1.75、1.72;臂 比>2 的 比 例 分 别 为0.07、0.14、0.14;核不对称系数分别为0.54、0.57、0.57。

表2 供试材料染色体参数
2.2 纤毛鹅观草原变种和竖立鹅观草变种分子FISH 核型建立利用 StY_34、StY_90、StY_93、StY_107、St2-80、45S rDNA 对15 份不同居群材料的根尖有丝分裂中期细胞进行连续FISH 建立FISH 核型(图1)。首先,将St2-80 和45S rDNA 进行FISH 杂交分别用于区分St 染色体和核仁组织区(NORs)。在15 份居群材料中,有14 条染色体除了近着丝粒区域外,整个染色体都显示出St2-80 信号为St 染色体,其余14 条仅在染色体端部区域显示出荧光信号为Y 染色体,St 和Y 染色体上分别有1 对染色体产生45S rDNA 的信号。其次,使用StY_107 和StY_93 重复序列对相同有丝分裂中期细胞进行了FISH 检测。StY_107 与染色体端部、近端部以及靠近着丝粒区域杂交产生信号,而StY_93则主要在(近)着丝粒区域显示强烈信号,在远端区域显示分散信号。最后,使用StY_90 和StY_34 重复序列对同一细胞进行FISH 检测,结果显示染色体端部区域有强烈信号。根据相对长度和FISH 信号分布模式,用编号Ⅰ-Ⅶ任意指定了标记良好的染色体。
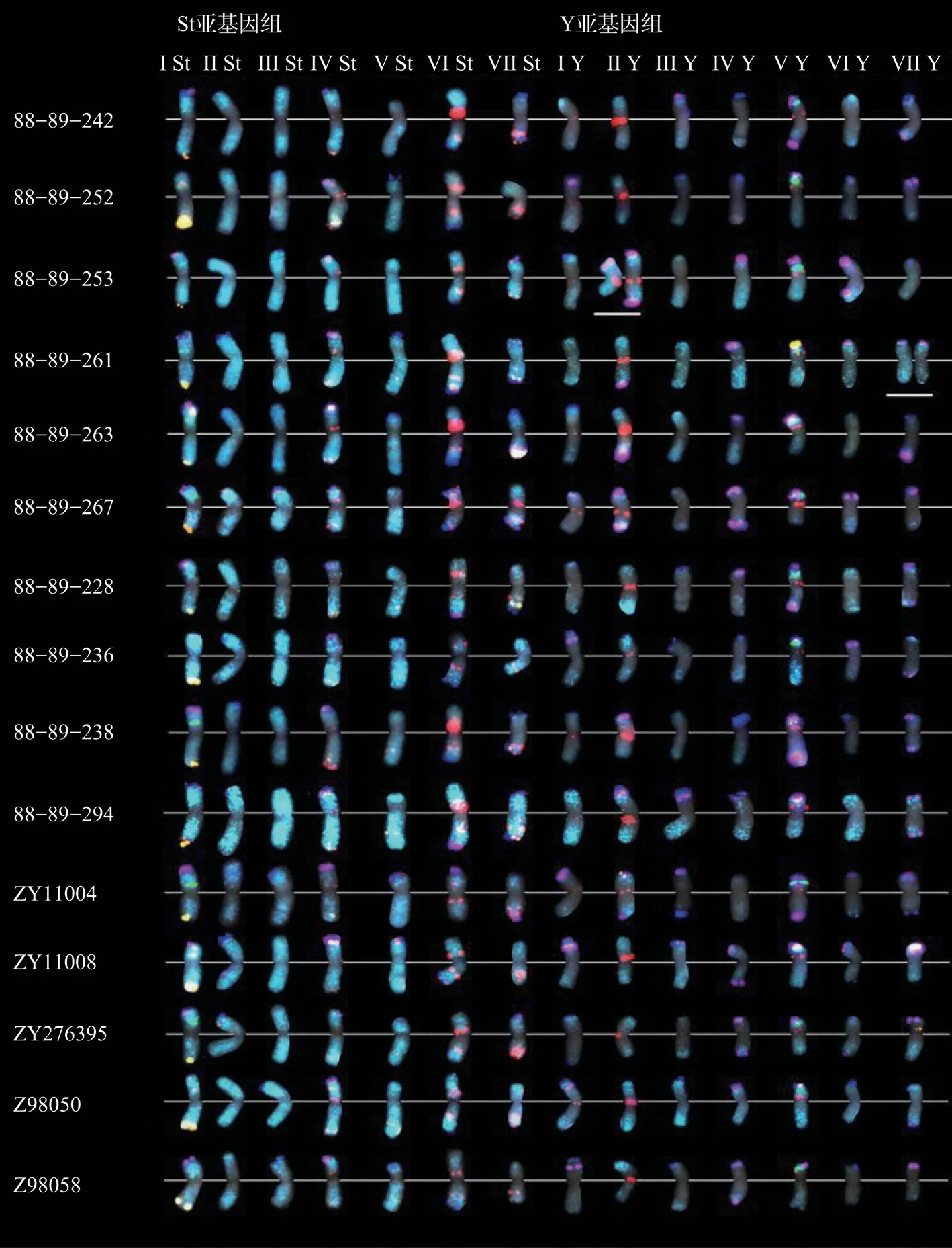
图1 纤毛鹅观草原变种和竖立鹅观草变种居群间FISH 核型图
在9 份纤毛鹅观草原变种居群材料中除St2-80 以外的5 个重复序列探针,共杂交产生了755对 信 号,45S rDNA、StY_93、StY_107、StY_34、StY_90 分别产生36、216、191、220、92 对信号;在6 份竖立鹅观草变种居群材料中除St2-80 以外的5 个重复序列探针,共杂交产生了458 对信号,45S rDNA、StY_93、StY_107、StY_34、StY_90分别产生24、137、116、126、55 对信号。6 个重复序列在纤毛鹅观草原变种和竖立鹅观草变种上的分布模式大致相同,但染色体上探针杂交信号的强弱存在差异,如探针StY_107 在纤毛鹅观草原变种的ⅠSt 上的杂交信号普遍强于竖立鹅观草变种,探针StY_93 在竖立鹅观草的Ⅵ St 染色体上出现的信号普遍强于纤毛鹅观草原变种。为了更直观准确地描述除St2-80 探针外的5 个探针的杂交信号在15 份居群材料的14 对染色体上的分布模式,建立了它们的FISH 核型模式图,对15 份居群材料的染色体上出现杂交信号位置和频率进行了统计,并将出现信号的频率和位置标注到相应染色体上构建了居群总的“泛FISH 核型”信号分布图(图2-A),“泛FISH 核型”显示探针更容易在纤毛鹅观草原变种染色体上产生特异的杂交位点,此外探针在竖立鹅观草变种的Y 染色体上也更容易出现特异的杂交位点。将染色体上出现频率≥ 50.0%的杂交信号构建居群“核心FISH 核型”(图2-B)。“核心FISH核型”显示了34 个FISH 信号:2 个45S rDNA 信号、12 个StY_34 信 号、7 个 StY_90 信 号、9 个StY_93 信号和4 个StY_107 信号。不同染色体的信号数量从0 到5 个不等,其中3 St 的信号最少(0个),4 St 和5 Y 信号最多(5 个)。“核心FISH 核型”只显示保守信号,因此更具有代表性并帮助确定纤毛鹅观草和竖立鹅观草染色体的身份。

图2 纤毛鹅观草原变种和竖立鹅观草变种的“泛 FISH 核型 ”(A)和 “核心 FISH 核型”(B)
2.3 纤毛鹅观草原变种和竖立鹅观草变种分子FISH 核型比较分析为了描述探针FISH 信号在不同居群间的多态性,对15 个居群的FISH 核型进行了比较分析(图3)。在试验中发现15 个居群的St 和Y 染色体上共发现 61 个染色体FISH 信号差异,其中Y 染色体的差异(33 个)大于St 染色体的差异(28 个)。在 St 亚基因组中,Ⅶ St 染色体对的变异最大(9 个),Ⅱ-Ⅵ St 均含有3 个染色体变异类型,而在Y 亚基因组中V Y 染色体对的变异最少(3 个),Ⅰ Y 和Ⅶ Y 的变异最多(6 个)。

图3 纤毛鹅观草原变种和竖立鹅观草变种居群间FISH 核型比较图
3 讨论
纤毛鹅观草具有丰富遗传多样性。前人对纤毛鹅观草原变种和竖立鹅观草变种的核型进行了大量的研究。如蔡联炳等[20]以及孙义凯等[21]对纤毛鹅观草的核型进行了报道,核型公式分别为2n=4x=28=20m + 8sm(4SAT),2n=4x=28=22m(2SAT)+6sm(2SAT),但是他们没有对纤毛鹅观草变种材料核型进行区分;卢宝荣等[9]和魏秀华等[10]报道了纤毛鹅观草的变种材料纤毛鹅观草原变种和竖立鹅观草变种的核型公式,但是核型公式间存在差异。本试验中纤毛鹅观草原变种的核型公式为2n=4x=28=22m(4sat)+6sm、2n=4x=28=22m(2sat)+6sm(2sat)或2n=4x=28=24m(4sat)+4sm;竖立鹅观草变种的染色体核型公式为2n=4x=28=24m(4sat)+4sm、2n=4x=28=24m(6sat)+4sm、2n=4x=28=22m(4sat)+4sm、2n=4x=28=24m(2sat)+4sm(2sat)或2n=4x=28=22m(4sat)+6sm,与前人已报道的核型公式存在差异,这可能与试验材料的采集地、试验方法有关。纤毛鹅观草原变种、竖立鹅观草变种以及两个变种的居群材料间在平均臂比、染色体长度比、核不对称系数、臂比>2 的比例存在差异,且同一变种来自同一地方的不同居群间也存在一定的差异,表明纤毛鹅观草种下水平存在丰富的遗传多样性。
本研究利用FISH 技术对纤毛鹅观草物种的变种材料染色体进行细胞遗传学核型分析,为纤毛鹅观草物种种下水平的遗传多样性提供参考依据。结果表明这6 个重复序列探针在这2 个变种的15 份居群材料中的分布模式基本相似但也存在差异,且竖立鹅观草变种的同源染色体变异的频率(33.3%)大于纤毛鹅观草原变种(0)。近年来,关于St 和Y 亚基因组的染色体结构变异展开了大量的研究,如Liu 等[22]利用FISH 和GISH 技术表征了不同鹅观草属物种(StY)的St 基因组和Y 基因组核型以探究不同StY 物种间的基因组分化;Dou等[23-24]在披碱草属物种Elymus nutans(StYH)中发现St 和Y 染色体存在丰富的染色体结构变异,且St 亚基因组的变异少于Y 亚基因组;Cheng等[13]对53 个野生纤毛鹅观草(R.ciliaris)品系建立FISH 核型发现了纤毛鹅观草存在丰富的染色体结构变异,且Y 基因组的染色体变异较多。本研究通过FISH 核型比较分析结果表明纤毛鹅观草种下水平的居群材料间具有丰富的变异,且Y 亚基因组的变异多于St 亚基因组,这同样也表明了纤毛鹅观草和竖立鹅观草具有丰富的遗传多样性。
植物染色体FISH 核型图的建立在植物分类及进化研究和植物育种特别是植物远缘杂交育种中具有重要的作用。本研究通过连续FISH,对出现频率≥50.0%的杂交信号构建纤毛鹅观草与竖立鹅观草的“核心FISH 核型”,为纤毛鹅观草原变种与竖立鹅观草变种的染色体结构变异提供了细胞学基础,也为远缘杂交育种产生的小麦-纤毛鹅观草异附加系中纤毛鹅观草的染色体鉴定提供了依据。
4 结论
纤毛鹅观草作为小麦的三级基因资源库含有优异的抗病和抗逆基因,但关于纤毛鹅观草变种之间和居群之间的遗传多样性和染色体水平的变异的研究较少,使纤毛鹅观草的有效利用受到限制。本研究通过染色体核型和分子FISH 核型揭示了纤毛鹅观草种下水平具有丰富遗传多样性和染色体结构变异,并建立了种下变种纤毛鹅观草变种和竖立鹅观草变种的“核心FISH 核型”,为了解纤毛鹅观草以及鹅观草属野生材料染色体特征提供依据,同时也为纤毛鹅观草的进一步利用提供了染色体水平依据。
