凋落物输入对土壤呼吸温度敏感性的影响*
2023-12-24张彦军李天姿
于 媛,张彦军,陈 曦,李天姿
(宝鸡文理学院 地理与环境学院,陕西 宝鸡 721013)
土壤呼吸温度敏感性一般用Q10表示,其含义是温度每升高10 ℃后土壤呼吸所增加的倍数,反映的是土壤呼吸对温度的敏感程度[1]。大量的研究结果显示Q10呈现出巨大的时空变异性,且Q10的时空变异性与地形[2]、气候[3-4]、植被类型[5]等因素密切相关。地表凋落物(简称为凋落物,本文如果不特指凋落物均表示地表凋落物)作为联系地上和地下部分的枢纽,对陆地生态系统碳循环产生显著的影响。大量的研究结果表明,凋落物管理措施对土壤呼吸影响显著,添加凋落物会导致土壤呼吸的显著增加,而去除凋落物则会导致土壤呼吸速率减少[6-7],而凋落物管理措施对土壤呼吸温度敏感性的影响相对研究较少,且目前没有定论[7-8]。例如,刘益君等[9]发现凋落物添加后导致Q10呈现增加的趋势,但是覃志伟等[10]研究认为凋落物添加后导致Q10降低;同时,YAN et al[11]研究认为凋落物去除后Q10呈现出增加的趋势,而杨淞等[12]研究发现凋落物去除会导致Q10降低。Q10通常被用来衡量全球变暖和土壤碳循环之间的反馈强度,在各种碳循环估算模型中被广泛应用,因此Q10估算的准确与否也会对模型预测结果产生显著的影响[13]。基于此,本文利用国内外已发表的凋落物管理措施与Q10相关的47篇研究论文,全面阐述凋落物管理措施对Q10的影响,并分析可能的影响因素,以期为全球气候变化以及固碳减排、碳循环的研究提供重要的依据和支撑[14]。
1 材料与方法
1.1 文献检索
以“土壤呼吸温度敏感性、凋落物呼吸、凋落物与土壤呼吸,soil respiration temperature sensitivity, litter respiration, litter and soil respiration”等作为关键词,在中国知网、维普、万方、Web of Science等中英文数据库中搜寻2008年1月-2022年6月发表的“凋落物土壤呼吸温度敏感性”相关文献展开分析,为本研究提供所需数据。
为使研究结果更加精确,更好地达到研究目的,本文所选的文献必须满足以下条件:(1)研究试验必须是田间定点试验;(2)当试验地点、试验时间、凋落物处理一样时,应选择研究时限最长、发表时间最新的文献。通过上述条件,最终筛选出符合条件的文献47篇。
1.2 数据处理
在获取本文所需要的数据时,一部分数据可以从论文中直接获得,另一部分数据需要根据论文中提供的数据用式(1)进一步计算:
Q10=e10b
(1)
式中,b为拟合参数。
在SigmaPlot10.0软件中,输入不同凋落物管理措施下的去凋、对照、倍增及凋落物Q10值,应用数据透视表根据不同凋落物管理措施对数据进行组合分类。
为证明本研究结果的合理性,运用公式(2)对获取的Q10值进行对数转换,在OriginPro9软件中,输入处理后的数据进行K-S检验并绘制直方图,得出P>0.05,即本研究获得的有关呼吸温度敏感性的数据符合正态分布,从而佐证了本研究结果的合理性(图1)。
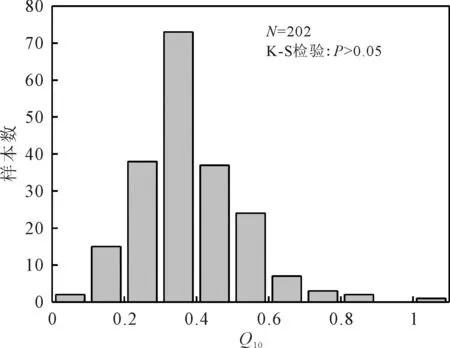
图1 Q10分布直方图Fig. 1 Q10 distribution histogram
Y=log10X
(2)
式中,X为获取的Q10值,Y为转换后的对应值。
1.3 数据分组
由于植被因素(植被类型、凋落物输入量和林龄)、气候(温度和降水)、地形因素(海拔)、土壤理化性质(土壤容重、土壤pH、土壤碳氮比C∶N、土壤有机碳SOC)等因素会对Q10造成影响,本研究对获取的数据首先根据不同调控因子的属性将其划分为植被、地形、气候、理化性质4组;其次将植被类型划分为草、针叶林、阔叶林、混交林,而凋落物输入量、林龄等其他调控因子的数据分组则是根据每个因素的具体数据量进行划分,进而分析在不同的凋落物、不同的调控因子影响下Q10的变化规律及趋势,具体分类如表1所示。

表1 数据分组依据Tab. 1 Data grouping basis
2 结果分析
2.1 植被因素调控凋落物管理措施对Q10的影响
本研究结果表明,凋落物管理措施对Q10影响显著,基本呈现出凋落物Q10>对照Q10>倍增Q10>去凋Q10的趋势,但这种影响会因植被因素而发生显著的改变(图2)。

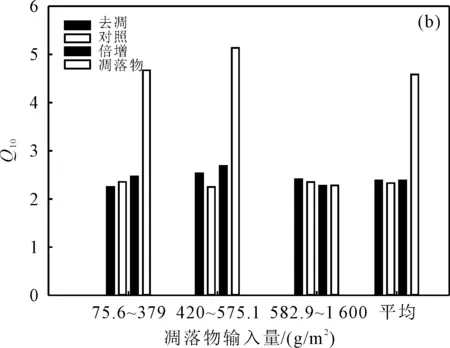
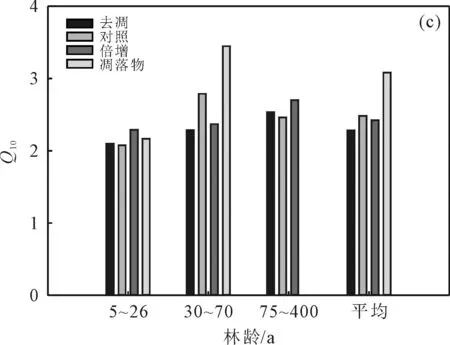
图2 植被因素调控凋落物管理措施对Q10的影响Fig. 2 Vegetation factors regulate the effects of litter management measures on Q10
凋落物Q10在不同植被条件下呈现出阔叶林>针叶林的趋势(图2(a));去凋Q10在草、混交林、阔叶林和针叶林条件下依次为1.75,2.41,2.54和2.06;对照Q10最大值出现在针叶林条件,高达2.70,最小值出现在草条件,仅为1.94;倍增Q10在混交林和阔叶林条件下分别是针叶林条件下的1.04和1.18倍。
凋落物Q10在不同凋落物输入条件下呈现出中凋落物输入>低凋落物输入>高凋落物输入的趋势(图2(b));去凋Q10在低凋落物输入、中凋落物输入和高凋落物输入条件下依次为2.25,2.53和2.41;对照Q10最大值出现在低凋落物输入条件下,高达2.35,最小值出现在中凋落物输入条件下,仅为2.25;倍增Q10在低凋落物输入和中凋落物输入条件下分别是高凋落物输入条件下的1.08和1.18倍。
凋落物Q10在不同林龄条件下呈现出中林龄>低林龄的趋势(图2(c));去凋Q10在低林龄、中林龄和高林龄条件下依次为2.10,2.29和2.51;对照Q10最大值出现在中林龄条件下,高达2.79,最小值出现在低林龄条件下,仅为2.08;倍增Q10在中林龄和高林龄条件下分别是低林龄条件下的1.04和1.18倍。
2.2 气候因素调控凋落物管理措施对Q10的影响
不同气候条件下的Q10呈现出凋落物Q10>对照Q10>倍增Q10>去凋Q10的趋势,但会受到气温和降水等气候因素的影响(图3)。
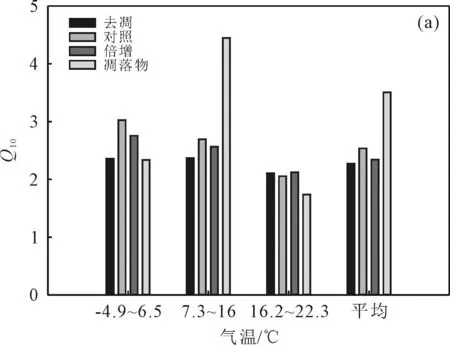

图3 气候因素调控凋落物管理措施对Q10的影响Fig. 3 Climate factors regulate the effects of litter management measures on Q10
凋落物Q10在不同温度条件下呈现出中温>低温>高温的趋势(图3(a));去凋Q10在低温、中温和高温条件下依次为2.36,2.37和2.11;对照Q10最大值出现在低温条件下,高达3.02,最小值出现在高温条件下,仅为2.05;倍增Q10在低温和中温条件下分别是高温条件下的1.30和1.21倍。
凋落物Q10在不同降水条件下呈现出中降雨量>高降雨量>低降雨量的趋势(图3(b));去凋Q10在低、中和高降雨条件下依次为2.02,2.61和2.24;对照Q10最大值出现在中降雨条件,高达2.87,最小值出现在高降雨条件,仅为2.22;倍增Q10在低降雨和中降雨条件下分别是高降雨条件下的1.14和1.17倍。
2.3 地形因素调控凋落物管理措施对Q10的影响
地形因素会调控凋落物管理措施对Q10的影响,但整体上仍然呈现出凋落物Q10>对照Q10>倍增Q10>去凋Q10的趋势(图4)。凋落物Q10在不同海拔条件下呈现出中海拔>低海拔>高海拔的趋势;去凋Q10在低、中和高海拔条件下依次为2.37,2.23和2.44;对照Q10最大值出现在高海拔条件,高达2.82,最小值出现在低海拔条件,仅为2.28;倍增Q10在中海拔和高海拔条件下分别是低海拔条件下的1.08和1.09倍。
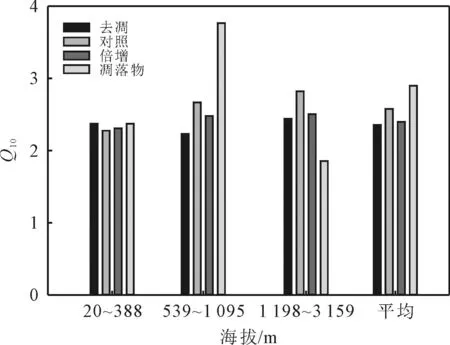
图4 地形因素调控凋落物管理措施对Q10的影响Fig. 4 Topographic factors regulate the effects of litter management measures on Q10
2.4 土壤理化性质调控凋落物管理措施对Q10的影响
不同凋落物管理措施下的Q10会因土壤理化性质发生显著改变,但整体上依然呈现出凋落物Q10>对照Q10>倍增Q10>去凋Q10的趋势(图5)。




图5 土壤理化性质调控凋落物管理措施对Q10的影响Fig. 5 Effects of soil physicochemical properties regulating litter management measures on Q10
凋落物Q10在不同容重条件下呈现出中容重>低容重>高容重的趋势(图5(a));去凋Q10在低、中和高容重条件下依次为2.21,2.08和2.07;对照Q10最大值出现在低容重条件,高达2.57,最小值出现在高容重条件,仅为2.10;倍增Q10在低容重和中容重条件下分别是高容重条件下的1.06和1.85倍。
凋落物Q10在不同pH条件下呈现出高pH>中pH>低pH的趋势(图5(b));去凋Q10在低、中和高pH条件下依次为2.16,2.56和2.06;对照Q10最大值出现在高pH条件,高达2.83,最小值出现在低pH条件,仅为2.19;倍增Q10在中pH和高pH条件下分别是低pH条件下的1.01和1.46倍。
凋落物Q10在不同C∶N条件下呈现出低C∶N>中C∶N>高C∶N的趋势(图5(c));去凋Q10在低、中和高C∶N条件下依次为2.34,2.46和2.19;对照Q10最大值出现在中C∶N条件,高达2.63,最小值出现在低C∶N条件,仅为2.41;倍增Q10在低C∶N和中C∶N条件下分别是高C∶N条件下的1.25和1.37倍。
凋落物Q10在不同SOC条件下呈现出低SOC>高SOC>中SOC的趋势(图5(d));去凋Q10在低、中和高SOC条件下依次为2.18,2.32和2.38;对照Q10最大值出现在中SOC条件,高达2.69,最小值出现在低SOC条件,仅为2.16;倍增Q10在中和高SOC条件下分别是低SOC条件下的1.05和1.22倍。
3 讨论
3.1 底物输入对Q10的影响
呼吸底物的分解是土壤呼吸过程中产生CO2的主要来源,底物(凋落物)质量和数量会对Q10造成影响。本文结果显示,凋落物Q10显著大于土壤呼吸Q10(去凋Q10、对照Q10、倍增Q10),主要是因为底物质量不同,底物的分解速率会随着底物质量提高而加快;例如MELILLO et al[15]在北美中纬度硬木林10 a的土壤升温实验中表明,土壤中质量较高、易分解的碳库Q10比较高,而质量较低的、难分解的碳库对温度相对较不敏感。
底物数量也会影响Q10的变化。本研究中呈现出对照Q10>倍增Q10>去凋Q10的趋势,表明添加或去除凋落物输入都会不同程度降低土壤呼吸Q10,这与RICHARD et al[16]的研究结果相同,可能是因为去除凋落物后会使土壤对外界环境变化的抵抗能力减弱,进而导致Q10降低;同时,去除凋落物后会引起土壤水分降低,导致底物的可利用性降低,也会降低Q10[17];添加凋落物后Q10减小是由于添加凋落物的屏蔽作用阻隔了土壤与外界空气的热交换,进而对土壤温度、湿度产生影响[18],导致土壤呼吸对温度变化的敏感性降低;本研究中去凋Q10降低幅度显著大于倍增Q10降低幅度,与葛晓改等[19]的研究结果一致,这是因为去除凋落物相较于添加凋落物对外界环境的影响更为明显。
3.2 植被因素对Q10的影响
陈玮[5]在太行山南麓展开研究得出林地Q10>灌丛Q10>草地Q10>裸地Q10,说明植被是影响Q10的一个重要因素。本研究中,Q10呈现阔叶林>混交林>针叶林>草的趋势。冯华敏等[20]分析了重庆市缙云山自然保护区内的毛竹林、针叶林和针阔混交林的土壤呼吸速率,发现毛竹林Q10>针阔混交林Q10>针叶林Q10,这可能与不同植被的郁闭度不同有关。不同植被的生态环境因子(湿度、温度、土壤酸碱度及有机质的含量等)是不同的,因此土壤呼吸的强度也会存在差异;并且不同植被的凋落物数量及质量也是不同的,这些都会导致真菌的异氧呼吸或者土壤表层微生物的分解时间发生变化[21],进而影响Q10变化。
本文研究结果显示,Q10随着林龄的增加呈上升趋势,与文献[22-23]的研究结果相类似。例如文献[23]研究了黄土高原的不同林龄刺槐林,得出了林龄增大时Q10也随之增大的结果,这是由于林龄会影响林下植被生物量、凋落物量等,也会对气温、土壤温度的影响造成差异[24];林龄发生改变时,森林生态系统中的群落结构、物种组成、物种丰富度、碳的分配及生物量的积累等都会随之改变,同时森林生态系统中的土壤碳库和碳吸存潜力也会产生变化[25-26],从而影响土壤呼吸和Q10。
3.3 气候因子对Q10的影响
研究结果显示,随着温度的增加,Q10呈现出低温>中温>高温的趋势,而降雨量增加后Q10呈现出先增后减的趋势。
温度会影响酶的活性,当温度过高甚至超过一定界限时,酶的活性则会显著降低,但是土壤呼吸离不开酶的作用,因此温度也会影响到土壤呼吸及其温度敏感性[1]。张克胜等[27]通过研究中国不同气候带(热带、亚热带、暖温带、中温带以及高原气候区)的土壤呼吸差异,得出了Q10沿气候梯度从南到北逐渐增加的结论。胡汗等[4]在太白山北坡的研究结果表明,在低温时土壤呼吸对温度的变化更敏感。这些均与本文研究结果一致。
降雨对土壤温度和土壤湿度均会产生影响,并影响土壤呼吸的动态[28];土壤中生物活动和植物根系生长需要的水量、土壤水分含量与土壤温度等均会受到降水的影响,从而影响土壤呼吸[21]。文献[3,29]的研究结果与本研究结果相反,如文献[29]在中国科学院黄河三角洲滨海湿地生态试验站内的研究结果显示,随着降水量的增加,Q10也随之增大,这可能与降雨前土壤中水分含量密切相关,如果降雨前土壤含水量已达到了某个临界值,则整个降水过程都会抑制土壤呼吸[30]。
3.4 地形对Q10的影响
地形会导致水热条件变化,使得凋落物的分解过程随海拔而变化[31],海拔的改变会导致植被类型、土壤生物、林分生产力、立地环境条件等因子发生变化[32],同时对土壤呼吸的大小及Q10产生显著影响。本文研究结果显示,随着海拔升高,Q10也随之增大,这与文献[2,33]的研究结果相类似。例如文献[33]在武夷山自然保护区内部的研究表明,Q10会随着海拔的升高而增大。随着海拔的升高,温度也随之降低,而本文本研究结果表明Q10随温度增加而降低,这也证明了高海拔地区土壤呼吸的温度敏感性更强。
3.5 土壤理化性质对Q10的影响
Q10还会受到理化性质调控而发生变化。其中,容重与Q10呈负相关;pH和SOC与Q10呈正相关;Q10会随着C∶N的增大先增大后减小。
土壤容重反应的是土壤呼吸排放通道的顺畅程度[34],它通过影响土壤环境中的水分、有机质、温度、气体等,进而引起植物的生长发育发生变化,是土壤紧实度和孔隙状况的指标之一[35]。张金波等[36]研究了三江平原在不同土地利用条件下的Q10变化,发现当容重为0.46 g/cm3时,Q10为4.29;当容重增加至1.05 g/cm3时,Q10降低为1.45。这是由于土壤通气性、持水性和渗透性会随土壤容重的增加逐渐降低,土壤表面附着的凋落物数量也随之减少,致使土壤根系对营养元素的吸收受到限制,进而影响土壤根系呼吸。而通气孔隙度也随着容重的增大逐渐降低,使土壤内部逐渐趋于厌氧环境,土壤微生物活性受到限制,从而影响微生物参与的一系列土壤呼吸过程[37]。
土壤pH可调控化学反应和微生物酶的多样性[38],对微生物的生长以及土壤呼吸均有明显的影响,土壤pH过低会抑制土壤微生物和酶的活性,不同微生物活动有其适宜生存的pH,pH越适宜则微生物活性更强[39]。土壤pH变化的一个重要因素是酸雨,例如冯继广等[40]的研究表明低和高强度的酸雨都会降低Q10;梁国华等[41]通过在鼎湖山国家级自然保护区内的酸雨模拟实验得出了Q10随pH的降低而呈现减小的趋势,这些都说明酸雨在一定程度上会降低Q10。文献[5,42]的研究结果与本研究结果不符,可能是由于植被类型、凋落物以及其他环境因子等的不同。
土壤C∶N是土壤质量的敏感指标,会通过影响土壤中的微生物活性,进一步影响土壤中的有机碳和氮循环[43],不同的微生物分解有其最佳的土壤C∶N值[34],土壤中的C和N含量的变化都会对C∶N造成影响。刘宝等[44]在研究中亚热带常绿阔叶林改造后土壤呼吸变化时,得出Q10与C∶N极显著正相关(P<0.1),这与本文的研究结果不相符,可能是因为土壤C∶N的高低对植物生长发育较为重要,C∶N过高时则会出现微生物在分解有机质的过程中存在氮受限的情况[45],而本文所选取的C∶N超过了这个范围,因此会呈现出先增后减的趋势。
微生物的分解活动排放CO2是以土壤有机碳SOC为物质基础的,因而SOC对土壤呼吸十分重要[46]。土壤全碳及有机碳对微生物的生长发育意义重大,其含量的高低会直接引起土壤微生物活性的变化,从而影响土壤呼吸[47]。文献[46,48]的结论与本文一致,文献[46]研究了新疆玛纳斯河流域不同恢复模式下SOC及土壤呼吸速率的变化特征,发现SOC与Q10呈正相关关系,但其相关性不显著。这可能是由于随着SOC的增加,土壤养分也随之增加,供给土壤微生物呼吸的基质随之增多,而温度对有机质降解影响较大,因而对于温度更为敏感[46-49]。
4 结论
(1)凋落物输入后凋落物Q10显著大于土壤呼吸的Q10,且呈现出对照Q10>倍增Q10>去凋Q10的趋势,但同时受到植被类型、气候、地形及土壤理化性质等因素的调控。
(2)Q10的增加程度在不同植被类型下呈现出阔叶林>混交林>针叶林>草的趋势,且Q10随着林龄的增加而增大;
(3)Q10随着温度的增加呈现出逐渐降低的趋势,随着降雨量增加Q10呈现出先增后减的趋势;
(4)在不同海拔梯度条件下,高海拔Q10最大,而低海拔Q10最低;
(5)Q10随着容重的增加而逐渐减小,pH以及SOC与Q10呈正相关,Q10随着C∶N的增大呈现先增大后减小的趋势。
