基于叶片解剖性状探究29 种植物对热带珊瑚岛的适应策略
2023-12-21周雨珩刘慧张世柯刘芳延刘楠
周雨珩, 刘慧, 张世柯, 刘芳延, 刘楠*
(1. 中国科学院华南植物园,中国科学院海岛与海岸带生态修复工程实验室,广州 510650;2. 中国科学院大学,北京 100049;3. 华南国家植物园,广州 510650)
我国南海海域分布着200 多个热带珊瑚岛、珊瑚礁和沙洲,这些珊瑚岛具有重要的战略地位和开发利用潜力[1],受到诸多海洋生态学家和自然资源保护者的关注。以南海诸岛为代表的中国热带珊瑚岛缺少常规土壤结构,基质主要由珊瑚砂构成,保水保肥能力差且养分贫瘠,其环境具有高盐、高温、强碱、强光以及季节性干旱明显等极端环境特点,植被极易退化[2]。植被作为海岛的三大基本组成要素之一,具有供给、调节、文化和支持服务等生态系统服务功能,是人类和其它动物赖以生存的基础,也是海岛宜居和可持续发展的基础。因此,对热带珊瑚岛植被进行人为恢复确有必要且具有重要意义[3]。由于热带珊瑚岛的极端环境,选取适应性强的树种可有效提高植被恢复效率[4]。
叶片功能性状是植物在与外界环境长期相互作用下形成的形态结构和生理生态等特征,是植物响应外界胁迫的重要组成部分,具有较强的可塑性,常用于反映植物的生长状况和环境适应性[5]。植物在长期演化过程中,不断适应周边的复杂生长环境,产生了一系列适合自身生长发育的生理生态行为。以干旱逆境为例,大量研究表明干旱条件下植物的叶片厚度、栅栏组织与海绵组织厚度以及二者之比均显著增加,同时气孔导度、气孔密度降低[6–8]。其中,叶片厚度是反映植物抗旱性的重要指标,较厚的叶片可以降低水分散失,并提高保水能力[9]。高度发达的栅栏组织在避免植物叶肉细胞过度失水的同时又可有效利用衍射光进行光合作用[10],排列紧密的海绵组织是植物适应干旱与强光环境的结构基础[11–12]。因此,旱生植物的比叶面积(SLA)较低, 其叶片较厚,比叶面积偏低,在减少蒸腾面积的同时也降低了光资源竞争能力,叶肉倾向于向提高光合效能的方向发展。气孔是植物与外界进行气体交换的主要通道,其大小和密度对植物水分状况有着重要影响,较低的气孔密度和气孔长度有利于减少蒸发丧失的水分从而增加植物的抗旱能力[13]。这些在逆境胁迫下植物叶片形态结构特征的变化反映了植物的抗逆性。因此,叶片功能性状是研究植物响应逆境机制的重要切入点。
目前有大量关于植物功能性状与环境因子关系的研究,但较少聚焦于热带珊瑚岛的极端环境。关于植物在该环境条件下的适应性研究,多为对个别生物物种进行比较[4,8–9,14],难以揭示植物对热带珊瑚岛极端环境普适的适应机制。基于叶片形态解剖结构分析植物的适应性研究较多[15–16],但针对退化生态系统植被恢复及物种选择方面的植物适应性分析较少,还有待进一步深入。本研究以生长于海南文昌苗圃和移栽到热带珊瑚岛2 种不同生境的29 种热带海岛及海岸带常见植物为研究对象,希望回答如下的科学问题:相对于环境条件良好的苗圃,移栽到热带珊瑚岛上的植物,如何通过调节叶片解剖性状适应干旱、强光等极端环境?相关研究结果将为热带珊瑚岛植被新建物种的选择与利用提供科学依据。
1 材料和方法
1.1 研究地概况
海南省文昌市(110°45′ E、19°31′ N),地势低平,海拔约10 m,属热带海洋性季风气候;年均气温约24 ℃, 年降雨量约1 800 mm,土壤类型为滨海沉积物沙壤土。热带珊瑚岛位于海南省三沙市,海拔约5 m,属热带季风海洋性气候。太阳辐射约6 000 MJ/m2,年均温约28 ℃,年降雨量约2 800 mm,降雨量充沛但分配不均,每年6 月—11 月为湿季,12 月—翌年5 月为干季,雨季旱季明显;基质为珊瑚砂,保水保肥性差,盐碱度高,养分贫瘠[2]。
1.2 试验材料
本研究以前期西沙群岛植被调查为基础[1],参照《热带珊瑚岛礁植被恢复工具种图谱》[17],选取了29 种全球热带海洋岛屿共有优势物种作为研究对象(表1),包括椰子(Cocos nucifera)、木麻黄(Casuarina equisetfolia)、草海桐(Scaevola sericea)、扶桑(Hibiscus rosa-sinensis)等19 种木本物种,以及滨豇豆(Vigna mrina)、海刀豆(Canavalia biflora)等10 种草藤本植物。

表1 热带珊瑚岛29 种植物基本信息Table 1 Information of 29 common species in tropical coral islands, China
1.3 研究方法
本试验中的29 种苗木原先种植于海南省文昌市苗圃基地,苗圃位于海南省文昌市郊区,水肥管理采用常规的养护方式,提供充足的水肥,保证植株生长情况良好。然后选取大小及长势一致,生长情况良好的部分植株于2015年5月移植至热带珊瑚岛,在3 d 内完成种植,施入客土,浇水定根。
在苗木移栽后的第2年进行两地植物叶片形态解剖性状的测定,选择胁迫严重的3 月完成叶片采集。采集时排除幼叶和残缺叶,选取枝条顶端长势良好、完全展开的叶片,每种植物选择5 株苗木, 每株苗木选择3~5 片叶片。采集后将样品分别装袋密封低温保存,一并带回实验室,测定其形态学特征、解剖结构和气孔特性等指标(表2),每个指标测量10 个重复值。
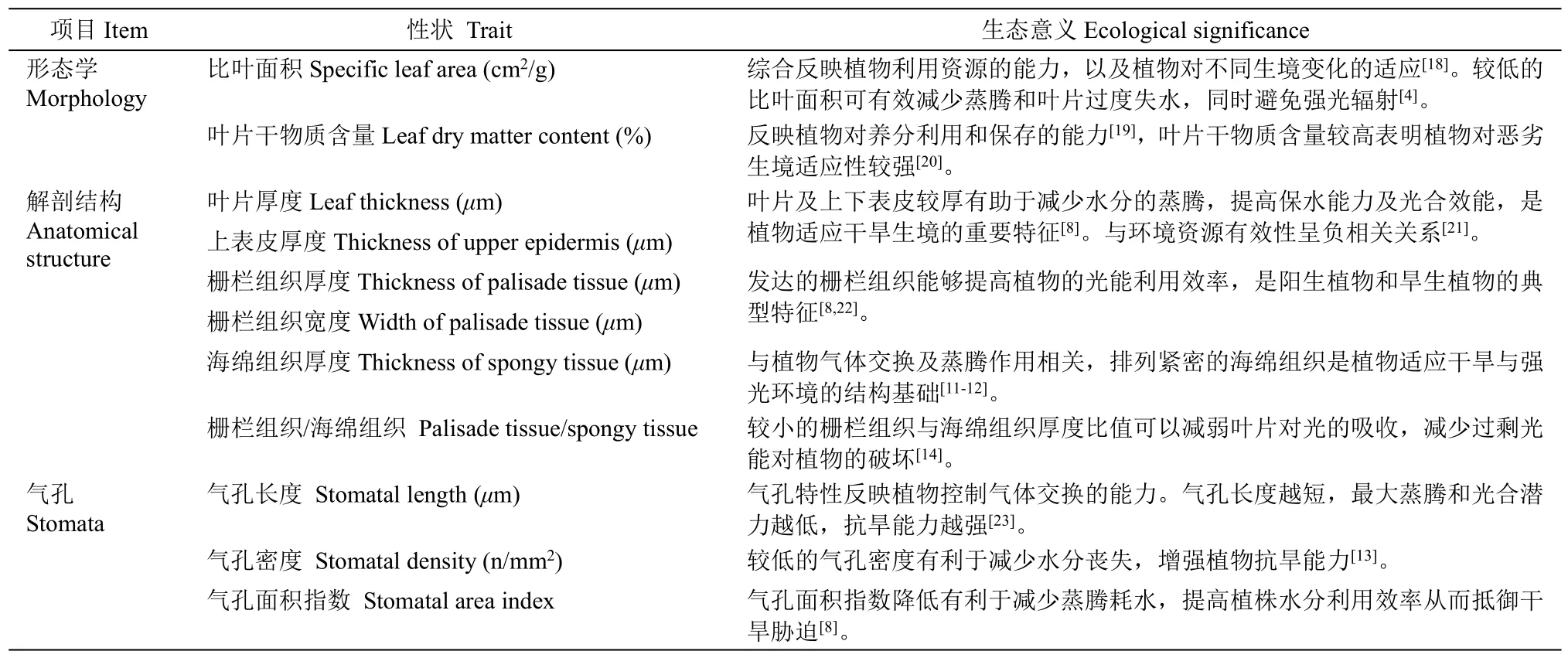
表2 性状指标及其生态学意义Table 2 Character index and its ecological significance
1.3.1 形态学特征测定
采用电子天平称量植物叶片鲜重(FW),然后置于65 ℃的烘箱中烘干,称量叶片的干重(DW),计算叶片干物质含量LDMC=DW/FW 和比叶面积SLA=LA/DW[24]。
1.3.2 解剖结构特征测定
叶片解剖结构的观察采用徒手切片法,选取健康植株成熟完整的叶片,参照常英俏等[25]的方法制作横切面切片。将切片置于显微镜(Leica DM 4000B)下,测量并记录叶片的厚度、上表皮厚度、栅栏组织厚度、栅栏组织宽度及海绵组织厚度,并计算栅栏海绵比PST=栅栏组织厚度/海绵组织厚度。
1.3.3 叶片气孔特性测定
采用指甲油印膜法[26]观察叶片气孔特性,将无色指甲油涂于植物叶片下表面,待指甲油干后取下表皮制成临时切片。将切片置于光学显微镜(Leica DM 4000B)下,选取合适视野进行观察记录。统计300 μm×300 μm 视野中的气孔数量,测量气孔长度,计算气孔密度。根据气孔长度(SL)及气孔密度(SD)计算气孔面积指数SPI (%)=SD×SL2×10–4[21]。每片叶随机选取10~15 个视野进行观察。
1.4 数据分析
采用SPSS 18.0 软件对生长在不同生境的同种苗木的性状进行成对t 检验,比较木本植物和草藤本植物在不同生境下的叶片解剖性状差异。采用Pearson相关系数检验木本和草藤本植物在不同生境下SLA和LDMC间的相关关系,利用Origin 2019b进行作图。
2 结果和分析
2.1 叶片形态学特征
与海南文昌植株相比,热带珊瑚岛地区木本和草藤本植物的LDMC 均显著上升(P<0.05),木本植物的SLA 显著下降(P<0.01),但草藤本植物未有显著变化(图1)。
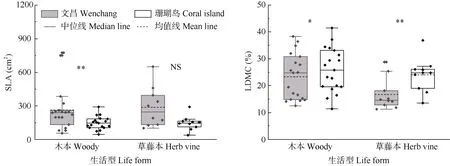
图1 两地植物的比叶面积(SLA)和叶片干物质含量(LDMC)比较。***: P<0.001; **: P<0.01; *: P<0.05; NS: P>0.05。下同Fig. 1 Comparison of specific leaf area (SLA) and leaf dry matter content (LDMC) of plants in two places. ***: P<0.001; **: P<0.01; *: P<0.05; NS: P>0.05.The same below
2.2 叶片解剖结构特征
从图2 可见,热带珊瑚岛木本植物的叶片厚度极显著高于海南文昌的(P<0.001),栅栏组织厚度和海绵组织厚度也显著上升(P<0.05),但栅栏海绵组织比、栅栏组织宽度及上表皮厚度无显著变化。草藤本植物的叶片解剖结构性状在两地间均无显著差异(P>0.05)。

图2 两地植物的叶片解剖结构性状的比较。LT: 叶片厚度; UET: 上表皮厚度; PT: 栅栏组织厚度; PW: 栅栏组织宽度; ST: 海绵组织厚度。Fig. 2 Comparison of leaf structure traits of plants in two places. LT: Leaf thickness; UET: Thickness of upper epidermis; PT: Thickness of palisade tissue;PW: Width of palisade tissue; ST: Thickness of spongy tissue.
2.3 叶片气孔特性
两地29 种植物木本和草藤本植物的叶片气孔长度、气孔密度和气孔面积指数的差异均未达显著水平(P>0.05)(图3)。
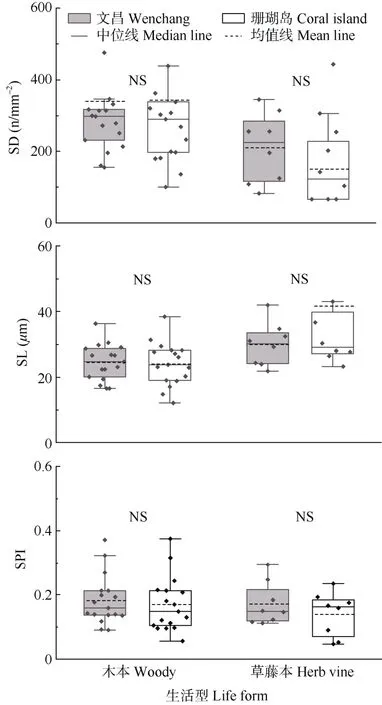
图3 两地植物叶片气孔性状的比较。SL: 气孔长度; SD: 气孔密度; SPI:气孔面积指数。Fig. 3 Comparison of leaf stomatal traits of plants in two places. SL: Stomatal length; SD: Stomatal density; SPI: Stomatal area index.
2.4 植物SLA 与LDMC 的关系
对两地木本植物与草藤本植物的SLA 和LDMC进行相关分析,结果表明,海南文昌地区木本植物与热带珊瑚岛地区木本植物的SLA 和LDMC 间的相关系数分别为-0.535 和-0.572,呈显著负相关(P<0.05),而两地草藤本植物的SLA 与LDMC 间的相关系数分别为-0.298 和-0.182,负相关关系均不显著(P>0.05)。
3 结论和讨论
3.1 热带珊瑚岛生境对植物功能性状的影响及其生存策略
热带珊瑚岛的极端生境(高温、强光、季节性干旱和盐碱)对植物的生存和生长具有重要影响[4]。水分、光照强度、土壤养分等是影响热带珊瑚岛植被恢复的重要生态因子。本研究表明,移栽至热带珊瑚岛地区的21 种植物的SLA 显著降低,叶片厚度和LDMC 显著增加,表明热带珊瑚岛的极端环境促使植物将更多的能量用于叶片的构建,降低SLA 以提高对胁迫的适应性。其余植物形态学特征未有显著变化,主要通过改变栅栏组织厚度、海绵组织厚度等叶片解剖结构特征,以适应热带珊瑚岛的胁迫环境。
已有研究表明,SLA 较小的植物,叶片通常较厚,具有较强的贮水能力,能够更好地适应干旱环境[27–28]。本研究中种植于热带珊瑚岛植物的平均SLA 为146 cm2/g,低于干旱地带植物SLA 平均值158 cm2/g[29],表明移栽后的植株经过一段时间的适应后抗旱能力有所提高,能够通过降低SLA 较好地适应热带珊瑚岛土壤保水能力差和季节性干旱等不利条件。LDMC 常被用于资源利用分类轴上植物种类的划分定位,不仅比较稳定且易于测定[19,30–31]。研究表明适应性较强的植物LDMC 通常较高,耐旱树种新疆杨(Populus alba var. pyramidalis)的LDMC[(420.378±68.046) mg/g]极显著高于二白杨[P. gansuensis, (301.107±32.647) mg/g]和小叶杨[P. simonii,(287.678±72.055) mg/g][32]。本研究中,移栽至热带珊瑚岛植物的LDMC 均显著高于生长于海南文昌的,表明其对热带珊瑚岛养分贫瘠的恶劣生境适应性较强。
植物叶片性状的变化并不是孤立的,而是紧密相关的。SLA 与LDMC 常用于综合反映植物对资源的利用能力及策略[31,33]。一般而言,采取资源获取型策略的植物通常具有较高的SLA 和较低的LDMC,叶片寿命较短但光合能力较强,养分循环较快,适应于养分资源丰富和湿润的环境;而采取资源保守型利用策略的植物通常具有较低的SLA 和较高的LDMC,叶片寿命较长且具有很好的保水能力,养分循环较慢,能更好地适应养分贫瘠和干旱的环境[33–34]。本研究中的热带珊瑚岛养分贫瘠,降水分布不均,存在严重的季节性干旱。相较于文昌苗圃的植物,生长于热带珊瑚岛的木本植物SLA 显著降低,LDMC显著提高,说明热带珊瑚岛木本植物通过不断权衡资源配置,采取慢速投资-收益的资源获取策略,将更多的资源投入到叶片构建中,提高保水能力,有效防止水分流失和维持渗透压[31]。这是木本植物适应热带珊瑚岛土壤保水能力差和季节性干旱环境的结果,也是其在干旱环境中长期生存的保水策略。而草本植物SLA 的降低并不显著,说明其可能更倾向于采取快速投资-收益策略以应对热带珊瑚岛的胁迫环境。
以往的诸多研究均表明,SLA 与LDMC 间呈现负相关关系[19,35–37]。随着LDMC 的增加,叶组织密度增加,从而导致SLA 降低。这也体现出植物在长期进化过程中,通过自身不同功能间的调整权衡, 形成一系列组合性状以适应外界环境的变化[32]。本研究结果与其一致,生长于海南文昌地区的木本植物及移栽至热带珊瑚岛地区的木本植物SLA 和LDMC 均呈显著负相关(P<0.05)。而草藤本植物的SLA 与LDMC的负相关关系在两地均不显著(P>0.05)。这可能是由于相较SLA,LDMC 受叶片含水量(LWC)的影响较大[38–39]。不同植物对水分胁迫的适应方式不同, 叶片性状对环境变化的响应是差异性的[29,40]。肉质和硬化是植物叶片适应低水分可利用性的不同方法,硬叶植物和肉质植物的SLA 均较低,但硬叶植物的LWC 较低,表现出相对较强的耐失水能力,而肉质植物的LWC 较高,表现出相对较强的保水能力[18,31]。在热带珊瑚岛的干旱胁迫下,不同物种间LWC 存在差异,这导致LDMC 在计算时存在较大的波动区间,从而削弱了LDMC 与SLA 之间的相关性[29,41]。
3.2 植物对热带珊瑚岛强光环境的适应
光是植物生长发育的必需资源,光合作用是大多数维管植物唯一的碳吸收机制,而水分亏缺在很大程度上限制光合作用的进行[42]。热带珊瑚岛光照强烈且干旱严重,对植物的光合作用以及生长发育存在较大影响。
相较于文昌苗圃的植物,移栽至热带珊瑚岛的木本植物叶片厚度增加,栅栏组织和海绵组织更加发达,栅栏组织与海绵组织的比值呈下降趋势。表明木本植物叶片受热带珊瑚岛高温、强光照的环境胁迫,叶肉形成一定的适应性,具有较高的光合潜能和光能利用效率[22]。而发达的栅栏组织以及排列紧密的海绵组织既可避免干旱地区强烈光照对叶肉细胞的灼烧,又可有效利用衍射光进行光合作用[10],是植物适应强光生境的结构基础[11]。
此外,气孔在植物光合作用过程中也起着重要作用,较小的气孔面积和较高的气孔密度有利于提高水分利用效率,减少植物光合过程中的蒸腾失水[43]。对青藏高原地区25 种耐旱树种的研究显示,胡颓子(Elaeagnuspungens)等灌木的气孔密度为57.27~360.05 n/mm2[44]。而本研究中移栽至热带珊瑚岛的植物平均气孔密度仅为279.58 n/mm2,密度较小且大小一般,有利于植物在提高光合能力的同时避免因蒸腾作用而过度失水[14],体现了植物光合作用与蒸腾作用之间的协同关系。
3.3 植物对热带珊瑚岛干旱胁迫的适应
植物适应干旱环境的机制包括气孔调节和非气孔调节两种方式。气孔通过生理学途径对蒸腾作用实现的调节作用统称为气孔调节,主要表现在气孔特征构造差异和气孔对环境诱导的反馈机制上[45]。非气孔调节则主要通过改变叶片导水率、叶肉组织、比叶面积等来实现[4]。干旱胁迫下,叶片的形态结构会发生变化,主要朝着保持水分吸收和减少水分丧失两个方面发展以维持体内的水分平衡[46]。李芳兰等[47]对白刺花(Sophoradavidii)响应干旱胁迫的研究表明,叶肉组织厚度的变化与植物的生长及水分利用间关系密切。发达的机械组织与极端干旱生境之间存在着明显的相关性,能够缓解干旱胁迫造成的损伤和失水的不利影响。从叶片形态解剖学特征来看,移栽至热带珊瑚岛的木本植物叶片厚度、栅栏组织厚度以及海绵组织厚度显著增加。这表明移栽至热带珊瑚岛的木本植物表现出明显的耐旱特征[48],能够通过增加叶片厚度和叶肉组织等非气孔调节方式来防止过度失水,提高水分利用效率,从而在热带珊瑚岛干旱等不利条件下保持正常的生长发育。
此外,本研究结果还表明,移栽至热带珊瑚岛的木本植物和草藤本植物在干旱胁迫下生长良好,而叶片气孔数量和密度未发生显著变化。前人[43]研究表明,适度的水分亏缺增加植物叶片的气孔密度,而重度水分胁迫使气孔密度降低,气孔调节能力下降。韩刚等对西北干旱半干旱地区6 种抗旱植物的研究发现,抗旱性强的物种气孔密度低[49]。因此,本试验结果中气孔调节失效可能是由于热带珊瑚岛地区的季节性干旱所致,即气孔调节需要在一定的土壤水分范围内,过度干旱时气孔的调节能力下降[50],植物通过非气孔调节的方式来提高抗旱能力以适应热带珊瑚岛干旱的气候条件。为更深入的探究植物对热带珊瑚岛干旱胁迫的适应,未来需要进一步对气体交换参数进行研究。
相较于文昌苗圃植物,移栽至热带珊瑚岛的草本植物除干物质含量增加外其他性状均无显著改变;而木本植物则采取慢速投资-收益的资源获取策略,将更多的资源投入到叶片构建中。热带珊瑚岛木本植物的比叶面积、叶片干物质含量显著下降、叶片厚度显著增加、海绵组织更厚且栅栏组织排列更紧密,表现出明显的旱生结构和阳生结构,这些形态解剖特征使植物能够更为有效地利用珊瑚岛生境中的光照和水分资源;采取非气孔调节方式,对珊瑚岛干旱、强光、高盐碱度的胁迫生境表现出良好的生态适应性。因此,从水分胁迫和光照胁迫的角度考虑,相对于草本植物,本研究选择的19 种木本植物可通过自身调节充分适应热带珊瑚岛的逆境胁迫,研究结果对我国热带珊瑚岛植被恢复和重建提供物种选择具有重要的理论和实践价值。
