枸杞深色有隔内生真菌定殖规律及其与土壤因子的相关性
2023-12-20王若彤陈思杰鞠明岫闫思远李金顾沛雯
王若彤 陈思杰 鞠明岫 闫思远 李金 顾沛雯


摘 要 利用采自宁夏银川市金凤区和西夏区两个枸杞栽培区0~20、20~40和40~60 cm共3个土层深度的枸杞根系和根围土样,研究枸杞 (Lycium barbarum)根系深色有隔内生真菌 (dark septate endophytes,DSE)的定殖规律及其与土壤因子的相关性,以期为充分利用DSE资源和基于生物技术促进荒漠植被恢复和生态改良提供依据。结果表明,DSE可定殖于枸杞根系形成深色、有隔菌丝,微菌核多呈聚合型和离散型。两样地DSE定殖率差异显著,金凤区样地DSE平均总定殖率(73.27%)显著高于西夏区样地 (23.54%);不同土层DSE定殖率有差异,两样地DSE总定殖率、菌丝定殖率和微菌核定殖率最大值均出现在20~40 cm土层。两样地不同土层深度间土壤因子的分布差异显著,主成分分析显示pH、碱解氮、速效磷、有机质、碱性磷酸酶和蔗糖酶能综合反映银川市两枸杞样地根际土壤养分状况。方差分解分析结果表明,DSE定殖受到样地和土壤因子的共同影响。相关性分析表明,DSE定殖在两样地间存在较大差异,金凤区样地DSE总定殖率与有机质显著正相关,与速效钾极显著负相关;DSE菌丝定殖率与有机质、速效磷和碱解氮显著正相关,与速效钾和pH显著负相关。西夏区样地DSE总定殖率和微菌核定殖率与pH极显著正相关;DSE菌丝定殖率与有机质、速效磷和脲酶显著负相关。上述结果说明,枸杞根系能被DSE侵染,DSE定殖具有明显的垂直空间异质性,并与土壤因子密切相关。
关键词 深色有隔内生真菌;枸杞;定殖状况;土壤因子
枸杞(Lycium barbarum)属茄科(Solanaceae)多年生落叶灌木,是中国传统的药食同源植物,主要生长在中国西北干旱地区[1-2]。枸杞作为中药材和重要的保健食品在中国已有上千年的历史,其果、叶、根、花皆具有较高的药用价值和保健效果,现代药理研究表明枸杞具有调节免疫、抗氧化、抗癌抗肿瘤等功效[3]。枸杞具有生态适应性强、抗寒、抗旱、耐贫瘠、耐盐碱等特点,是理想的生态和经济兼用的树种[4]。
深色有隔内生真菌(dark septate endophytes, DSE)是指定殖于植物根内的土壤真菌,其特点是寄居在健康植物根表皮、皮层乃至维管组织细胞内或细胞间隙,形成深色有隔菌絲、微菌核或泡囊结构,促进植物生长但不会引起典型的病理学特征[5]。研究发现,DSE宿主范围非常广泛,其宿主植物超过110科320属600余种[6],如绝大多数农作物、牧草、莎草科、十字花科、藜科等植物根系均有定殖[5]。DSE可能在生态学功能方面与菌根真菌相似,并对不同生态系统至关重要[5]。He等[7]发现接种DSE可以促进根系及植株对氮和磷的吸收,增加植株的根长、表面积、生物量。Jin等[8]研究表明DSE显著降低烟草叶片中镉和铅含量,提高烟草的生物量和重金属耐受性。DSE还可作为植物病害的生物防治剂,Harsonowati等[9]得出DSE Cladophialophora chaetospira SK51在草莓根系定殖,与草莓枯萎病病原Fusarium oxysporum f.sp.fragariae竞争占据植物根系的生态位,对草莓枯萎病的防效达90.48%,并促进增长。胡丽杰等[10]已证明DSE在枸杞中具有丰富的生物多样性,可在枸杞根部皮层细胞内定殖并形成菌丝和形态各异的微菌核,能够促进枸杞生长,提高枸杞对炭疽病的抗性,具有良好的促生防病作用[11],但目前鲜见有关枸杞根部DSE定殖状况与土壤环境因子相关的报道。
本研究通过对宁夏银川市金凤区和西夏区两个不同枸杞栽培区枸杞根系样品和根际土壤样品的采集,系统调查两个不同栽培区枸杞根系内DSE的定殖情况,同时测定各栽培区样地的土壤理化性质及酶活;分析DSE各结构定殖规律与土壤因子之间的相关性,旨在阐明影响枸杞根系DSE定殖和分布的主要土壤因子,为DSE资源的充分利用和进一步产业化应用提供依据。
1 材料与方法
1.1 材 料
于2020年8月采集枸杞根系样品和根围土土壤样品。选取宁夏银川市金凤区森淼枸杞园和西夏区红玛瑙枸杞园为两个典型采样地,两地面积均为3.33 hm2。金凤区森淼枸杞园(海拔 1 117 m,38°25′10″N,106°10′25″E),土壤类型为风沙土,该样地地势平坦,周边绿化植被较多,田间管理如灌水、施肥等良好;西夏区红玛瑙枸杞园(海拔1 127 m,38°31′31″N,106°4′20″E),土壤类型为砾质灰钙土,该样地地势较陡,植被类型单一,田间管理如灌水、施肥等频次较少。枸杞品种为‘宁杞5号,树龄8 a,其根系垂直分布在地下 0~60 cm的土层深度。本试验在每个采样地设置3个小样方(100 m×100 m),小样方间隔大于1 000 m,呈棋盘式。在每个小样方采用对角线采样法,随机选择5株长势良好的健康枸杞植株,株距大于100 m。在距枸杞植株主干东西南北4个方位水平方向0~30 cm范围内,除去其枯枝落叶层挖土壤剖面,按0~20 cm、20~40 cm和40~60 cm 3个土层采集土样和根样,每土层各1 kg,将其装于隔热性能良好的塑封袋中,记录采样时间、地点等信息,及时带回实验室进行处理。将每个小样方中同一土层的土样和根样均匀混合,过2 mm筛,随机挑选50~60个根段(长度为5~ 6 cm,直径小于2 mm),保存于FAA固定液(甲醛、冰醋酸及75%酒精体积比为1∶1∶18)中,用于DSE结构和定殖状况观测。筛下的土样经自然风干后,过1 mm筛,用于土壤因子测定。两个样地共计18份根样和18个土样。
1.2 试验方法
1.2.1 DSE定殖观察 参考Phillips等[12]的染色方法,从FAA固定液中取出根段,无菌水淋洗,将根段剪成1 cm后移入10% KOH中,90 ℃水浴1 h,乳酸酸化5~10 min。将根段酸化后经0.05%台盼蓝酸性甘油溶液(乳酸、甘油和水的体积比为3∶1∶1)染色5 min,乳酸甘油(乳酸和甘油的体积比为1∶1)脱色。随机选取50个根段,在Olympus BX53(奥林巴斯株式会社,日本)显微镜下观察DSE的侵染状况和定殖结构,并统计定殖率[13]。
DSE定殖率=(微菌核视野数+菌丝视野数)/总视野数×100%
1.2.2 土壤因子测定 土壤理化性质测定[14]:配制质量比为1∶5的土壤∶水悬浮液,采用PHS-25精密酸度计测量pH;重铬酸钾-硫酸氧化法测定土壤有机质;碱解扩散法测定碱解氮; 0.5 mol/L NaHCO3浸提-钼锑抗比色法测定速效磷;火焰光度法测定速效钾。土壤酶活测定[15]:采用靛酚蓝比色法测定土壤脲酶,用24 h后1 g土中NH3-N的毫克数表示其活性;采用3,5-二硝基水杨酸比色法测定蔗糖酶,用24 h后1 g土生成葡萄糖毫克数表示其活性;采用高锰酸钾滴定法测定过氧化氢酶;采用磷酸苯二钠比色法测定碱性磷酸酶,用24 h后1 g土中释放出的酚的毫克数表示其活性。
1.3 数据处理
将获得的数据使用 Microsoft Excel 2007软件整理;采用SPSS 25.0 进行单因素方差(OneWay ANOVA);采用Origin 2021通过主成分分析对土壤因子进行降维处理,确定影响两样地的主要土壤因素;采用美吉云平台生信工具VPA(Variance partitioning analysis)进行方差分解分析,分析样地、土层深度、土壤因子对DSE定殖的影响;采用Pearson对DSE定殖情况与土壤因子进行相关性分析。
2 结果与分析
2.1 DSE定殖结构
两样地健康枸杞根系DSE的定殖结构无明显差别。由图1观察结果表明,DSE可侵染并定殖于枸杞根系,菌丝有横隔,多呈深色,可侵入根系表皮和维管组织(图1-A~E),菌丝能够延伸至皮层细胞在胞内形成菌丝(图1-C),能够在皮层细胞间延伸形成菌丝网(图1-D和图1-E)。微菌核由细胞壁加厚的膨大细胞密集积聚形成,形状不一(图1-F~1-P)。根据其形态可分为聚合型和离散型。聚合型常见有叶状(图1-F~H)、脑状(图1-I)、锤状(图1-J);离散型可见带状(图1-K和图1-L)、链状(图1-M和图1-N)、串珠状(图1-O)和星点状(图1-P)。
2.2 DSE空间分布
由表1可知,同一样地不同土层,金凤区样地总定殖率在20~40 cm土层显著高于其他土层,菌丝定殖率在0~20 cm和20~40 cm土层显著高于40~60 cm土层,微菌核定殖率在20~40 cm土层高于其他土层;西夏区样地20~40 cm土层总定殖率和微菌核定殖率高于0~20 cm和 40~60 cm土层,20~40 cm土层菌丝定殖率显著高于0~20 cm土层。两个样地DSE总定殖率、菌丝定殖率和微菌核定殖率最大值出现在 20~40 cm土层。
同一土层不同样地,金凤区样地DSE平均总定殖率(73.27%)、菌丝定殖率(56.80%)和微菌核定殖率(32.36%)均显著高于西夏区样地的平均总定殖率(23.54%)、菌丝定殖率(20.1%)和微菌核定殖率(12.09%)。说明银川市两枸杞样地DSE的分布具有明显的垂直空间特异性,这可能与宿主植物根系分布有关。
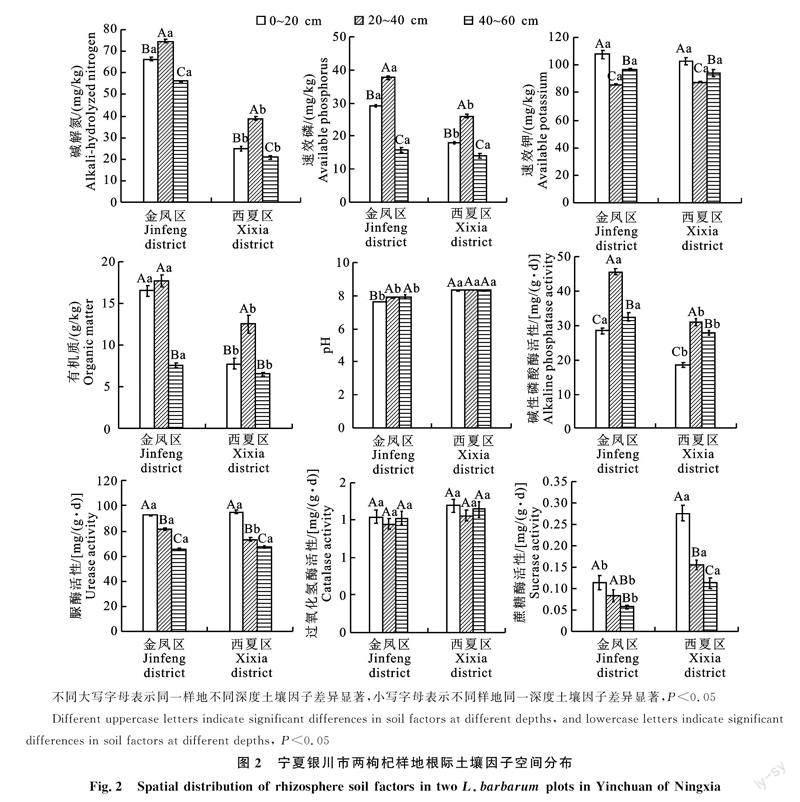
2.3 土壤因子空间分布
由图2可知,银川市金凤区和西夏区两枸杞樣地不同土层深度之间土壤因子差异显著。同一样地不同土层,碱解氮、速效磷、有机质和碱性磷酸酶的最大值均出现在20~40 cm,速效钾、脲酶、过氧化氢酶和蔗糖酶的最大值均出现在0~20 cm。碱性磷酸酶最小值均出现在0~20 cm,速效钾、过氧化氢酶最小值均出现在20~40 cm,碱解氮、速效磷、有机质、脲酶、蔗糖酶最小值均出[CM(21]现在40~60[KG*2]cm。碱解氮、速效磷、有机质、碱性磷酸酶随土层加深先升后降,速效钾、过氧化氢酶随土层加深先降后升,脲酶、蔗糖酶随土层加深呈下降趋势。碱解氮、速效钾、速效磷、碱性磷酸酶、脲酶在3个土层之间有显著差异,过氧化氢酶在3个土层之间无显著差异;金凤区样地有机质在 0~20、20~40 cm土层显著高于40~60 cm,西夏区样地有机质在20~60cm土层显著高于0~20、40~60 cm土层;金凤区样地蔗糖酶在0~20 cm土层显著高于40~60 cm土层,西夏区样地三个土层之间蔗糖酶有显著差异。金凤区pH的最大值出现在40~60 cm,最小值出现在0~20 cm,随土层加深呈上升趋势,且pH在20~40、40~60 cm土层显著高于0~20 cm;西夏区pH的最大值出现在40~60 cm,最小值出现在0~20 cm,随土层加深无明显变化规律。
同一土层不同样地,金凤区样地碱解氮、有机质、碱性磷酸酶均显著高于西夏区样地,金凤区样地pH和蔗糖酶均显著低于西夏区样地。金凤区速效磷在0~20 cm、20~40 cm土层均显著高于西夏区样地,在40~60 cm两样地无显著差异;金凤区脲酶在20~40 cm土层显著高于西夏区样地,在0~20 cm、40~60 cm土层两样地无显著差异。速效钾和过氧化氢酶在两个样地之间无显著差异。说明银川市两枸杞样地土壤因子的分布表现出明显的空间异质性。
2.4 土壤因子主成分分析
对pH、碱解氮、速效磷、速效钾、有机质、碱性磷酸酶、过氧化氢酶、脲酶、蔗糖酶9个土壤因子进行主成分分析,按照相关矩阵特征值大于1,方差累计贡献率大于75%的原则,选出3个主成分。分析结果如表2,提取的3个主成分方差累计贡献率达85.56%,能基本反映全部土壤因子的指标信息。第一主成分中,pH、碱解氮、速效磷、有机质、碱性磷酸酶载荷较大(权重>0.800);第二主成分中,蔗糖酶载荷较大(权重为0.943);第三主成分中,速效钾载荷较大(权重为0.907)。第一主成分(PC1)所占信息量达48.494%,第二主成分(PC2)所占信息量为21.841%,两者的方差贡献率均较大,且由图3可知,第一主成分(PC1)和第二主成分(PC2)中的方差贡献率最大,分别可以解释总方差的48.494%和21.841%,累计解释总方差的70.335%。所以pH、碱解氮、速效磷、有机质、碱性磷酸酶和蔗糖酶能综合反映银川市两枸杞样地根际土壤养分状况。
2.5 不同因子对DSE真菌定殖影响的方差分解分析
将不同样地、土层深度、土壤因子对枸杞根系DSE的定殖率进行方差分解分析,定量评估不同因子对枸杞根系DSE定殖率的贡献率[16]。由图4可知,不同样地、土层深度和土壤因子对DSE定殖的总解释量为76.9%,其中土壤因子与样地对DSE定殖的共同解释量为75.3%,说明宁夏银川市枸杞样地DSE定殖主要受到样地和土壤因子的共同影响。
2.6 DSE定殖与土壤因子相关性分析
由表3 Pearson相关性分析表明,DSE定殖在两样地间存在较大差异。金凤区样地枸杞根系[CM(21]DSE总定殖率与有机质显著正相关,与速效钾极显著负相关;DSE菌丝定殖率与有机质、速效磷和碱解氮显著正相关,与速效钾和pH显著负相关。西夏区样地枸杞根系DSE总定殖率和微菌核定殖率与pH极显著正相关;DSE菌丝定殖率与有机质、速效磷和脲酶显著负相关。对比不同样地可知,碱解氮、速效磷、有机质等土壤因子可以影响两样地DSE的定殖。
3 讨 论
3.1 DSE定殖与空间分布
DSE宿主范围和生态分布极广,定殖于植物根组织后不会引起明显的病理学特征,它寄生于植物根系或与其共生,作用于宿主植物并对生态系统的稳定和生产力造成一定影响[17]。研究结果表明,宁夏银川市两样地的枸杞根系可被DSE侵染,并形成深色、有隔菌丝和典型的微菌核结构。菌丝可通过变窄穿过植物细胞壁供DSE与植物进行营养物质交换[17];微菌核由菌丝膨大加厚堆积而成,可作为DSE的繁殖结构[18]。本研究得出枸杞根系DSE总定殖率在23.54%~ 73.27%,平均定殖率为48.41%,说明DSE真菌可与其宿主枸杞根系形成良好的共生关系。李雪梦[19]研究得到DSE在枸杞根系内所形成的不同定殖结构的定殖率间有差异,表现为菌丝定殖率高于微菌核定殖率,本研究得到了相同的结论。
DSE定殖率在不同土层深度间具有明显的垂直空间特异性[20]。蔚杰等[21]发现构树根系在0~20 cm土层的DSE总定殖率显著高于20~40 cm土层。王少杰等[18]发现蒙古沙冬青DSE的定殖率在0~20 cm土层最大。孙茜等[22]发现白刺、柠条锦鸡儿、无叶沙拐枣、红花锦鸡儿等蒙古沙冬青的伴生植物DSE定殖率最大值在0~20 cm土层,且定殖率均表现为浅土层高于深土层。本研究同样发现DSE定殖率在不同土层间有差异,但其中20~40 cm土层DSE定殖率高于0~20 cm和40~60 cm土层,这可能是与宿主植物根系分布有关。构树、蒙古沙冬青、沙蒿及白刺等植物根系入土浅,主要分布在0~20 cm土层,可能是浅土层土壤表面落叶及腐殖质积累较多,土壤通气性佳,微生物资源多样,对DSE生长有利。而枸杞根系侧、须根分布的高密度区是在距地表20~40 cm的较深土层内,生物量最大[23],可为DSE提供更多的寄居环境。因此,根系的空间分布可能导致植物DSE定殖不同。DSE较高定殖可使根系生物量显著提高,对根系生长范围扩大有益[24]。
DSE广泛分布于各类生境,特别是在胁迫环境及受干扰严重区域,其组成在不同生境中有明显差别[5]。余鑫[25]分析认为在重金属矿区中芦竹的DSE定殖率和生态环境密切相关,相比于水生环境,陆生环境下DSE定殖率显著较大,且能达到水生环境芦竹根内定殖率的3倍多。Casanova-Katny等[26]通过分析智利中部安第斯山脉不同海拔中23 种植物共生真菌的定殖情况,得到DSE侵染程度随海拔升高而增大。李宝库等[27]也得到了一致的結论。本研究发现金凤区样地DSE总定殖率、菌丝定殖率与微菌核定殖率均显著高于西夏区样地,可能是因为金凤区的风沙土表层营养物质流动性大,根系较浅的植株只能在浅土层获取有限的营养物质,根系较深的植物能够利用更深土层养分,而在西夏区高肥力土壤中DSE的矿化作用[28]下降,植物可以直接从土壤吸收所需营养物质,使DSE与植物之间的互动联系降低,可能造成西夏区样地DSE的定殖率下降。由此可见,DSE的定殖可能受到生境和宿主根部结构的双重影响。
3.2 DSE与土壤因子的关系
土壤理化性质对微生物的群落结构有重要影响[29]。土壤pH可影响植物对土壤元素的有效利用,大多数植物的最大养分利用率pH 为 6.0~7.5,土壤pH升高反而会限制土壤养分的吸收[30]。本研究中,金凤区DSE定殖与土壤pH在7.5~8.2显著负相关,当pH较高时,可能会通过促进土壤养分的矿化而在某种程度抑制DSE的定殖。土壤有机质作为土壤中各种营养元素尤其是氮、磷的重要来源,其含量高低直接影响了土壤肥力和土壤质量[31]。Li等[32]发现土壤有机质与菌丝、微菌核定殖呈显著正相关。Hou等[20]也得出微菌核定殖与土壤有机碳(SOC)呈显著正相关。本研究发现金凤区样地枸杞根系的DSE总定殖率、菌丝定殖率与土壤有机质呈显著正相关,与前人结论一致。而西夏区DSE菌丝定殖率与有机质呈显著负相关可能也与pH过高,抑制了DSE的定殖相关。张娟等[33]发现沙鞭群落空地DSE总定殖率、菌丝定殖率和微菌核定殖率与碱解氮、速效磷呈极显著正相关,与本试验金凤区所得结果一致。氮和磷是植物生长的必要营养元素,作为植物生长最重要的限制资源之一,两者影响植物从代谢到生物量生产和生长的各个层面,DSE真菌则可在宿主根部定殖促进宿主对氮素和磷的吸收[34]。
土壤酶作为土壤评价体系中一项重要的生物学指标,是具有催化能力的“特殊有机体”,能够参与土壤新陈代谢并表示土壤养分转化能力的强弱[35]。土壤磷酸酶、脲酶、蔗糖酶等水解酶活性能够表征土壤碳氮磷等营养物质的循环情况,与土壤养分密切相关[36]。王少杰等[18]研究得出乌海、阿拉善左旗和民勤样地DSE菌丝定殖的主要土壤影响因子均有碱性磷酸酶。石经新[37]认为西北荒漠黑枸杞的菌丝定殖率、总定殖率和定殖强度与碱性磷酸酶、脲酶显著负相关。贺学礼等[38]通过揭示DSE在荒漠沙蒿根围的空间分布及其与土壤因子的相关程度,指出DSE定殖与脲酶呈负相关。葛佳丽等[17]认为不同根系来源样品DSE定殖率与脲酶存在极显著负相关性,与本研究西夏区枸杞根系DSE菌丝定殖率与脲酶显著负相关的结果一致。Xie等[39]认为DSE能够促进土壤氮和磷的循环利用是经过脲酶和磷酸酶活性升高间接作用实现的。脲酶参与的酶促反应可产生一种氮源氨,能被植物有效利用,其活性也体现出土壤将有机态氮转化为有效态氮及土壤供应无机氮的能力[40]。方差分解分析表明,土壤因子会影响宁夏银川市枸杞DSE的侵入和定殖。同时,DSE定殖与样地、土层深度、土壤因子(土壤理化性质和土壤酶活)的方差分解有23.1%的总方差未被解释,说明还有其他因子可能影响DSE定殖,有待进一步探索。
4 结 论
宁夏银川市金凤区和西夏区两枸杞样地根系均能被DSE定殖,可形成深色有隔菌丝、聚合型或离散型微菌核,具有明显的空间异质性。pH、碱解氮、速效磷、有机质、碱性磷酸酶和蔗糖酶能综合反映两枸杞样地根际土壤养分状况。DSE定殖受到样地和土壤因子的共同影响,并与土壤因子密切相关。说明DSE能与枸杞根系形成良好的共生关系,为充分利用DSE资源和生物技术促进荒漠植被恢复和生态改良提供依据。
参考文献 Reference:
[1]胥生荣,张恩和,马瑞丽,等.干旱胁迫及复水对耐旱枸杞水力学特性的影响[J].中国生态农业学报,2017,25(8):1190-1197.
XU SH R,ZHANG E H,MA R L,et al.Hydraulic characteristics of Lycium barbarum L.seedlings under drought stress and re-watering conditions[J].Chinese Journal of Eco-Agriculture,2017,25(8):1190-1197.
[2]朱娟娟,馬海军,张 琇,等.盐胁迫下解钾菌对枸杞幼苗的促生效应[J].应用生态学报,2021,32(4):1289-1297.
ZHU J J,MA H J,ZHANG X,et al.Effects of potassium-solubilizing bacteria promoting the growth of Lycium barbarum seedlings under salt stress[J].Chinese Journal of Applied Ecology,2021,32(4):1289-1297.
[3]如克亚·加帕尔,孙玉敬,钟烈州,等.枸杞植物化学成分及其生物活性的研究进展[J].中国食品学报,2013,13(8):161-172.
RUKEYA·JIAPAER,SUN Y J,ZHONG L ZH,et al.A review of phytochemical composition and bio-active of Lycium barbarumfruit (Goji)[J].Journal of Chinese Institute of Food Science and Technology,2013,13(8):161-172.
[4]赵 琴,潘 静,曹 兵,等.气温升高与干旱胁迫对宁夏枸杞光合作用的影响[J].生态学报,2015,35(18):6016-6022.
ZHAO Q,PAN J,CAO B,et al.Effects of elevated temperature and drought stress on photosynthesis of Lycium barbarum[J].Acta Ecologica Sinica,2015,35(18):6016-6022.
[5]刘茂军,张兴涛,赵之伟.深色有隔内生真菌(DSE)研究进展[J].菌物学报,2009,28(6):888-894.
LIU M J,ZHANG X T,ZHAO ZH W.Advances in the research of dark septate endophytes[J].Mycosystema,2009,28(6):888-894.
[6]ARI J,JAMES M T.Dark septate endophytes:a review of facultative biotrophic root-colonizing fungi[J].New Phytologist,1998,140(2):295-310.
[7]HE C,WANG W Q,HOU J L.Characterization of dark septate endophytic fungi and improve the performance of liquorice under organic residue treatment[J].Frontiers in Microbiology,2019,10:1364.
[8]JIN H Q,LIU H B,XIE Y Y,et al.Effect of the dark septate endophytic fungus Acrocalymma vagum on heavy metal content in tobacco leaves[J].Symbiosis,2018,74(2):89-95.
[9]HARSONOWATI W,MARIAN M,SURONO,et al.The effectiveness of a dark septate endophytic fungus,Cladophialophora chaetospira SK51,to mitigate strawberry Fusarium wilt disease and with growth promotion activities[J].Frontiers in Microbiology,2020,11:585.
[10]胡丽杰,闫思远,孙牧笛,等.宁夏枸杞深色有隔内生真菌的侵染特征及多样性[J].微生物学通报,2019,46(11):2973-2984.
HU L J,YAN S Y,SUN M D,et al.Infection characteristics and diversity of dark septate endophytic fungi isolated from Lycium barbarum in Ningxia[J].Microbiology China,2019,46(11):2973-2984.
[11]胡丽杰,闫思远,李嘉泓,等.枸杞内生真菌对胶孢炭疽菌Colletotrichum gloeosporioides的拮抗作用及生防潜力[J].植物保护,2020,46(1):125-133.
HU L J,YAN S Y,LI J H,et al.Antagonism and control potential of endophytic fungi from Lycium barbarum against Colletotrichum gloeosporioides[J].Plant Protection,2020,46(1):125-133.
[12]PHILLIPS J M,HAYMAN D S.Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection[J].Transactions of the British Mycological Society,1970, 55(1):158-161.
[13]曹冠华,张 雪,顾 雯,等.不同产地滇黄精丛枝菌根真菌、深色有隔内生真菌定殖调查及与主要功效成分含量相关性分析[J].中草药,2019,50(16):3930-3936.
CAO G H,ZHANG X,GU W,et al.Colonization investigation of arbuscular mycorrhizal fungi (AMF) and dark septate endophytes (DSE) in roots of Polygonatum kingianum and their correlations with content of main functional components in rhizomes[J].Chinese Traditional and Herbal Drugs,2019,50(16):3930-3936.
[14]鮑士旦.土壤农化分析[M].第3版.北京:中国农业出版社,2008.
BAO SH D.Agrochemical Analysis of Soil [M].3rd ed Beijing:Chinese Agriculture Press,2008.
[15]关松荫.土壤酶及其研究法[M].北京:农业出版社,1986.
GUAN S Y.Soil Enzyme and Its Research Method[M].Beijing:Agriculture Press,1986.
[16]赵 昕,高慧利,龙俊萌,等.西北沙区梭梭根系深色有隔内生真菌等微生物时空分布及对根际土壤环境的响应[J].菌物学报,2021,40(10):2716-2734.
ZHAO X,GAO H L,LONG J M.Spatial and temporal distribution and the response to rhizosphere soil environment of dark septate endophyte and other microorganisms in roots of Haloxylon ammodendron in sand area of Northwest China[J].Mycosystema,2021,40(10):2716-2734.
[17]葛佳丽,张琳琳,李欣玫,等.极旱荒漠植物深色有隔内生真菌定殖及其与土壤因子的相关性[J].河北大学学报(自然科学版),2018,38(1):73-82.
GE J L,ZHANG L L,LI X M,et al.Colonization of dark septate endophytes in roots of different plants and its correlation with soil factors in extreme arid desert environment[J].Journal of Hebei University (Natural Science Edition),2018,38(1):73-82.
[18]王少杰,侯力峰,解琳琳,等.蒙古沙冬青根系DSE定殖与土壤因子典范对应分析[J].河北大学学报(自然科学版),2017,37(5):497-507.
WANG SH J,HOU L F,XIE L L,et al.CCA analysis of colonization of DSE in the rhizosphere of Ammopiptanthus mongolicus in relation to soil factors[J].Journal of Hebei University (Natural Science Edition),2017,37(5):497-507.
[19]李雪梦.四子王旗荒漠草原植物深色有隔内生真菌(DSE)研究[D].呼和浩特:内蒙古农业大学,2019.
LI X M.Study on dark septate endophytes of defferent plants in the desert steppe of Siziwang banner[D].Hohhot:Inner Mongolia Agricultural University,2019.
[20]HOU L F,HE X L,LI X,et al.Species composition and colonization of dark septate endophytes are affected by host plant species and soil depth in the Mu Us sandland,northwest China[J].Fungal Ecology,2019,39:276-284.
[21]蔚 杰,贺学礼,赵丽莉,等.河北峰峰矿区深色有隔内生真菌定殖及群落组成[J].菌物研究,2018,16(4):228-238.
YU J,HE X L,ZHAO L L,et al.Colonization and community composition of dark endophytic fungi in Fengfeng mining area of Hebei[J].Journal of Fungal Research,2018,16(4):228-238.
[22]孙 茜,贺 超,贺学礼,等.沙冬青与伴生植物深色有隔内生真菌定殖规律及其与土壤因子的相关性[J].植物生态学报,2015,39(9):878-889.
SUN Q,HE CH,HE X L,et al.Colonization of dark septate endophytes in roots of Ammopiptanthus mongolicusand its associated plants as influenced by soil properties [J].Chinese Journal of Plant Ecology,2015,39(9):878-889.
[23]占玉芳,鲁延芳,甄伟玲,等.河西灌区枸杞根系空间分布特征研究[J].水土保持研究,2016,23(4):133-137
ZHAN Y F,LU Y F,ZHEN W L,et al.Spatial distribution characteristics of root system of Lycium barbarum L.in Hexi irrigation area[J].Research of Soil and Water Conservation,2016,23(4):133-137.
[24]NEWSHAM K K.A meta-analysis of plant responses to dark septate root endophytes[J].New Phytol,2011, 190(3):783-793.
[25]余 鑫.云南募乃鉛锌矿区陆生和水生芦竹DSE多样性及其功能初探[D].昆明:云南大学,2016.
YU X.Diversity and functions of dark septate endophytes (DSE) colonizing the aquatic and terrestrial Arundo donax in Munai Pb/Zn slag heap,Yunnan,SW China[D].Kunming:Yunnan University,2016.
[26]CASANOVA-KATNY M A,TORRES-MELLADO G A,PALFNER G,et al.The best for the guest:high Andean nurse cushions of Azorella madreporica enhance arbuscular mycorrhizal status in associated plant species[J].Mycorrhiza,2011, 21(7):613-622.
[27]李宝库,贺学礼,吴伟家.不同海拔蒙古沙冬青根系深色有隔内生真菌(DSE)定殖的空间变化[J].江苏农业科学,2016,44(5):502-505.
LI B K,HE X L,WU W J.Spatial changes of dark septate endophytic fungi (DSE) colonization in roots of Ammopiptanthus mongolicus at different altitudes[J].Jiangsu Agricultural Sciences,2016,44(5):502-505.
[28]HOEKSEMA J D,CHAUDHARY V B,GEHRING C A,et al.A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi[J].Ecology Letters,2010,13(3):394-407.
[29]王蓥燕,王子芳,黄 容,等.缙云山不同森林植被下土壤微生物群落结构特征研究[J].土壤学报,2019,56(5):1210-1220.
WANG Y Y,WANG Z F,HUANG R,et al.Characterization of soil microbial community structure as affected by vegetation in Jinyun mountain[J].Acta Pedologica Sinica,2019,56(5):1210-1220.
[30]ZHAO J,DONG Y,XIE B B,et al.Effect of annual variation in soil pH on available soil nutrients in pear orchards[J].Acta Ecologica Sinica,2011,31(4):212-216.
[31]吳海燕,金荣德,范作伟,等.基于主成分和聚类分析的黑土肥力质量评价[J].植物营养与肥料学报,2018,24(2):325-334.
WU H Y,JIN R D,FAN Z W,et al.Assessment of fertility quality of black soil based on principal component and cluster analysis[J].Journal of Plant Nutrition and Fertilizers,2018,24(2):325-334.
[32]LI B K,HE X L,HE C,et al.Spatial dynamics of dark septate endophytes and soil factors in the rhizosphere of Ammopiptanthus mongolicus in Inner Mongolia,China[J].Symbiosis,2015,65(2):75-84.
[33]张 娟,贺学礼,赵丽莉,等.荒漠土壤因子和DSE定殖对克隆植物入侵的响应[J].生态学报,2015,35(4):1095-1103.
ZHANG J,HE X L,ZHAO L L,et al.Responses of desert soil factors and dark septate endophytes colonization to clonal plants invasion[J].Acta Ecologica Sinica,2015, 35(4):1095-1103.
[34]VERGARA C,ARAUJO K E C,ALVES L S,et al.Contribution of dark septate fungi to the nutrient uptake and growth of rice plants[J].Brazilian Journal of Microbiology,2018,49(1):67-78.
[35]王 锐,闫鹏科,马婷慧,等.行内生草对土壤微环境和酿酒葡萄品质的影响[J].干旱地区农业研究,2020,38(3):195-203.
WANG R,YAN P K,MA T H,et al.Effects of intra-row planted grass on soil microenvironment and wine grape quality[J].Agricultural Research in the Arid Areas,2020,38(3):195-203.
[36]刘红梅,周广帆,李 洁,等.氮沉降对贝加尔针茅草原土壤酶活性的影响[J].生态环境学报,2018,27(8):1387-1394.
LIU H M,ZHOU G F,LI J,et al.Effects of nitrogen deposition on soil enzyme activities of stipa baicalensis steppe[J].Ecology and Environmental Sciences,2018,27(8):1387-1394.
[37]石经新.西北荒漠黑枸杞深色有隔内生真菌时空分布和促生抗旱研究[D].河北保定:河北大学,2021.
SHI J X.Spatiotemporal distribution and drought promotion of dark septate endophytes fungi in Lycium ruthenicum in Northwest China [D].Baoding Hebei:Hebei University,2021.
[38]贺学礼,王银银,赵丽莉,等.荒漠沙蒿根围AM真菌和DSE的空间分布[J].生态学报,2011,31(3):812-818.
HE X L,WANG Y Y,ZHAO L L,et al.Spatial distribution of arbuscular mycorrhizal fungi and dark septate endophytes in the rhizosphere of Artemisia sphaerocephala from Inner Mongolia desert[J].Acta Ecologica Sinica,2011,31(3):812-818.
[39]XIE L L,HE X L,WANG K,et al.Spatial dynamics of dark septate endophytes in the roots and rhizospheres of Hedysarum scoparium in northwest China and the influence of edaphic variables[J].Fungal Ecology,2017,26:135-143.
[40]徐浩博,贺学礼,许 伟,等.蒙古沙冬青根围丛枝菌根和深色有隔内生真菌的空间分布[J].贵州农业科学,2013,41(12):105-109.
XU H B,HE X L,XU W,et al.Spatial distribution of arbuscular mycorrhizal fungi and dark septate endophytes in the rhizosphere of Ammopiptanthus mongolicus[J].Guizhou Agricultural Sciences,2013,41(12):105-109.
Colonization of Dark Septate Endophytes Fungi in Lycium barbarum and Its Correlation with Soil Factors
WANG Ruotong, CHEN Sijie, JU Mingxiu, YAN Siyuan, LI Jin and GU Peiwen
Abstract In the present study, we investigated the colonization of dark septate endophytes (DSE) in the roots of Lycium barbarum and its correlation with soil factors, and collected the root and inter-root soil samples at three soil depths of 0-20 cm, 20-40 cm and 40-60 cm from cultivation area of L.barbarum in Jinfeng district and Xixia district of Yinchuan in Ningxia, so as to provide a basis for the full utilization of DSE resources and biotechnology-based promotion of desert vegetation restoration and ecological improvement.The results showed that DSE could colonize the root system of L.barbarum, form dark and septate hyphae, and generate polymeric and discrete microsclerotia.The rate of DSE colonization was significantly different between the two sample plots.The mean total colonization rate of DSE in Jinfeng (73.27%) was significantly higher than that in Xixia (23.54%).The DSE colonization rates varied among different soil layers.The maximum values of total DSE colonization rate, hyphae colonization rate and microsclerotia colonization rate were all found at the 20-40 cm soil layers of both sample plots.The distribution of soil factors differed significantly among different soil depths in the two samples.The principal component analysis showed that the rhizosphere soil nutrient status was affected comprehensively by pH, alkali hydrolyzable nitrogen, available phosphorus, organic matter, alkalin phosphatase and sucrase in two sample plots of Yinchuan.Variation partitioning revealed that the colonization of DSE fungi was affected by a combination of sample site and soil factors.The correlation analysis showed that DSE colonization differed significantly between the two samples.In the sample plot of Jinfeng district, the total DSE colonization rate had significantly positive correlation with organic matter, and significantly negative correlation with available potassium.The DSE hyphae colonization rate had significantly positive correlation with organic matter, available phosphorus and alkaline nitrogen, and significantly negative correlation with available potassium and pH.In the sampling site of Xixia, the total DSE colonization rate and the microsclerotia colonization rate had significantly positive correlation with pH, while the DSE hyphae colonization rate had significantly negative correlation with organic matter, available phosphorus and urease.The findings in this study indicated that the root system of L.barbarum can be infested by DSE, the colonization exhibits obvious vertical spatial heterogeneity and is closely related to soil factors.
Key words Dark septate endophytes; Lycium barbarum;Colonization status;Soil factors
Received2022-07-29Returned 2022-08-01
Foundation item National Natural Science Foundation of China(No.31460484);Key Research and Development Plan of China(No.2019YFD10025002).
First author WANG Ruotong, female,master student.Research area:biological control and mycological resources utilization.E-mail:wangruotong0729@163.com
Corresponding author GU Peiwen, female,Ph.D,professor.Research area:biological control and mycological resources utilization.E-mail:gupeiwen2013@126.com
(责任编辑:史亚歌 Responsible editor:SHI Yage)
收稿日期:2022-07-29修回日期:2022-08-01
基金项目:国家自然科学基金(31460484);国家重点研发计划(2019YFD10025002)。
第一作者:王若彤,女,硕士研究生,研究方向为生物防治与菌物资源利用。E-mail:wangruotong0729@163.com
通信作者:顧沛雯,女,博士,教授,研究方向为生物防治与菌物资源利用。E-mail:gupeiwen2013@126.com
