煤矿复垦区不同恢复年限沙棘人工林土壤细菌群落结构与功能预测
2023-12-14同晓冬杨妮秦家凤刘广全艾宁刘长海
同晓冬, 杨妮, 秦家凤, 刘广全, 艾宁,, 刘长海*
(1.延安大学生命科学学院, 延安 716000; 2.中国水利水电科学研究院, 北京 100038)
中国是煤炭开采量全球最高的国家[1],大量煤矿资源的开采,满足了经济建设需要的同时也严重扰动了区域的自然生态系统,尤其是采矿导致的土壤损毁、土壤污染及水土流失问题[2]。目前,矿产资源的开采所引发的生态问题已上升为世界问题,各国学者及政府对此给予了广泛的关注,并进行了深入研究[3-5]。中国在该方面的研究较晚且对矿区的修复率不足,这不利于区域社会经济发展和环境安全[6]。排土场是露天煤矿开采造成土壤损毁的主要表现形式之一,该区域土壤动植物多样性低下,土壤养分减少,土壤功能下降[7],因此对其进行复垦已成为矿区生态修复与重建的关键,对于矿区的可持续发展具有重要意义。
土壤复垦对土壤环境产生影响,进而推动土壤微生物群落的变化。土壤微生物是土壤生物区系中的重要组成部分,可参与有机物质的分解和矿化,腐殖质形成,土壤养分转化及循环,促进生物地球化学循环[8-9],是连接地上与地下部分的重要环节,向上促进植被的定植和生长,向下维持土壤的结构及理化性质的稳定性[10]。土壤细菌是土壤微生物中最丰富的部分,参与土壤系统中大多数的养分转化[11-12],对土壤微环境变化敏感,可短期内快速对环境变化产生反应,其群落结构和功能的变化可指示植被群落的演替和土壤肥力的改变[13-14],因此,应用微生物群落特征来评估矿区土壤健康及生态修复效果已成为中外学者研究的热点问题[15]。张小玲等[16]对不同复垦年限煤矸山的功能细菌进行研究,发现复垦年限的增加促进了功能菌总量的增加,速效磷、含水量和碱解氮是影响功能菌的主要因子。马守臣等[17]研究了山西长治司马矿煤矸石填埋场土壤微生物学特性的时空变异,表明各土层微生物数量和真菌、放线菌的丰度随复垦年限逐渐增加。Li等[18]评估了复垦对黄土高原不同植被类型土壤微生物及酶活性的影响,发现土壤有机碳是影响土壤微生物群落和酶活性的主要因素,微生物多样性在混交林土壤中最高,复垦对微生物丰度、多样性和组成的影响在各个植被类型存在差异。
Plassart等[19]对法国北部粉砂田研究表明,草地恢复可逐渐增加微生物量和微生物活性,对保持土壤状况有积极影响。陈熙等[20]对赣州-安远稀土弃废尾矿进行研究,结果显示,经不同的植被修复后,原尾矿土壤理化性质改善明显,土壤微生物结构发生改变且土壤微生物多样性、丰富度、均匀度均得到显著的提高。Garaiyurrebaso等[21]在矿区选择两个被重金属严重污染的植被场地,分析了使用几种不同改良剂5 a后的微生物生物量、多样性等指标,结果表明,造纸厂污泥与鸡粪混合处理后,镉、铅和锌的生物有效性降低幅度最大,并刺激了土壤微生物活性和多样性,土壤质量大幅提高。但以上研究多集中于植被对微生物生物量、群落组成及结构的影响,少见于植被不同恢复年限土壤微生物功能的研究,更鲜见于连续恢复年限对土壤微生物结构和功能的影响,而研究土壤微生物功能对于揭示其在土壤生态系统修复中的作用具有重要意义[22-23]。目前,随着分子生物技术的快速发展,PICRUSt作为一种预测细菌代谢功能的新算法已被广泛应用于海洋、湖泊、土壤领域的细菌功能分析中并取得了显著成效[24-25]。这是一种基于高通量测序程序,将得到的16S rRNA基因与数据库中已知代谢功能的细菌基因序列进行比对,从而对细菌进行预测的一种方法[26],为对细菌功能的深入认识提供了新途径。
基于此,本研究以鄂尔多斯聚鑫龙煤矿复垦区3~7 a恢复年限(连续年份)沙棘人工林为研究对象,采集研究区土壤样品,应用Illumina高通量测序技术和PICRUSt算法对土壤样品中的细菌群落结构、多样性和功能进行分析,并揭示土壤细菌与土壤理化性质之间的关系,以期为研究区矿区土地复垦与环境修复提供数据支撑与科学依据。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古自治区鄂尔多斯市的聚鑫龙煤矿(110°4′2″E,39°54′16″N),地势平坦,海拔约1 260 m,水蚀风蚀严重,属温带大陆性气候区,全年无霜期约135 d, 年均温7.5 ℃,年均降水量400 mm,集中于7—9月。研究区于2011年开始造林进行矿区生态修复建设,主要种植大果沙棘(HippophaerhamnoidesLinn.cv)。样地概况如表1所示。

表1 样地概况
1.2 样地设置与样品采集
在研究区选取恢复年限为3~7 a的沙棘林作为研究对象,并以撂荒草地做对照进行土壤样品的采集。各采样区布设20 m × 20 m的固定样地,每个样地设置1 m×1 m的3个样方,每个样方按“S”形5点法进行取样,如图1所示。用土钻(直径5 cm、高20 cm)进行垂直取样,取样深度为10 cm。将5份土样放在油布上混合均匀,去除其中的枯落物等杂物后过2 mm土筛,取2份土样,每份土样500 g,一份于无菌袋放入干冰盒内,将其置于-70 ℃冰箱中保存用于微生物DNA提取及高通量测序;另一份于塑封袋中带回实验室,自然风干后进行理化性质的测定。

图1 样地及采样基本情况图Fig.1 Sample plot and basic sampling situation diagram
1.3 样品测定
1.3.1 土壤基本特性
土壤理化性质的测定依照《土壤农化分析》[27]进行:土壤pH用酸度计法测定,有机质用重铬酸钾容量法,碱解氮采用碱解扩散法,全磷用钼锑抗法,速效钾用NH4OAC浸提-火焰光度法,土壤含水量依据《土壤物理性质测定方法》[28]中的环刀浸提法进行测定。
1.3.2 土壤细菌高通量测序
土壤样品总DNA的提取采用E-Z 96 Mag-Bind Soil DNA Kit(M5636-02,Omega,CA,USA)试剂盒进行,取0.5 g新鲜土样,按照试剂盒说明书进行具体操作。细菌的V3V4的扩增引物为338F(5′-ACTCCTACGGGAGGCAGCA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)。PCR的反应体系包括:DNA模板1 μL,上游引物和下游引物 (10 μmol/L) 各1 μL,Fast pfu DNA聚合酶0.25 μL,2 μL的dNTP(2.5 mmol/L),超纯水(ddH2O)14.75 μL,共25 μL。扩增条件为:98 ℃预变性5 min,28个循环(98 ℃ 30 s,52 ℃ 30 s,72 ℃ 45 s),最后72 ℃延伸5 min。用1.2%琼脂糖凝胶电泳检测PCR扩增产物,用磁珠对产物进行纯化后进行荧光定量,荧光试剂为Quant-iT PicoGreen dsDNA Assay Kit,定量仪器为Microplate reade (BioTek,FLx800)。根据荧光定量结果,按照每个样本的测序量需求,对各样本按相应比例进行混合。产物委托上海派森诺生物科技有限公司进行高通量测序。高通量测序得到的数据用Vsearch软件分析流程进行质量过滤,拼接和嵌合体的去除。对得到的高质量序列进行OTU类分析,基于OTU聚类的结果,之后用QIIME2 (2019.4)分析软件对物种分类及生物多样性指数进行分析。
1.4 数据分析
采用Excel 2010和SPSS 22.0软件对土壤理化性质及细菌群落组成数据进行处理,应用单因素方差分析(ANOVA)和多重比较法(Duncan)进行数据显著性差异分析。PICRUSt基因功能预测由派森诺生物公司进行,细菌群落冗余分析在软件Canoco 5.0中完成,其余图形绘制均采用Origin 2021和R4.1.2软件完成。
2 结果分析
2.1 土壤基本性质
不同恢复年限沙棘林土壤基本性质如表2所示,研究区土壤偏碱性,pH变化范围处于7.47~8.29;土壤有机质随恢复年限增长呈先减小后显著增大的趋势,恢复7 a 时土壤有机质最大,且高于草地有机质含量;碱解氮和全磷的平均值分别为9.31 g/kg和0.48 g/kg,二者的最大值都出现在恢复年限6 a;速效钾和水分的平均值分别为81.53 g/kg和6.76 g/kg,速效钾的最大值出现在最大恢复年限7 a,水分的最大值出现在恢复年限3 a。

表2 土壤理化指标
2.2 不同恢复年限土壤细菌群落结构
2.2.1 土壤细菌群落组成
不同恢复年限沙棘人工林土壤样品共检测到细菌29门、91纲、193目、305科和438属,在门分类水平上(图2),各样地以放线菌门(Actinobacteria)、变形菌门(Proteobacteria)为优势菌门,拟杆菌门(Bacteroidetes)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、浮霉菌门(Patescibacteria)、疣微菌门(Verrucomicrobia)、蓝藻门(Cyanobacteria)为常见菌门,以上菌门在各样地中总占比达到80%以上,但各个菌门分布存在差异。放线菌门在样地3 a中为第二优势菌门,而在其他样地均为第一优势菌门,且在各样地中的比例也存在差异,在5 a最高,7 a最低。
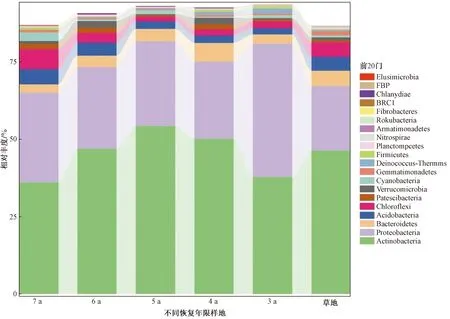
Actinobacteria为放线菌门; Proteobacteria为变形菌门; Bacteroidetes为拟杆菌门; Acidobacteria为酸杆菌门; Chloroflexi为绿弯菌门; Patescibacteria为浮霉菌门; Verrucomicrobia为疣微菌门; Cyanobacteria为蓝藻门;Genunatimonadetes为芽单胞菌门; Deinococcus-Thermus为异常球菌-栖热菌门; Firmicutes为厚壁菌门; Planctomycetes为浮霉菌门; Nitrospirae为硝化螺旋菌门; Armatimonadetes为装甲菌门; Rokubacteia为己科河菌门; Fibrobacteres为纤维杆菌门; Chlanydiae为衣原体门; Elusimnicrobia为迷踪菌门
图3为细菌属水平上丰度前20个属的相对丰度。就整个研究区而言,无优势类群,常见类群为不动杆菌属(Acinetobacter)、类诺卡氏菌属(Nocardioides),二者在各恢复年限样地中总占比为6.09%。其次为韩国生工菌属(Kribbella)、康奈斯氏杆菌属(Conexibacter)、短波单胞菌属(Brevundimonas)等稀有类群。不同恢复年限土壤细菌类群相对丰度存在差异。类诺卡氏菌属为恢复年限4、5、6、7 a沙棘林中相对丰度最高的菌群,不动杆菌属为恢复年限3 a沙棘林的优势类群且其相对丰度显著高于草地土壤。其他稀有类群随着人工林恢复年限的延长波动变化明显。Thermus、Deinococcus菌属相对丰度随恢复年限延长逐渐降低,CL500-29 marine group、Streptomyces菌属相对丰度则逐渐增加。
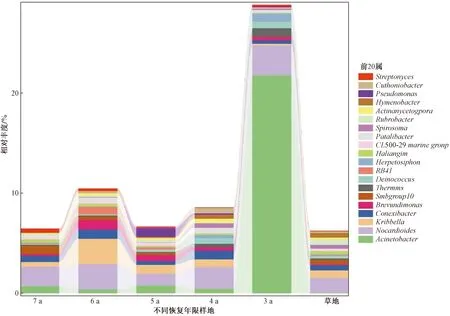
Acinetobacter为不动杆菌属; Nocardioides为类诺卡氏菌属; Kribbella为韩国生工菌属; Conexibacter为康奈斯氏杆菌属; Brevundimonas为短波单胞菌属; Therrmus为栖热菌属; Deinococcus为异常球菌属; Herpetosiphon为滑柱菌属; Patulibacter为散生杆菌; Spirosoma为螺状菌属; Rubrobacter为红色杆菌属; Actinomycetospora为放线胞菌属; Hymnenobacrer为薄层菌属; Psendomonas为假单胞菌属; Streptomyces为链霉菌属
2.2.2 土壤细菌群落差异性分析
为了进一步了解不同生境群落物种组成的组间差异性及相似性,因此进行了β-多样性分析。基于Bray-Curtis距离算法,采用 PCoA 分析不同恢复年限沙棘林土壤细菌群落组间差异(图4)。由图4可知,PC1解释了样品差异的27.2%,PC2解释了样品差异的23.9%,二者共同解释了物种分布差异的51.1%。PC1可明显将恢复年限3、4 a的细菌群落与5、6 a分开,PC2可明显将恢复年限3、4、7 a的细菌群落与5、6 a、草地分开。整体而言,恢复年限3、4 a的细菌群落结构较为相似,而5、6、7 a及草地的细菌群落较为相似,其中草地细菌群落与恢复6a的沙棘林最为相似。可见,恢复年限较短的细菌群落与较长年限的细菌群落差异性较大,恢复年限较长的沙棘林与撂荒草地细菌群落结构相似性较大。

图4 不同恢复年限沙棘林土壤细菌群落PCoA分析Fig.4 PCoA analysis of soil bacterial communities in seabuckthorn forest with different restoration years
2.3 细菌群落与土壤环境因子的相关性
为探讨土壤环境因子对土壤细菌群落组成的影响,对土壤细菌属水平群落与土壤理化性质进行冗余分析(RDA),如图5所示。第一排序轴解释了细菌群落相对丰度变化的39.89%,第二轴解释了细菌群落变化的32.72%,二者共同解释了细菌群落变化的72.61%。不动杆菌属(Acinetobacter)、类诺卡氏菌属(Nocardioides)与土壤pH和土壤含水量呈显著正相关关系,与土壤有机质呈显著负相关关系;链霉菌属(Streptomyces)、Subgroup10菌属、RB41菌属、Haliangium菌属与土壤碱解氮和速效钾含量呈显著正相关;康奈斯氏杆菌属(conexibacter)、短波单胞菌属(Brevundimonas)、韩国生工菌属(Kribbella)、红色杆菌属(Rubrobacter)与土壤有机质呈显著正相关,与土壤pH、含水量和速效钾显著负相关。土壤pH和速效钾是影响研究区土壤细菌群落结构的主要影响因子。

图5 主要细菌类群与土壤环境因子的RDA分析Fig.5 RDA analysis of major bacterial groups and soil environmental factors
2.4 土壤细菌PICRUSt功能预测
为了解不同恢复年限人工林土壤细菌代谢功能,采用PICRUSt软件进行菌群预测(图6)。基于KEGG 数据库,在一级代谢通路层对比共得到6类生物功能:代谢(metabolism)、基因信息处理(genetic information processing)、细胞进程(cellular processes)、环境信息处理(environmental information processing)、人类疾病(human diseases)和生物体系统(organismal systems)。其中,代谢、基因信息处理和细胞进程为主要功能,占比分别为82.00%~83.56%,10.06%~11.59%和3.26%~3.88%。与草地相比,沙棘林土壤细菌代谢、环境信息处理、人类疾病、生物体系统4种功能基因丰度较高;且随恢复年限的增加,基因信息处理、细胞进程、人类疾病、生物体系统功能基因呈波动增长趋势。

图6 不同恢复年限土壤细菌基因功能预测(一级功能)Fig.6 Prediction of soil bacterial gene function at different restoration years (level 1 function)
二级功能预测表明,沙棘林土壤和草地土壤均存在碳水化合物代谢、氨基酸代谢、脂质代谢、能量代谢、信号转导、膜运输、细胞生长和死亡、免疫系统等35个子功能(图7)。其中,碳水化合物代谢(13.01%~13.87%)、氨基酸代谢(13.13%~13.46%)、辅因子和维生素代谢(10.42%~11.72%)为主要功能。与草地相比,沙棘林土壤膜运输、信号传导、癌症、传染病功能基因丰度较高,转录、氨基酸代谢、其他氨基酸的代谢和萜类和聚酮的代谢基因丰度较低。其中,折叠、分拣和降解、复制和修复、转录、神经变性疾病、萜类和聚酮的代谢、核苷酸代谢功能基因丰度随恢复年限增长而增加,其他氨基酸代谢、外源生物降解和代谢、内分泌系统、环境适应功能基因丰度随恢复年限增长而减小。
3 讨论
3.1 恢复年限对沙棘林土壤理化性质的影响
沙棘属于复合型根系,低耗水、萌蘖能力极强的固氮植物[29-31],连续种植使区域土壤理化性质发生改变。沙棘可短期内迅速提高土壤有机质含量及土壤养分[32]。研究发现,随恢复年限的增加,土壤有机质显著增加(P<0.05),这与张伟华等[33]、常丽等[34]的研究结果相似,这可能是因为沙棘属于落叶灌木且根系发达,林下凋落物及植物根系残体随恢复年限积累,经腐殖化作用使土壤有机质含量不断增加[35]。土壤碱解氮含量随恢复年限增加呈先增加后减小的趋势,这可能是因为在复垦前中期(3~6 a),沙棘根系被根瘤菌侵染形成共生结构,随恢复年限的延长瘤块不断趋于成熟,因而固定到土壤中的氮素也随之增加[36],而后氮素含量又减小可能是因为植物生长发育的需求量较大。与土壤有机质和碱解氮变化规律不同,土壤水分含量随恢复年限的延长逐渐减少,这是因为随着林龄的增大,植被耗水量急剧增加,因此土壤含水量不断下降[37]。在干旱半干旱地区植被生长所需水分主要来自于大气降水,土壤对水分的调控能力很差,因此,恢复年限为6 a沙棘林应采取平茬[38]等措施,从而减少林分对土壤水分的不利影响,促进沙棘林的更新和健康发展。
3.2 恢复年限对沙棘林土壤细菌群落结构的影响
森林植被类型、恢复年限是影响土壤细菌群落的主要因素[39-40]。不同植被地表凋落物数量和质量、地下根系的结构、分泌物的不同,造成土壤微生境存在异质性,使土壤细菌组成及数量等发生改变[41-42]。研究发现,在门分类水平上,沙棘人工林和撂荒草地土壤细菌物种组成相似,均以放线菌门、变形菌门为优势菌门,拟杆菌门、酸杆菌门、绿弯菌门、浮霉菌门、疣微菌门、蓝藻门为常见菌门,但各菌群丰度存在差异。变形菌门以土壤中甲烷、氨气等满足自身代谢,喜好富营养土壤[43]。沙棘土壤变形菌门丰度显著高于草地,说明沙棘林的建植有利于土壤有机质及土壤养分的提高。随着恢复年限的延长,变形菌门丰度呈现先减少后增加的趋势,且在恢复期3 a时丰度最大,这可能是因为土壤酸碱度对细菌的影响较大,而变形菌门更适宜生存于碱性土壤环境[44],3 a时土壤pH最大,而后先减小后增加。放线菌门生态幅较宽,在不同生态系统中多以优势菌群出现。在本研究中,沙棘林放线菌门相对丰度随着恢复年限的延长呈现先增加后减少的趋势,且在恢复年限第5年时显著高于草地。放线菌在木质素、纤维素、动物残体等的降解和转化方面具有重要作用[45]。恢复年限第5年时草地土壤C、N含量远低于沙棘林,这可能是其放线菌门相对丰度低于沙棘林的原因。6 a开始沙棘林土壤中放线菌门含量逐渐下降,可能是因为沙棘林开始退化,种间竞争激烈而开启自毒功能,土壤中可利用资源减少从而对细菌产生不利影响。 在属分类水平上,沙棘林土壤不动杆菌属、类诺卡氏菌属相对丰度整体高于草地,这可能是植被类型不同的原因。不动杆菌属、类诺卡氏菌属等可通过产生抗生素对植物病原菌产生拮抗作用[46-48]。而沙棘具有庞大的根系,随着恢复年限的延长,其根系分泌的有机酸等物质不断积累[49],在化感作用的影响下,某些物质可能有利于增加对病原菌有拮抗作用的有益菌群的活性,与有益菌群产生协同拮抗效应[50]。长期种植沙棘使土壤中的栖热菌属(Thermus)、奇球菌属(Deinococcus)相对丰度逐渐减小。 栖热菌属和奇球菌属间具有非常相近的亲缘关系,二者均对干燥、紫外线、强氧化剂等各种恶劣环境有极强的抵抗力[51],随着恢复年限的延长,土壤环境改善,因此二者含量逐渐减少。另外,二者均与有机质、碱解氮、速效钾呈显著负相关,再次说明了其更适宜生活在低营养的贫瘠土壤中。因此,在一定程度上,可以把栖热菌属和奇球菌属作为指示物种,通过其在土壤中含量的变化来表征生境的土壤健康状况。
3.3 恢复年限对沙棘林土壤细菌基因功能的影响
植被通过影响地上与地下部分影响细菌群落结构,进而对细菌功能产生影响[52]。对人工林和草地土壤细菌基因进行PICRUSt功能预测显示,研究区土壤细菌主要有代谢、基因信息处理、细胞进程、环境信息处理、人类疾病和生物体系统5个一级功能,以及碳水化合物、氨基酸代谢、脂质代谢、能量代谢和信号传导等35个二级功能,细菌生态功能较丰富。其中,细菌代谢功能尤为突出,这与王晓菲等[53]的研究结果一致。其二级功能中碳水化合物代谢的通过糖酵解等过程产生的能量与代谢产物有利于植物和细菌的生长和发育[54]。另外,碳水化合物代谢与植物体内所需的氮、磷元素紧密相关,对促进植物氮磷循环有积极作用[55]。氨基酸代谢有利于细菌个体对氨基酸的吸收利用,并可参与多种重要的生理过程,有益于细菌的生存和增殖[56]。部分细菌类群如不动杆菌属、类诺卡氏菌属等自身代谢可产生抗生素、生长激素、抗菌蛋白等对植物病原菌产生拮抗作用,促进植物生长。研究区沙棘林各恢复年限间土壤细菌代谢功能差异不明显,但均高于草地,说明沙棘人工林的建植有利于细菌各代谢功能的提升,从而对植被自身生长起到积极作用。随着恢复年限的延长,折叠、分拣和降解、复制和修复、转录等细菌功能基因丰度逐渐增加,而氨基酸代谢、外源生物降解和代谢、内分泌系统、环境适应功能基因丰度逐渐减小,这可能与植被恢复过程中气候、土壤、生物因素变化有关。
4 结论
(1)与草地相比,沙棘人工林的建植使土壤有机质、碱解氮、速效钾、含水量呈增加趋势,土壤pH和全磷变化不明显。
(2)研究区共检测到细菌29门、91纲、193目、305科和438属,门分类水平上优势类群为放线菌门、变形菌门;属分类水平上无优势类群,常见类群为不动杆菌属、类诺卡氏菌属。
(3)沙棘林土壤细菌不动杆菌属、类诺卡氏菌属相对丰度整体高于草地;随着恢复年限的延长,栖热菌属和奇球菌属相对丰度随恢复年限延长逐渐降低,CL500-29marinegroup、链霉菌属相对丰度则逐渐增加。土壤pH和速效钾是影响研究区沙棘人工林土壤细菌群落结构的主要影响因子。
(4)研究区沙棘林和草地土壤细菌主要包含5个一级功能和35个子功能,细菌生态功能丰富。最主要的细菌代谢功能在沙棘林土壤中较草地更为活跃,说明沙棘林土壤较草地细菌代谢潜力更强,而细菌代谢功能反过来也会通过分泌各种代谢产物来促进植物更好的生长和发育。
