丛枝菌根真菌与褪黑素对多年生黑麦草耐热性的影响
2023-12-08卫宏健贺文员王越唐明陈辉
卫宏健,贺文员,王越,唐明,陈辉
(华南农业大学林学与风景园林学院, 岭南现代农业科学与技术广东省实验室, 广东 广州 510642)
褪黑素(N-乙酰-5-甲氧基色胺,melatonin, MT,化学式C13H16N2O2)是广泛存在于动植物体内的吲哚类化合物,具有很强的抗氧化作用[7]。植物中褪黑素的合成主要涉及色氨酸脱羧酶(tryptophan decarboxylase, TDC)、色胺5 羟化酶(tryptamine 5-hydroxylase, T5H)、血清素N-乙酰转移酶(serotonin N-acetyltransferase, SNAT)、咖啡酸O-甲基转移酶(caffeic acid O-methyltransferase, COMT)和乙酰血清素甲基转移酶(acetylserotonin methyltransferase, ASMT)[8-9]。褪黑素作用于多种植物细胞代谢和生物过程,包括生根、光合作用和胁迫耐受性[10-11]。特别是褪黑素(无论是外源施用或基因表达生物合成的内源褪黑素)作为一种有效的抗氧化剂、膜稳定剂和生物刺激剂,可改变氧化还原基因的表达或者渗透物质的水平,从而显著提高植物对生物与非生物胁迫的耐受性[12]。外源褪黑素通过提高高羊茅(Festuca arundinacea)幼苗体内抗氧化酶活性水平和叶片的光合效率,同时降低膜渗透性、丙二醛(malonaldehyde, MDA)和过氧化氢(hydrogen peroxide, H2O2)含量,以提高高羊茅幼苗的耐热性[13]。高温胁迫下植物调节褪黑素合成的相关基因(如BjTDC1和BjSNAT1)的表达,以增强芥菜(Brassica juncea)的高温耐受性[14]。因此,富含褪黑素的植物或外源施用褪黑素在改善植物生长和抗胁迫能力方面具有很高的潜力。
已知大多数陆生植物的根系能与丛枝菌根真菌(arbuscular mycorrhizal fungi, AMF)形成共生关系[15]。前人研究发现,丛枝菌根(arbuscular mycorrhizal, AM)共生能够提高宿主植物对干旱、盐、高温和低温等非生物胁迫的抗性[16-18]。高温胁迫下,接种AMF 通过延缓叶片的可溶性糖、可溶性蛋白质、游离脯氨酸和叶绿素质量分数的下降以提高杜鹃花(Rhododendron simsii)的耐热性[19]。接种AM 真菌(Septoglomus deserticola和Septoglomus constrictum)可以通过降低番茄(Solanum lycopersicum)脂质过氧化水平和H2O2积累量,提高活性氧清除剂的活性,进而缓解番茄的高温损伤[20]。Zhang 等[21]发现AM 真菌异形根孢囊霉(Rhizophagus irregularis)通过提高蒺藜苜蓿(Medicago truncatula)内源褪黑素的含量,提高了对重金属铅(Pb)的耐受性,证明了共生微生物在植物中调节和合成褪黑素的潜力。目前关于菌根化植物在热胁迫下调节褪黑素产生的机制,以及AM 真菌与褪黑素联合应用对宿主植物的发育和耐热性方面的影响尚不清楚。因此,本研究以多年生黑麦草为试验材料,通过接种AM真菌异形根孢囊霉和浇灌外源褪黑素,探索AMF 和外源褪黑素对高温胁迫下多年生黑麦草的生长、内源褪黑素含量及其合成基因的表达水平,抗氧化酶活性和小分子渗透物质含量的影响,为解析褪黑素对菌根化植物的非生物胁迫抗逆性机制提供理论基础。
1 材料与方法
1.1 试验材料与基质
本研究所用的多年生黑麦草种子(品种:首相Ⅲ)购买于天津百绿国际草业有限公司。多年生黑麦草种子在75%的乙醇溶液中浸泡5 min 进行表面消毒,然后用无菌蒸馏水漂洗4 次。此外,将冲洗过的种子种植在含有基质(沙子∶泥炭=1∶3,v/v)的塑料盆中(高10 cm,直径10 cm),播种密度为20 g·m-2。基质中有机质含量5.68 g·kg-1、速效磷含量22.01 mg·kg-1、速效氮含量34.53 mg·kg-1和速效钾含量71.23 mg·kg-1。在种植多年生黑麦草种子之前,用75%的乙醇溶液对塑料盆进行消毒,并在播种前连续3 d 在高压灭菌锅中(121 ℃,0.11 MPa)对基质进行消毒1 h。在最适温度条件[25 ℃/20 ℃(昼/夜)]、60%的相对湿度、14 h/10 h(光/暗)的光周期以及650 mmol·m-2·s-1的光照强度下,将幼苗养在生长室中。为保持土壤湿润,初始阶段花盆在早上和晚上都进行了灌溉。出苗后每天给植物浇水一次,每7 d 对植物进行修剪,以保持8 cm 的冠层高度,同时每周使用半浓度的Hoagland 营养液浇灌一次,以保证足够的营养供应。外源褪黑素(AR 级),购于上海麦克林生化科技有限公司。研究使用的丛枝菌根真菌为异形根孢囊霉(BGC BJ09,Ri)来自华南农业大学林学与风景园林学院,利用玉米(Zea mays)作为宿主植物进行扩繁后,用蔗糖梯度离心法获得的孢子作为接种物[22]。将1 mL 含有约400 个孢子的水溶液作为孢子剂接种到14 d 苗龄的多年生黑麦草幼苗根系附近,非菌根(non-mycorrhizal,NM)处理为接种相同体积的灭菌孢子剂(在121 ℃的高压灭菌锅中灭菌15 min)。
1.2 试验设计
在接种Ri 45 d 后于2022 年10 月在华南农业大学林学与风景园林学院进行盆栽试验。选取48 盆多年生黑麦草幼苗,均分为最适温度条件(CK)和高温条件(high temperature,HT)两组,每组包含4 个处理,包括:1)既不接种AM 真菌也不浇灌外源褪黑素溶液(NM-NT);2)仅根部接种AM 真菌(AM-NT);3)仅用褪黑素溶液灌根(NM-MT);4)褪黑素和AM 真菌联合处理(AM-MT)。每个处理6 个重复(6 个盆栽),最适温度(CK)为25 ℃/20 ℃(日/夜),高温处理(HT)为38 ℃/33 ℃(日/夜),生长室其他条件不变:60%的相对湿度、14 h/10 h(光/暗)的光周期以及650 mmol·m-2·s-1的光照强度,高温处理共进行28 d。参考Zhang 等[23]的试验设计,在热胁迫处理前3 d,每天用5 mL 20 mmol·L-1褪黑素(MT)灌溉幼苗,随后在余下的28 d 内每隔7 d 浇灌一次,对不施用褪黑素(NT)的幼苗浇灌等体积的蒸馏水。高温处理28 d 后收样,将地上地下部分分开,洗净后测量鲜重后放进80 ℃烘箱内烘干至恒重,称量并记录其干重。其中每个处理选取4 盆植株根系来评估AM 定殖的效果,样品的其余部分用液氮研磨成粉末,并保存在-80 ℃冰箱后进行下一步分析。
1.3 菌根定殖率的测定
每个处理随机选取4 盆多年生黑麦草的根系,按照Koske 等[24]的方法对根系进行染色,利用放大十字交叉法[25]计算菌根定殖率。菌根定殖率(%)=侵染点数量/检测的总交叉点数量×100。
1.4 抗氧化酶活性测定
采用氮蓝四唑法测定超氧化物歧化酶(superoxide dismutase,SOD)活性[26]。采用愈创木酚法测定过氧化物酶(peroxidase, POD)活性[27]。采用紫外吸收法测定过氧化氢酶(catalase,CAT)活性[28]。采用Nakano 等[29]的方法测定抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性。参考Chen 等[30]的方法测定吡咯啉-5-羧酸还原酶(pyrroline-5-carboxylate reductase,P5CR)活性。采用深圳子科生物科技有限公司的试剂盒,测定苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)和多酚氧化酶(polyphenol oxidase,PPO)活性。
1.5 膜脂过氧化指标与渗透调节物质测定
参照高俊凤[31]的方法测定根系MDA 含量和相对电导率(electrolytic leakage,EL)。采用苏州科铭技术有限公司生产的试剂盒测定渗透调节物质含量,按照试剂盒说明进行测定并计算脯氨酸、可溶性糖、类黄酮、总酚和甜菜碱含量。
1.6 内源褪黑素含量测定
取0.1 g 根样,液氮速冻后低温研磨至粉末,加入900 μL PBS 缓冲液(pH=7.4)充分混合,12000 r·min-1离心15 min 后取上清液为待测样品。使用深圳子科生物科技有限公司褪黑素酶联免疫分析试剂盒(酶联生物)测定褪黑素含量。
(5)成效慢.白猿通背拳需要日积月累的练习才能达到一定的效果,但反观现在发展的比较出色的跆拳道、空手道等,其基本内容略微简单,训练体系较为明晰,学生学上数月,便可看出成效.但白猿通背拳的练习则要以年来计算,并且对身体协调性与力量有着极高的要求,若没有足够的时间练习,则很难收获想要的成绩.
1.7 基因表达分析
总RNA 采用Trizol 试剂盒参考说明书进行提取,经Nanodrop 2000 检测浓度和纯度以及用1%琼脂糖凝胶电泳检测完整性后,根据Fermentas cDNA synthesis 试剂盒说明书进行反转录合成cDNA。参考TaKara 公司的SYBR System 操作手册,采用实时荧光定量PCR 仪(BioRad,CFX96,美国)进行PCR 扩增。每个处理设置 6 个生物学重复,每个生物学重复设置3 个技术重复。基因特异性引物序列参考表 1 进行合成,以LpeIF4A基因为内参基因[32]。根据得到的Ct 值,利用2-ΔΔCt法,分别计算目标基因在不同处理下的相对表达量。

表1 试验所用引物Table 1 Primers used in the study
1.8 数据分析
采用Microsoft Excel 2019 对试验数据进行图表分析,用 SPSS 22.0 软件进行方差分析,用 Duncan’s 法在P<0.05 水平下进行多重比较。采用Origin 2022 进行制图以及主成分分析(principal component analysis,PCA)。
2 结果与分析
2.1 外源褪黑素处理对高温胁迫下AMF 定殖率的影响
无论是否添加了外源褪黑素,热胁迫均显著降低了Ri 在多年生黑麦草根系的定殖率(P<0.05)。在最适温度和热胁迫条件下,褪黑素处理均显著提高了Ri 的定殖率(P<0.05),其中最适温度下褪黑素处理的菌根化多年生黑麦草表现出更高的定殖率(87.61%)(图1)。

图1 不同处理下丛枝菌根真菌在多年生黑麦草根系的定殖率Fig. 1 Colonization rate of perennial ryegrass roots by arbuscular mycorrhizal fungi under different treatments
2.2 外源褪黑素处理和AM 共生条件下多年生黑麦草内源褪黑素含量及其合成基因转录水平对高温胁迫的响应
最适温度条件下,相比于NM-NT 和AM-NT 处理,NM-MT 和AM-MT 处理的根系内源MT 含量分别显著增加了1.12 和1.03 倍(P<0.05)。在高温条件下,外源褪黑素显著提高了AM 和NM 多年生黑麦草的根系内源MT 含量(P<0.05)(图2A)。

图2 不同处理下的多年生黑麦草内源褪黑素含量及其合成基因转录水平Fig.2 Endogenous melatonin content and transcript levels of its synthetic genes in perennial ryegrass under different treatments
高温胁迫下,相比于AM-NT 处理,AM-MT 处理的 MT 合 成 基 因LpTDC1、LpTDC2、LpT5H、LpSNAT、LpASMT1和LpASMT3的转录水平显著上升了43.84%、50.55%、21.25%、32.63%、40.25% 和44.83%(P<0.05),而LpCOMT1的相对表达量则无显著变化(P>0.05)(图2B~H)。与NM-MT 处理相比,AM-MT 处理的根系LpTDC1、LpT5H、LpSNAT、LpCOMT1、LpASMT1和LpASMT3基因相对表达量显著增加(P<0.05)(图2B~H)。此外,在最适温度条件下,AM-NT 和AM-MT 处理的根系LpTDC1、LpT5H和LpSNAT基因的转录水平与NM-NT 和NMMT 处理差异不显著(P>0.05)(图2B,D,E)。AMMT 和NM-MT 处理的根系LpTDC2、LpCOMT1、LpASMT1和LpASMT3基因转录水平显著高于(P<0.05)AM-NT 和NM-NT 处理。
2.3 外源褪黑素处理和AM 共生条件下多年生黑麦草生长参数对高温胁迫的响应
高温胁迫抑制了多年生黑麦草的生长,而AM-NT、NM-MT 和AM-MT 处理均减轻了热应激引起的生长抑制(图3A,B)。在高温条件下,AM-MT 处理的草坪质量、株高、地上和地下部分的鲜重和干重均显著高于AMNT 和NM-MT 处理(P<0.05)(图3C 和表2)。此外,在最适温度条件下,AM-NT 处理的草坪质量、株高、地上和地下部分的鲜重和干重与NM-MT 处理差异不显著(P>0.05)。

图3 不同处理对高温胁迫下多年生黑麦草生长和草坪质量的影响Fig.3 Effect of different treatments on growth and turf quality of perennial ryegrass under high temperature stress
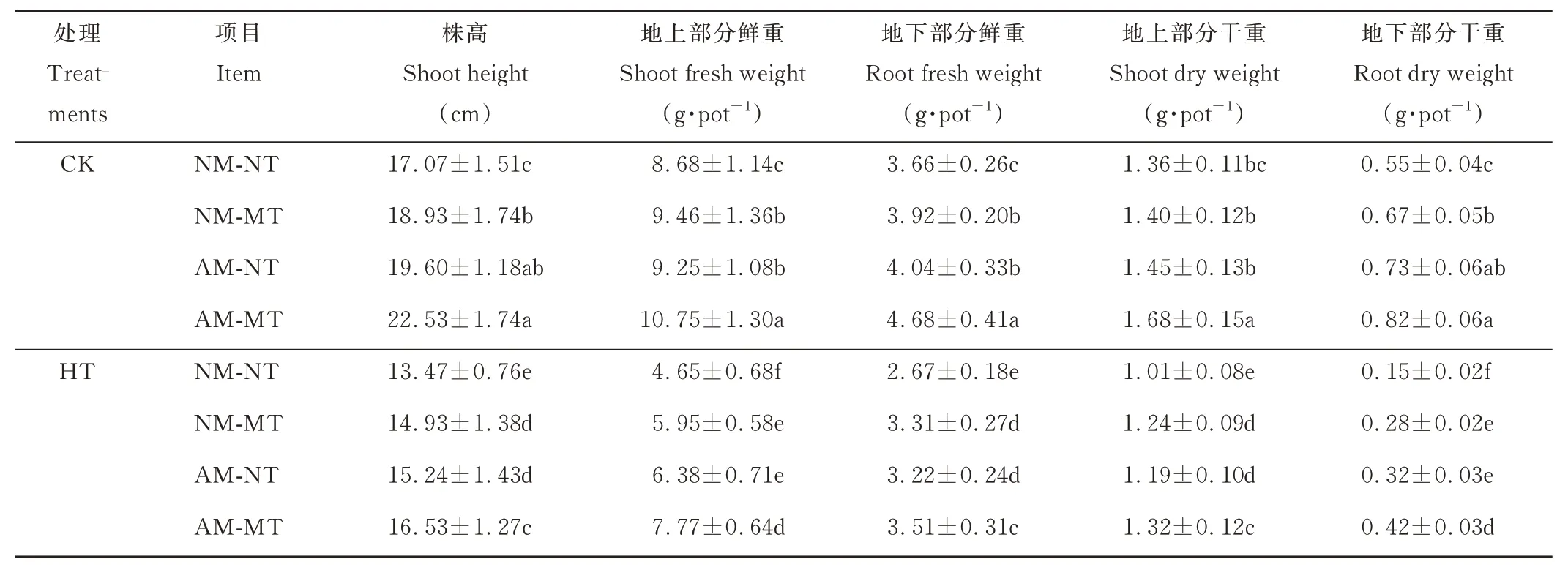
表2 不同处理下的多年生黑麦草生长参数Table 2 Growth parameters of perennial ryegrass under different treatments
2.4 外源褪黑素处理和AM 共生条件下多年生黑麦草膜脂过氧化和抗氧化酶活性对高温胁迫的响应
高温胁迫下,与NM-NT 相比,AM-NT、NM-MT 和AM-MT 处理均显著降低了根系MDA 含量和EL(P<0.05),且AM-MT 的根系MDA 含量和EL 显著低于其他处理(P<0.05)。此外,在最适温度条件下,NM-NT,NM-MT,AM-NT 和AM-MT 处理之间的根系MDA 含量和EL 无显著差异(P>0.05)(图4A, B)。

图4 不同处理下的多年生黑麦草膜脂过氧化指标和抗氧化酶活性Fig.4 Indicators of membrane lipid peroxidation and antioxidant enzyme activity in perennial ryegrass under different treatments
最适温度条件下,NM-NT,NM-MT,AM-NT 和AM-MT 处理之间的根系SOD、POD 和APX 活性无显著差异(P>0.05)(图4C,D 和F),而与NM-NT 和NM-MT 处理相比,AM-NT 和AM-MT 处理显著提高了根系CAT活性(P<0.05)(图4E)。热胁迫条件下,与NM-NT 相比,NM-MT,AM-NT 和AM-MT 处理均显著提高了根系SOD、POD、CAT 和APX 的活性(P<0.05),其中AM-MT 处理下这4 种酶的活性最高。
2.5 外源褪黑素处理和AM 共生条件下多年生黑麦草PAL 和PPO 活性及其编码基因转录水平对高温胁迫的响应
最适温度条件下,NM-NT,NM-MT,AM-NT 和AM-MT 处理之间的PAL 和PPO 活性,以及LpPAL和LpPPO的转录水平无显著差异(P>0.05)。高温胁迫下,与NM-NT 相比,AM-NT、NM-MT 和AM-MT 处理均显著提高了根系PAL 活性和LpPAL基因的转录水平(P<0.05),且AM-MT 处理显著高于其他处理(P<0.05)(图5A,B)。与NM-NT 相比,高温胁迫下AM-NT 和AM-MT 处理均显著降低了根系PPO 活性和LpPPO转录水平(P<0.05),其中AM-MT 处理显著低于其他处理(P<0.05)(图5C,D)。
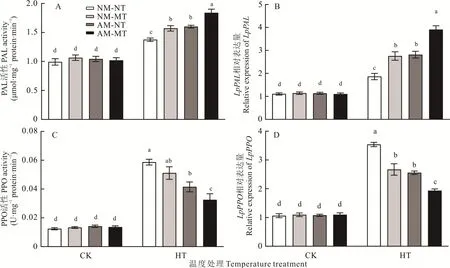
图5 不同处理下的多年生黑麦草PAL 和PPO 活性及其编码基因转录水平Fig.5 PAL and PPO activities and the transcription levels of their encoding genes in perennial ryegrass under different treatments
2.6 外源褪黑素处理和AM 共生条件下多年生黑麦草渗透调节物质对高温胁迫的响应
在最适温度条件下,NM-NT,NM-MT,AM-NT 和AM-MT 处理之间的根系脯氨酸含量(图6A)、P5CR 活性(图6B)和LpP5CS转录水平(图6C)无显著差异(P>0.05)。高温胁迫下,与NM-NT 相比,NM-MT 和AM-MT处理均显著提高了(P<0.05)根系的脯氨酸含量、P5CR 活性和LpP5CS转录水平(图6A~C),但NM-MT 与AM-MT 处理之间均无显著差异(P>0.05),且NM-MT 处理的根系脯氨酸含量显著低于AM-NT 处理(P<0.05)。高温胁迫下,NM-MT 处理的根系类黄酮含量显著高于AM-NT 和AM-MT 处理(P<0.05)(图7B),而NM-MT 和AM-NT 处理下的根系可溶性糖、总酚和甜菜碱含量则无显著差异(P>0.05),AM-MT 处理的根系可溶性糖、总酚和甜菜碱含量均显著高于其他处理(P<0.05)(图7A,C 和E)。

图6 不同处理下的多年生黑麦草脯氨酸含量、P5CR 活性和LpP5CS 转录水平Fig.6 Proline content, P5CR activity and LpP5CS transcript levels in perennial ryegrass under different treatments
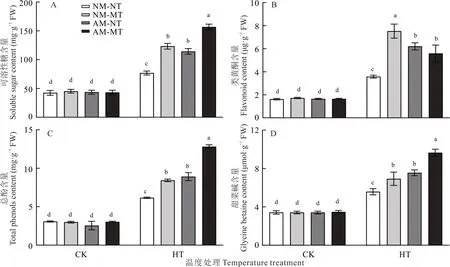
图7 不同处理下的多年生黑麦草渗透调节物质含量Fig.7 Content of osmoregulatory substances in perennial ryegrass under different treatments
2.7 主成分分析
为评价高温胁迫下不同处理对多年生黑麦草根系抗氧化系统与渗透调节物质的影响,进行了主成分分析,使用多年生黑麦草根系的抗氧化相关参数(MDA、EL、SOD、POD、CAT 和APX)和渗透调节物质相关参数(脯氨酸、总酚、类黄酮、甜菜碱和可溶性糖含量,以及PAL 和PPO 活性)的数据进行分析。每个点代表本试验中的一种处理类型,其中两个主成分中PC1 占比77.2%,PC2占比16.4%,PC1 显示CK(最适温度)处理与HT(高温)处理明显分开,AM-MT-HT 处理与其他处理的差异最大(图8)。

图8 不同处理下的多年生黑麦草抗氧化与渗透调节物质相关参数主成分分析Fig. 8 Principal component analysis of parameters related to antioxidant and osmoregulatory substances in perennial ryegrass under different treatments
3 讨论
在非胁迫条件下,与拟南芥(Arabidopsis thaliana)等植物类似,多年生黑麦草含有较低和恒定量的褪黑素[33],胁迫条件下褪黑素能够通过植物信号转导帮助植物应对氧化损伤,发挥其内源性抗氧化剂的作用[34]。本研究中高温胁迫刺激了根系中褪黑素的积累,表明高温胁迫下多年生黑麦草可能通过增加褪黑素的积累提高其抗氧化能力。AM 接种促进了高温胁迫下褪黑素的合成,这表明AM 植物在高温胁迫下可以产生丰富的褪黑素,以应对热应激导致的氧化损伤。Liu 等[35]观察到胁迫条件下真菌Trichoderma asperellum中褪黑素的积累,说明微生物在非生物胁迫条件下参与褪黑素的积累。因此,AM 真菌产生的褪黑素可能有助于热胁迫下多年生黑麦草根部褪黑素的积累。外源施用褪黑素或过表达褪黑素生物合成基因可提高植物内源褪黑素含量,增强植物抗逆性[36]。植物体内褪黑素合成首先通过TDC 和T5H 活性将色氨酸转化为血清素。在转基因水稻(Oryza sativa)种子中过表达OsTDC3导致褪黑素水平比野生型(wild type, WT)增加31 倍,进而延迟转基因水稻中暗诱导的叶片衰老[37]。SANT 催化5-羟色胺转化为N-乙酰血清素,人类SNAT基因在转基因水稻中的异位表达除了表现出强耐寒性外,还表现出更高的内源性褪黑素水平[38]。此外,COMT 将N-乙酰5-羟色胺甲基化为褪黑素,在转基因水稻中过表达OsCOMT也显著提高内源褪黑素含量,抑制OsCOMT的表达导致褪黑素水平降低[39]。ASMT 是褪黑素合成中的末端酶,在褪黑素合成过程中起限速作用[40]。Fu 等[41]发现在干旱和低温胁迫下多年生黑麦草通过上调LpASMT1和LpASMT3基因的转录水平,以增加内源褪黑素的积累,进而增强耐旱性和耐寒性。本研究发现外源MT 处理导致高温胁迫下NM 和AM 多年生黑麦草内源褪黑素含量显著增加,与较高的褪黑素合成基因LpTDC1、LpT5H、LpSNAT、LpCOMT1、LpASMT1和LpASMT3的转录水平相关。因此,褪黑素生物合成基因转录水平的提高和内源褪黑素含量上升可提高多年生黑麦草的耐热性。
热胁迫显著降低了Ri 在多年生黑麦草根系的定殖率。Wei 等[18]认为非生物胁迫下AMF 在植物根中定殖率的下降与植物的光合作用下降有关,因为光合作用下降减少了宿主植物对菌根真菌的碳分配。在对热胁迫下生菜(Lactuca sativa)[42]和薰衣草(Lavandula angustifolia)[43]的菌根真菌定殖研究中也发现了类似的结果。褪黑素处理和AM 接种提高了高温胁迫下多年生黑麦草的株高,地上和地下部分鲜重和干重,这是因为AM 共生和褪黑素都能保护宿主植物的光合系统,提高在高温胁迫下的光合速率,从而增加生物量,改善宿主植物对AMF 的碳分配并提高定殖率。
一般来说,在合适的生长条件下,植物细胞中的ROS 水平处于动态平衡状态,而非生物胁迫可以诱发过多的ROS 生成和膜脂过氧化,导致植物代谢功能失调[44]。细胞内MDA 的产生是过多ROS 引起细胞膜脂过氧化的结果,EL(反映膜的完整性)和MDA 水平是评估植物氧化应激水平的重要指标[45]。AM 接种和褪黑素处理均降低了EL 和MDA 水平,以保护多年生黑麦草免受氧化应激。植物活性氧清除系统大多包含抗氧化物质和抗氧化酶,包括SOD、APX 和POD,SOD 催化O2-歧化为O2和H2O2,而CAT 和POD 则分别在叶绿体和细胞质将H2O2降解为无毒害作用的水分子和氧气,同时APX 参与抗坏血酸-谷胱甘肽(AsA-GSH)循环,催化抗坏血酸和H2O2反应,从而协助清除植物体内的H2O2[18,46]。因此,褪黑素和AM 共生对高温耐受性的协同作用与减少氧化损伤和提高了抗氧化酶活性相关。
脯氨酸能调节氧化还原电位,保护大分子不被羟基自由基变性和清除,作为一种重要的渗透调节物质,可提高细胞的渗透调节能力,参与细胞代谢,维持膨压,保护膜结构的完整性,以维持正常的代谢活动,从而降低非生物胁迫对植物细胞的伤害[47]。1-吡咯啉-5-羧酸合成酶(1-pyrrolin-5-carboxylate synthetase, P5CS)和P5CR 是植物脯氨酸合成途径的关键酶[48]。Antoniou 等[49]发现褪黑素处理可以增加脯氨酸的积累以缓解干旱对紫花苜蓿(Medicago sativa)造成的损伤。也有研究发现褪黑素处理通过上调MtP5CS基因的转录水平和增加P5CR 活性来增加脯氨酸含量,以提高菌根化蒺藜苜蓿的重金属耐受性[21]。本研究结果与之相似,Ri 接种与褪黑素处理以及两者的联合,通过提高P5CR 活性和LpP5CS基因的转录水平,增加热胁迫条件下多年生黑麦草根部脯氨酸含量,提高渗透调节能力,进而提高植物的抗逆性。PAL 是苯丙烷代谢途径的一种关键酶,植物体内苯丙烷途径的代谢产物包括多酚类、黄酮类和花青素,高温会诱导植物体内这些保护性小分子物质的合成和积累,以缓解热胁迫损伤[50]。Liang 等[51]认为使用褪黑素可以通过提高SOD、CAT 和POD 活性,提高抗氧化能力,并通过促进类黄酮生物合成缓解叶片衰老。褪黑素处理增强了猕猴桃(Actinidia chinensis)的PAL 活性,提高了耐寒性,表明了褪黑素在类黄酮合成中的潜在作用[52]。热胁迫条件下PPO 通过苯酚氧化产生剧毒的醌类物质,降低膜的完整性和抗氧化活性,导致膜脂过氧化[53]。氧化和聚合会对酚类物质的特性产生不利影响,妨碍其清除ROS 的能力。接种Ri 和应用外源褪黑素降低了PPO 活性和增强了PAL 活性,表明AM 接种和应用褪黑素增加了酚类物质的代谢,提高了多年生黑麦草的高温耐受性。在涉及AMF 接种的植物生物相互作用过程中,经常会出现PAL 活性的增加,以及酚类物质的积累。此外,上调的苯丙素途径可能参与了宿主与AMF 之间信号传递,这可能是植物面对非生物胁迫的一种防御策略[54]。
可溶性总糖是草坪草细胞碳水化合物代谢中的关键物质,对组织、器官的构建起重要作用,有利于增强草坪草的抗逆性,含量高低一般与植物抵抗逆境的能力呈正比[55]。甜菜碱是植物面对非生物胁迫时一种常见的渗透调节物质,参与植物细胞的渗透调节,亦可作为渗透保护物质提高生物大分子结构和功能的稳定性,防止蛋白质结构受到其他物质的破坏,稳定蛋白质的四级结构,尤其是在胁迫条件下,甜菜碱可以作为一种低分子量的伴侣分子起到对酶蛋白构象的稳定作用,并使酶蛋白处于激活且有功能的状态,增强酶的活力[56]。因此,可溶性总糖和甜菜碱含量的上升,可以有效激活多年生黑麦草的酶促防御系统,增强高温胁迫下多年生黑麦草的抗氧化酶活性,进而提高多年生黑麦草抵御高温胁迫的能力。AM 接种与外源褪黑素均可以通过增加酚类物质、脯氨酸和可溶性糖的含量,改善植物的渗透调节,提高多年生黑麦草的耐热性。然而,AM 接种和褪黑素应用对脯氨酸和类黄酮积累没有协同作用,这表明AM 真菌和褪黑素的应用对提高高温耐受性有不同的作用。黄酮类化合物和脯氨酸的合成都受到水杨酸的调控[57-58],褪黑素应用[59]和AM 接种[60]对水杨酸调节的相反影响可能解释了这一结果。而水杨酸代谢是否导致AM 共生和褪黑素之间的相互作用,以及褪黑素-植物激素相互作用对AM 植物的非生物胁迫抗性的影响,还有待进一步研究。
4 结论
总而言之,高温胁迫下褪黑素处理促进了多年生黑麦草根部的AM 共生,且AM 接种与外源褪黑素处理可以通过上调多年生黑麦草根系中褪黑素合成基因的转录水平,提高内源褪黑素的含量。AM 接种和褪黑素处理对多年生黑麦草的生长和高温耐受性有协同作用,这种协同作用可能与减轻氧化损伤,调控抗氧化酶活性,以及渗透调节物质含量有关。所以,AM 接种与褪黑素处理联合应用是帮助宿主植物应对高温环境的潜在有效策略。
