贺兰山不同海拔植被生物量与土壤分形特征和土壤水分的相关关系
2023-12-08黄业芸邱开阳朱亚超谢应忠刘王锁杨壹王思瑶崔璐瑶鲍平安
黄业芸,邱开阳*,朱亚超,谢应忠,刘王锁,杨壹,王思瑶,崔璐瑶,鲍平安
(1. 宁夏大学林业与草业学院,宁夏 银川 750021;2. 宁夏大学宁夏草牧业工程技术研究中心,宁夏 银川 750021;3. 宁夏大学农业农村部饲草高效生产模式创新重点实验室,宁夏 银川 750021;4. 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川 750021;5. 宁夏贺兰山国家级自然保护区管理局,宁夏 银川 750021)
植物群落的生物量作为生态系统功能的重要组成部分,在体现生态系统功能、植被生产力与形成群落结构方面发挥着重要作用[1]。其中,草本层植物占领生态位较小,对环境变化响应敏锐[2],常被认为是衡量生态系统稳定性以及功能恢复的可靠指标[3];灌木生物量不仅能反映灌木与乔木幼苗竞争生存环境的能力,更能反映灌丛生态系统对生存环境的适应能力[4]。植被与土壤是一个紧密联系、相辅相成的系统:一方面,土壤是植被生长发展的基础,为植被生长提供必要的养分与环境;另一方面,植被能够有效保持水土,减少土壤水分流失,同时促进土壤养分循环[5]。土壤水分是植物赖以生存的重要基础,不仅影响土壤特征和植物生长,并能通过影响植物的蒸腾作用和光合过程来调整生态系统中的水分、物质、能量的流动和循环[6-7]。植被类型、植被覆盖度以及地形变化等因子的变化也会导致土壤水分空间分布的明显差异[8-10]。山地生态系统中,海拔变化引起的温度、降水环境的变化不仅是影响山地生态系统植被类型与植被生物量的重要因素[11],并能影响土壤水分获得补给与蒸发耗损[12]。同时,植被通过根系对土壤产生挤压、分割与穿插作用,对土壤结构产生重要的影响[13],进而影响土壤的水分含量。相关研究表明,海拔、土壤水分以及土壤颗粒组成等因子均能显著影响植被生物量。刘哲等[14]在青藏高原研究发现,植被地上生物量随海拔升高线性降低;王金兰等[15]研究得出灌丛和灌丛下草本生物量随海拔升高呈先增高后降低的趋势,呈单峰格局;祁连山北坡天然草地、青藏高原高寒草甸草地的植被生物量与土壤含水量呈显著正相关[16-17],Li 等[18]在青藏高原高山草甸研究发现,土壤水分和土壤颗粒组成是决定植被地上生物量的关键因子。
我国是一个多山国家,山地面积大,分布广,山地是我国生态与经济建设的重要组成部分[19]。贺兰山地处我国西北内陆,作为区分我国季风区与非季风区、温带草原与荒漠草原等重要气候和植被条件的地理分界线,对于维护西北乃至整个西部地区的生态安全具有重要的意义[20]。近年来,对于贺兰山山地生态系统已有广泛研究,主要涉及土壤理化性质[21-22]、土壤微生物群落[23]、微生物量碳氮磷[24]以及土壤酶活性[25]等领域。但在垂直海拔梯度上研究植被生物量、土壤分形特征对土壤水分响应的研究鲜有报道。基于此,研究贺兰山不同海拔5 种典型植被带植被地上生物量、土壤分形特征和土壤水分含量的分布特征以及相互关系,以期为贺兰山山地生态系统植被与土壤状况提供基础数据累积,也为贺兰山草地合理利用和可持续发展提供科学依据。
1 材料与方法
1.1 研究区概况
贺兰山位于阿拉善高原与银川平原之间(38°27′-39°30′ N,105°41′-106°41′ E)。山体由西南向东北方向延伸,北起内蒙古阿拉善左旗,南至宁夏中卫市,主峰敖包疙瘩3556.15 m。贺兰山地处内陆,气候类型属于典型的大陆性气候,具有明显的山地气候特征,年均气温-8 ℃,年均降水量481.1 mm,降水具有明显的垂直分异特征。降水量主要集中于6-8 月,占全年降水量的60%~80%。土壤质地(图1)主要为粉质壤土;植被具有明显的垂直分布规律,从下到上依次为荒漠草原、浅山灌丛、山地针叶林、亚高山灌丛草甸与高山草甸。
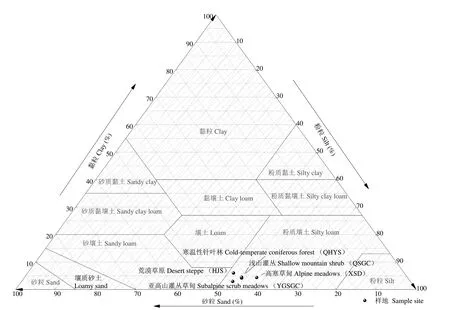
图1 研究区土壤质地Fig.1 Soil texture at sampling sites
1.2 样品采集与测定
于2021 年7 月下旬在贺兰山苏峪口海拔1300~2900 m 的区域,沿海拔选取1350 m 处以短花针茅(Stipa breviflora)为建群种的洪积扇荒漠草原(desert steppe,HJS),1800 m 处以蒙古扁桃(Prunus mongolica)为建群种的浅山灌丛(shallow mountain shrub,QSGC),2450 m 处以青海云杉(Picea crassifolia)为建群种的寒温性针叶林(cold-temperate coniferous forest,QHYS),2600 m 处以白毛银露梅(Potentilla glabra)与扁穗冰草(Agropyron cristatum)为建群种的亚高山灌丛草甸(subalpine scrub meadows,YGSGC)以及2900 m 处以高原嵩草(Kobresia pusilla)为建群种的高寒草甸(alpine meadows,XSD)5 种具有代表性的典型植被带,在每个研究区布设3 个样地,每个样地之间间隔50 m 以上,并在每个样地内随机设置3 个1 m×1 m 的草本样方;在有灌丛的研究区随机设置3个5 m×5 m 的灌木样方。样地基本信息如表1 所示。

表1 研究区基本信息Table 1 Basic information of the study areas
完成植被调查后采用齐地刈割的方法获取草本植被地上生物量(herb aboveground biomass, H-AGB)。在每个灌木样方中随机选取1 株灌木作为标准株(考虑其代表性),选取标准株的标准枝,用枝剪获取标准枝后装入档案袋标记好标准枝对标准株的占比后带回实验室,在实验室中置于恒温烘箱65 ℃条件下烘至恒重,计算标准枝干重、标准株干重以及样方内单位面积灌木地上生物量(shurb aboveground biomass, S-AGB)。将共同拥有草本植被地上生物量和灌木地上植被生物量的样地的两种植被类型的地上生物量求均值,计算得总平均生物量(total mean biomass, T-AGB)。
采用烘干法测定土壤水分。以梅花状采样法在每个样方中设置5 个取样点,去除各取样点地表枯落物以及石块等杂物后用直径为4 cm 的土钻对0~30 cm 深度的土壤进行取样,将每个样方内5 个取样点的土壤样品混合均匀后作为该样方的土壤样品,装入自封袋密封后带回实验室进行土壤水分含量的测定。因海拔1350 m 处为荒漠草原,土壤层厚度小,土壤层以下为坚硬石块,故在1350 m 荒漠草原研究区只取深度为0~10 cm 的土壤样品。
使用Mastersizer 2000 激光颗粒分析仪(Malvern,英国)进行土壤粒径的分析。仪器测量范围为0.01~2000.00 μm。本研究根据美国农业部(U.S. Department of Agriculture, USDA)制定的土壤粒径分级标准划分土壤颗粒组成:黏粒(<2 μm)、粉粒(2~50 μm)、极细砂粒(50~100 μm)、细砂粒(100~250 μm)、粗砂粒(250~2000 μm)。
通过世界气候网站(http://worldclim.org/)获取贺兰山年降水量(mean annual precipitation, MAP),年均温(mean annual temperature, MAT),通过ArcGIS 软件(ESRI,Redlands,美国)进行样地气候数据的提取。
1.3 土壤分形维数计算
由激光粒度仪(Mastersizer 2000,英国)获得土壤颗粒体积分布数据,采用王国梁等[26]提出的土壤颗粒体积分形模型进行分形维数的计算:
式中:Dv为土壤颗粒体积分形维数值;Vi为直径小于di的土壤颗粒的体积;V0为土壤颗粒的总体积;di是某粒径区间上下限的算术平均值;dmax为粒径分级中的最大粒径值。
1.4 数据处理
所有统计分析均在GraphPad Prism8.0 中完成,采用单因素方差分析(One way-ANOVA)对植被生物量、土壤水分含量、土壤颗粒组成与分形维数在不同海拔之间的差异性进行分析,同时采用最小显著差异法(LSD)检验差异显著性;采用Pearson 相关分析、冗余分析(redundancy analysis, RDA)和方差分解分析(variance partitioning analysis, VPA)对植被生物量、土壤水分含量与土壤颗粒组成、分形维数之间的关系进行分析。本研究数值为平均值±标准误(mean±SD)。
2 结果与分析
2.1 不同海拔植被生物量垂直分布特征
草本植被生物量沿海拔梯度呈明显的垂直分异特征(图2a)。随着海拔的升高,植被生物量呈“降低-增高-降低”的变化趋势,草本植被生物量由大到小依次为YGSGC>HJS>XSD>QSGC>QHYS,YGSGC 样地生物量为131.32 g·m-2,与其他海拔样地的植被生物量间均具有显著差异性(P<0.05);在QHYS 样地具有最小值,为26.40 g·m-2,与HJS 样地有显著差异(P<0.05)。灌木植被地上生物量随海拔变化并未表现出明显的差异性(图2b)。在HJS 样地,灌木植被地上生物量最小,QSGC 样地则最大,整体随海拔升高先升高后降低,呈单峰格局。

图2 各样地草本与灌木地上生物量Fig.2 Herb and shrub aboveground biomass of each study areas
对不同海拔样地草本植被地上生物量和灌木生物量进行平均以后得到单位面积总平均生物量(图3)。其中QHYS 样地主要优势植物为乔木青海云杉,故在计算总平均生物量时以林下草本生物量为主;XSD 样地无灌木,故以草本植被地上生物量为样地总平均生物量。不同海拔梯度样地间存在显著差异(P<0.05),YGSGC 样地和QSGC 样地总平均生物量分别为162.25 和121.55 g·m-2(图3),显著高于其他样地(P<0.05),QHYS 样地总平均生物量最低,为26.40 g·m-2,显著低于其他样地(P<0.05)。整体而言,草本植被与灌木地上植被总平均生物量随海拔升高呈“M”型变化趋势。

图3 各样地总平均生物量Fig.3 Total mean biomass of each study areas
土壤水分含量随海拔升高整体呈增加趋势(图4)。各样地土壤水分含量由大到小依次为:XSD>QHYS>YGSGC>QSGC>HJS。其中XSD 样地土壤水分含量为13.19%,显著高于YGSGC、QSGC 和HJS 样地(P<0.05);与QHYS 样地间无显著差异。HJS 样地土壤水分含量最低,为0.15%,显著低于其他样地(P<0.05);QHYS 与YGSGC 样地间土壤水分含量差异较小,且不显著。
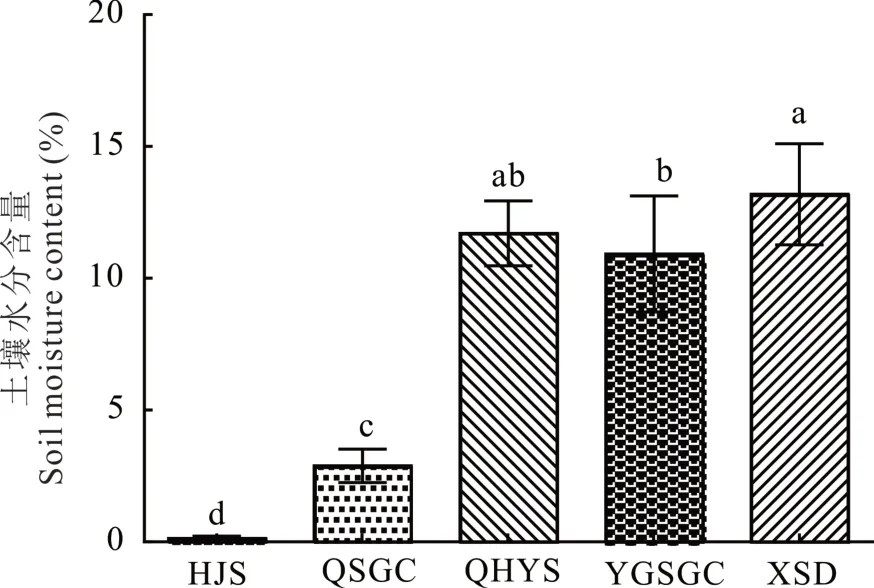
图4 各样地土壤水分含量Fig.4 Soil moisture content of each study areas
2.2 土壤颗粒组成的垂直分布特征
贺兰山土壤颗粒主要以粉粒和极细砂粒为主(图5a),整体上按体积分数由大到小依次为:粉粒>极细砂粒>细砂粒>粗砂粒>黏粒,颗粒组成随海拔升高逐渐变细。粉粒为贺兰山土壤最主要的组成成分,在研究区样地中,含量均达到50%以上,XSD 样地粉粒含量高达58%;极细砂粒含量次之,其含量为17%~31%。YGSGC 样地黏粒与粗砂粒含量均为3%,占比较小;HJS 样地粗砂粒含量较其他样地粗砂粒含量高,为12%。QSGC 样地与QHYS 样地的土壤颗粒组成含量相同,说明其土壤颗粒组成具有一定的相似性。各样地土壤分形维数数值相差不大但具有显著差异(P<0.05,图5b),具体为:HJS>QHYS>XSD>QSGC>YGSGC。其中HJS 土壤分形维数显著高于QSGC、YGSGC 和XSD 样地(P<0.05),但与QHYS 样地无显著差异;QSGC、YGSGC 和XSD 样地间分形维数无显著差异。

图5 各样地土壤颗粒组成和分形维数Fig.5 Soil particle composition and fractal dimension
2.3 不同海拔高度植被生物量、土壤分形特征与土壤水分的相关关系
年均降水量和土壤水分含量均与海拔呈极显著正相关关系(P<0.01,图6);粉粒含量以及极细砂粒含量分别与海拔呈显著(P<0.05)和极显著(P<0.01)正相关关系;同时,粉粒含量以及极细砂粒含量也分别与土壤水分含量呈显著(P<0.05)和极显著(P<0.01)正相关关系;土壤水分含量与黏粒和土壤分形维数呈显著负相关(P<0.05),与细砂粒和粗砂粒呈极显著负相关关系(P<0.01)。

图6 植被生物量与各因子的Pearson 分析Fig.6 Pearson analysis of vegetation biomass with each factors
将草本生物量、灌木生物量和总平均生物量作为响应变量,土壤颗粒组成特征及土壤水分含量作为解释变量,首先进行除趋势对应分析(detrended correspondence analysis, DCA)得到4 个排序轴最大梯度为1.0,故进行冗余分析研究植被生物量与土壤相关指标的关系。RDA 排序轴前两轴土壤因子对植被生物量的解释量分别为39.90%、28.51%,累计解释量为68.41%(图7),说明土壤因子解释了植被生物量的大部分变异,排序结果可靠。选择前2 轴绘制植被生物量与土壤因子的二维空间排序图。从排序轴上看,排序轴1 主要反映土壤VFS、D、C 等的影响,排序轴2 主要反映土壤FS 等的影响。根据箭头方向与夹角分析可知,植被生物量与MAP、VFS、海拔、SMC 均呈正比;与D、C、MAT 和FS 呈反比;H-AGB 与土壤S 呈正比;但S-AGB 和T-AGB 与土壤S 呈反比。C与SMC 的解释量较高,分别为30.1%和21.0%,且达到了显著水平(P<0.05,表2)。

表2 RDA 分析Table 2 RDA analysis
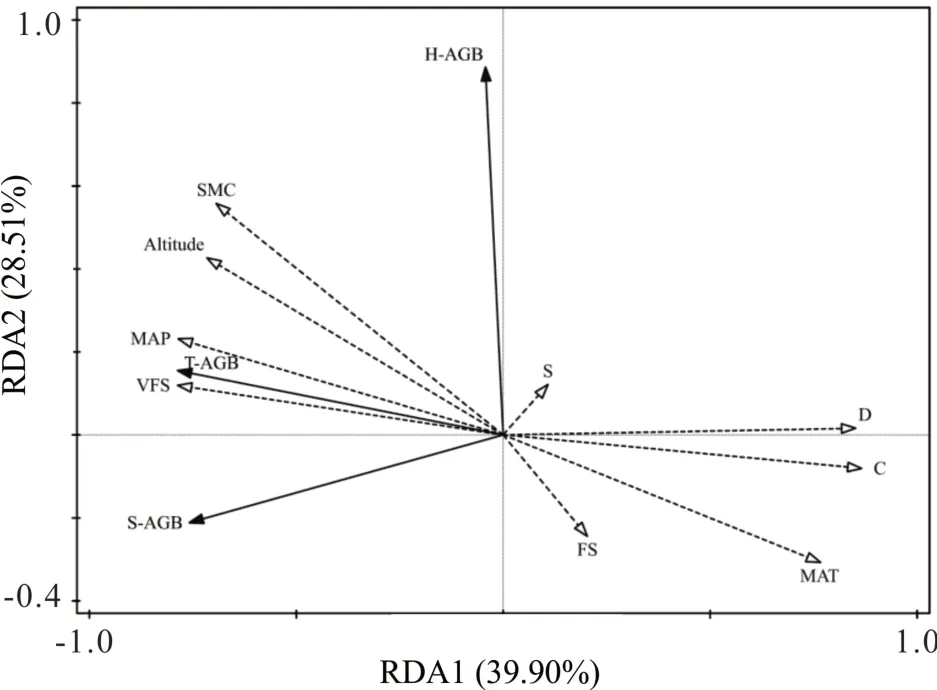
图7 植被生物量与土壤因子的RDA 排序Fig.7 RDA ranking of vegetation biomass and soil factors
3 讨论
3.1 海拔对植被生物量与土壤水分的影响
生物量作为生态系统最基本的数量特征,能够反映群落或生态系统获取能量的能力,是研究生态系统物质循环与能量流动的基础[27]。在本研究中,草本植被生物量随海拔变化呈“降低-升高-降低”的变化趋势,此结果与苏闯等[28]在贺兰山西坡的研究结果一致,即山地生态系统的植被生物量符合“中部膨胀”理论;但海拔对植被生物量的影响并不显著,这与韩彬等[29]在内蒙古的研究结果一致。在山地生态系统中,植被生物量的变化不仅受到海拔引起的水热因子的制约,还受到“环境约束”与“资源竞争”机制的影响[30]。低海拔区域干旱少雨,养分相对贫瘠,仅有少数耐旱性较强的灌木种与草本植物种生存,且因土壤水分限制以及种间竞争关系,使得植株高度、叶面积等植物性状受到影响,制约了植被生物量的积累[27];中海拔区域温度适宜,降水充沛,土壤养分充足,优越的水热条件使得植被具有更良好的长势,形成更为高大的植株体,因此,在此处能够获得最大的植被生物量;而在高海拔的区域,温度较低,风速较大,虽土壤水分蒸散量小,含量充足,但由于降水的冷却效应以及温度较低时常形成冻土,对植被的生长以及生物量的积累产生了负面效应[31],同时,海拔升高引起温度下降,植物体会将更多的生物量分配到地下部分,使得植被地上生物量降低。灌木生物量在HJS 样地较低,主要是由于荒漠草原土壤养分较低,水热条件较差,灌木不适宜生长;而随着海拔的升高,水热条件逐渐变得优越,土壤养分条件较海拔较低的HJS 区域肥沃,更适宜灌木生长[32]。同时,灌木层能够对草本层起到庇护的作用,促进草本层的生长,从而影响地上植被生物总量[33]。土壤水分状况是气象、植被和地形等多种环境因子综合作用的结果,具有一定的复杂性[12]。本研究中,随着海拔的增加,土壤水分含量整体呈增加的变化趋势。分析原因,首先认为,在山地生态系统中,降水量与海拔呈正相关关系,土壤水分含量对降水存在着明显的响应[34];其次,高海拔地区温度较低,削弱了土壤水分的蒸散与蒸发,并且深层土壤的冻融作用能够影响土壤水分的迁移和再分配[35],因此海拔越高,土壤水分含量越高。但本研究中,QHYS 样地土壤水分含量略高于YGSGC 样地,出现该现象可能是植被与土壤水分含量相互作用,植被类型不同,导致土壤水分含量沿海拔变化出现了部分的异质性[36]。本研究QHYS 样地的优势植物为青海云杉,青海云杉林郁闭度较大,为林下土壤尤其是深层土壤提供了良好的遮阴作用,减少了到达土壤表面的辐射,减少了土壤水分蒸发[37];再者,青海云杉作为涵养水源的主要造林树种,其立地土壤具有较好的孔隙度和持水能力,在降水事件发生时能够维持更多的水分[38]。而YGSGC 样地植物物种较多,对土壤水分竞争激烈,且更高海拔的YGSGC 样地面临着更强烈的太阳辐射与风力作用,使得土壤水分留存不易,其含量略低于QHYS 样地。
3.2 海拔对土壤颗粒组成和分形维数的影响
土壤颗粒组成受其成土母质、植被、海拔等因子的共同影响[39]。在本研究中,土壤粉粒含量和极细砂粒与海拔显著正相关,黏粒、细砂粒、粗砂粒含量以及分形维数值与海拔显著负相关。这是由于随着海拔的升高,植被覆盖增加,能够有效减少风蚀作用,降低了细小颗粒的损失[40]。另外,随着海拔的增加,植物凋落物覆盖度与厚度增加,增加了对土壤的保护作用,还增强了土壤微生物群落能量输入,使得微生物的生命活动增强,细化土壤颗粒的能力增强[41],因此表现出土壤颗粒组成沿海拔升高向着立即变小的方向演变。在本研究中,随着海拔的升高,土壤分形维数呈降低的趋势。这是由于土壤的风化发育程度会随海拔升高而降低[42];而非表层土壤由于表层植被凋落物以及表层土壤的保护作用,颗粒组成保持着稳定性。随着土壤深度的增加,分形维数表现出与海拔的正相关关系,这与刁二龙等[43]的研究结果一致,海拔通过影响地表水热条件来影响土壤颗粒的相互转化,对分形维数产生一定的影响。本研究中,土壤分形维数整体上与粒径较大的颗粒呈负相关,与粒径较小的颗粒呈正相关。这与兰龙焱等[44]的研究结果一致,因此在一定程度上可以用土壤分形维数来评估区域质地以及水土流失的情况。
3.3 植被生物量、土壤分形特征与土壤水分的关系
植被可通过改善土壤颗粒组成,如增加黏粒与粉粒的含量,来遏制土壤侵蚀,提高土壤质量[45-46],土壤颗粒通过固持水分与养分,供给植被生长发育所必需的资源条件。本研究发现,植被生物量并未表现出与土壤水分的强烈的相关关系,说明在贺兰山地区,土壤水分可能并不是制约植被积累生物量的主要环境因子。根据RDA 分析结果,推测土壤黏粒通过吸附土壤中养分与水分,对植被生物量的累积影响更大[47]。植被通过根系作用,能够对土壤颗粒产生挤压和碎裂作用,影响土壤的分形特征[13]。本研究中,粉粒与极细砂粒都表现出了与土壤水分含量的显著相关性,该研究结果与仓木拉等[48]的研究结果部分相同。有研究指出,粉粒的比表面积较大,具有较强的表面吸附能力,更易于吸附土壤中的养分与水分[49],因此,呈现出了一个正向相关结果。整体上,植被生物量受海拔变化引起的降水变化以及与土壤水分含量紧密相关的土壤黏粒的影响,在海拔梯度变化上呈明显的垂直差异变化,因此,贺兰山植被生物量受多种环境因子直接或间接综合作用。
4 结论
本研究基于贺兰山地区不同海拔的5 种典型植被带的草本、灌木和总平均生物量以及土壤水分含量与植被分形特征,分析了海拔梯度下植被生物量、土壤因子的垂直分布特征及相互关系,得出以下结论:1)草本植被地上生物量随海拔变化未表现出明显的增减趋势,而灌木植被生物量和总平均生物量沿海拔升高整体呈增加的变化趋势,在本研究样地中高海拔区域生物量略有增加,此变化印证了“中部膨胀”理论,说明中部植被条件较好,生物量较高。2)贺兰山土壤颗粒主要以粉粒和极细砂粒为主,土壤水分含量与海拔和降水呈极显著正相关关系,说明海拔变化以及海拔引起的水热变化是影响土壤水分含量的重要因子。3)植被生物量未体现出与海拔以及土壤水分含量的相关性,但与土壤颗粒组成表现出了强相关性,即在贺兰山地区,降水以及固持养分与水分的土壤黏粒对植被的影响较海拔和土壤水分的影响更大。该研究结果为认识贺兰山植被生物量与土壤颗粒组成以及土壤水分提供了基础数据,为贺兰山生态环境保护与治理提供了一定参考。
