夏桑菊低聚糖对益生菌增殖作用及其酶解工艺优化研究
2023-12-07黄志云孙维广黎尔纳万安凤邹宇晓高建胜
黄志云 孙维广* 黎尔纳 万安凤 邹宇晓 高建胜
1.广州白云山星群(药业)股份有限公司,广东 广州 510288;2.广东省农业科学院蚕业与农产品加工研究所,农业农村部功能食品重点实验室,广东省农产品加工重点实验室,广东 广州 510610
中药渣是中药提取后的固体废弃物,随着中药产业的飞速发展,中药渣的排放量也日益上升,据统计,中药渣的年排放量可达3000万吨。在生产中该类废弃物往往直接排放,造成资源浪费的同时,对生态环境有较大的破坏[1-3]。
夏桑菊颗粒源自清代著名温病学家吴鞠通《温病条辨》的经典名方“桑菊饮”,主要由夏枯草、桑叶和野菊花配伍而成。现代研究表明夏枯草含有三萜类、多糖类、黄酮类、甾体类与有机酸类化合物等成分,具有降血糖、降血压、调节免疫系统、抗肿瘤、抗炎抗菌及抗病毒等药理作用[4]。桑叶中含酚类及黄酮类、生物碱类、氨基酸、多糖和甾类化合物,具有降血糖、抗炎、抗菌、抗病毒、抗衰老及抗癌等生物活性[5]。野菊花含有挥发油、黄酮类、多糖类、有机酸类、氨基酸以及微量元素等化学成分,有抑菌、抗炎镇痛、抗肿瘤活性、抗氧化等生理活性[6]。由组方及其生理活性可知,夏桑菊药渣在某意义来说也是一种“中药抗生素”。
夏桑菊颗粒生产时,为了保证产品质量均一稳定,需要“水提”物质进行“醇沉”处理,醇沉后的上清液作为生产原料,醇沉物为废弃物,但废弃物中却含有丰富的多糖、多酚和黄酮类物质等成分。我司每年产生约几千吨夏桑菊药渣,传统的填埋处理方式经济价值低,而且造成药效成分浪费。本文拟采用生产夏桑菊颗粒过程中的“水提醇沉”物为原料,经生物酶解处理后获得夏桑菊低聚糖,考察其对益生菌的增殖作用,并优化酶解工艺,为未来将夏桑菊药渣开发成高价值的“减抗替抗”动物营养制剂提供参考。
1 仪器与材料
1.1 仪器 THZ-D型台式恒温振荡器(江苏省太仓市华美生化仪器厂);HWS26型电热恒温水浴锅、DHP-9082型电热恒温培养箱(上海一恒科学仪器有限公司);SW-CJ-2FD型洁净工作台(苏州安泰空气技术有限公司)。
1.2 材料 夏桑菊醇沉渣由广州白云山星群(药业)股份有限公司提供,利用苯酚-硫酸法检测其多糖浓度约为80 mg/mL。果胶酶(1000 U/mg)、纤维素酶(400 U/mg)、半纤维素酶(2 U/mg)、淀粉酶(4 U/mg)、葡聚糖酶(50 U/mg)、木聚糖酶(6000 U/mg)、甘露聚糖酶(50 U/mg)、糖化酶(100 U/mg)购于广州市齐云生物技术有限公司。其他试剂均为国产分析纯。
植物乳杆菌 (Lactobacillusplantarum,GIM1.191)、肠膜明串珠菌(Leuconostocmesenteroides,GIM1.774)、干酪乳杆菌(Lactobacilluscasei,GIM1.411)、鼠李糖乳杆菌(Lactobacillusrhamnosus,ATCC53103)、青春双歧杆菌(Bifidobacteriumadolescentis,ATCC15703)保藏于广东省农业科学院蚕业与农产品加工研究所蚕桑南药与微生物资源加工利用研究室。
2 方法与结果
2.1 多糖浓度的测定 称取干燥至恒重的葡萄糖500 mg,取少量蒸馏水溶解,并定容到500 mL,摇匀,即得质量浓度为1.00 mg/mL的葡萄糖标准溶液。采用苯酚-硫酸比色法,分别吸取 0.00 mL、0.50 mL、1.00 mL、1.50 mL、2.00 mL、2.50 mL葡萄糖标准溶液,定容到25 mL,得到不同浓度的葡萄糖稀释液,分别取1.00 mL稀释液于10 mL试管中,各加入质量浓度为5%的苯酚溶液1 mL,再迅速加入浓硫酸5 mL,充分震荡摇匀,室温静置冷却30 min,在490 nm下测吸光值。以吸光度为纵坐标,葡萄糖质量浓度为横坐标,制作标准曲线。吸取适当稀释的夏桑菊多糖溶液 1 mL 于 10 mL 试管中,以1 mL蒸馏水作为空白对照,按标准曲线的操作进行显色,于490 nm处测定吸光值,并计算多糖含量。结果显示,本试验所用夏桑菊醇沉渣中多糖浓度为80 mg/mL。
2.2 不同生物酶酶解制备的夏桑菊低聚糖促益生菌生长活性的测定 夏桑菊低聚糖制备:往夏桑菊醇沉渣中分别添加8种生物酶(果胶酶、纤维素酶、半纤维素酶、淀粉酶、葡聚糖酶、木聚糖酶、甘露聚糖酶、糖化酶),使其酶活力达到1000 U/mL,50 ℃酶解3 h,得到生物酶解制备的夏桑菊低聚糖,灭酶(沸水浴10 min)后121 ℃高压灭菌20 min,冷却后于4 ℃储存待用。
夏桑菊低聚糖促益生菌生长活性的测定:分别将夏桑菊低聚糖和商品化益生元(IMO、GOS)添加到MRS液体培养基,使其质量终浓度达到10 mg/mL。向培养基中接种1%过夜培养的益生菌菌液,37℃、180 r/min震荡培养过夜,按下公式计算增殖率。
益生菌增殖率=低聚糖与益生菌菌共培养后菌落数/空白对照组菌落数×100%
结果显示,不同生物酶对多糖的降解效果不同。图1-A中,利用半纤维素酶制备的夏桑菊低聚糖对植物乳杆菌的促增殖效果最好,增殖率为(156±9.8)%,其次为果胶酶,增殖率为(141±12.3)%,两组与阴性对照组相比,差异显著(P<0.05)。图1-B中,利用木聚糖酶制备的夏桑菊低聚糖对肠膜明串珠菌的促增殖效果最好,增殖率为(168±13.8)%,其次为葡聚糖酶,增殖率为(153±14.3)%,两组与阴性对照组相比,差异显著(P<0.05)。图1-C中,利用葡聚糖酶制备的夏桑菊低聚糖对干酪乳杆菌的促增殖效果最好,增殖率为(213±17.3)%,其次为果胶酶,增殖率为(189±15.7)%,两组与阴性对照组相比,差异显著(P<0.05)。图1-D中,利用葡聚糖酶制备的夏桑菊低聚糖对鼠李糖乳杆菌的促增殖效果最好,增殖率为(265±14.5)%,其次为半纤维素酶,增殖率为(226±27.3)%,两组与阴性对照组相比,差异显著(P<0.05);优于未酶解的夏桑菊多糖(151±4.2)%和商品化益生元低聚异麦芽糖(173±8.3)%、低聚半乳糖(182±9.2)%。图1-E中,利用葡聚糖酶制备的夏桑菊低聚糖对青春双歧杆菌的促增殖效果最好,增殖率为(176±15.3)%;其次为果胶酶,增殖率为(169±14.3)%,两组与阴性对照组相比,差异显著(P<0.05)。
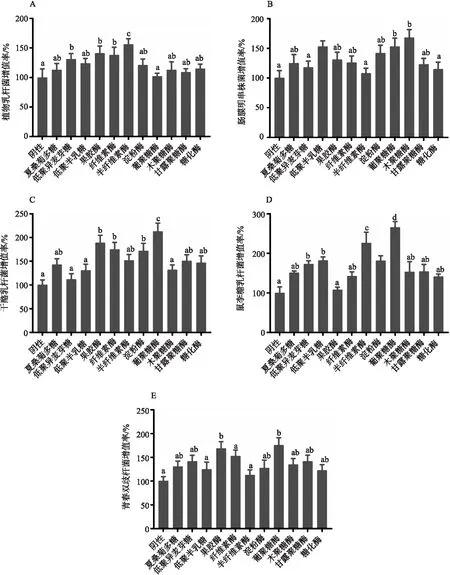
注:柱形图中相同字母表示差异不显著(P>0.05);不同字母表示差异显著(P<0.05)。
综上,8种不同酶酶解制备的夏桑菊低聚糖对5种益生菌都具有较好的促增殖作用,其中利用葡聚糖酶制备的夏桑菊低聚糖对鼠李糖乳杆菌的增殖效果最好,后续将以此菌为增殖效果为指标,优化葡聚糖酶生物酶解夏桑菊低聚糖工艺参数。
2.3 夏桑菊低聚糖浓度对其促鼠李糖乳杆菌生长活性的影响 将夏桑菊低聚糖添加到MRS液体培养基中,使其质量终浓度分别达到40 mg/mL、20 mg/mL、10 mg/mL、5 mg/mL、2.5 mg/mL。向培养基中接种1%过夜培养的鼠李糖乳杆菌菌液,37 ℃、180 r/min震荡培养过夜,计算增殖率。结果显示,低聚糖质量终浓度在2.5~40 mg/mL范围内,鼠李糖乳杆菌的增殖率随浓度的增大呈先上升后平缓下降的趋势,当质量终浓度为 10 mg/mL 时增殖率达到最大值(273±16.4)%,如图2。

图2 夏桑菊低聚糖浓度对鼠李糖乳杆菌增殖率的影响图
2.4 夏桑菊低聚糖生物酶解制备工艺的单因素优化 以鼠李糖乳杆菌增殖率为指标,添加低聚糖质量终浓度为10 mg/mL,选择不同酶添加量、酶解温度、酶解时间,研究其对夏桑菊低聚糖酶解工艺的影响。实验参数见表2。
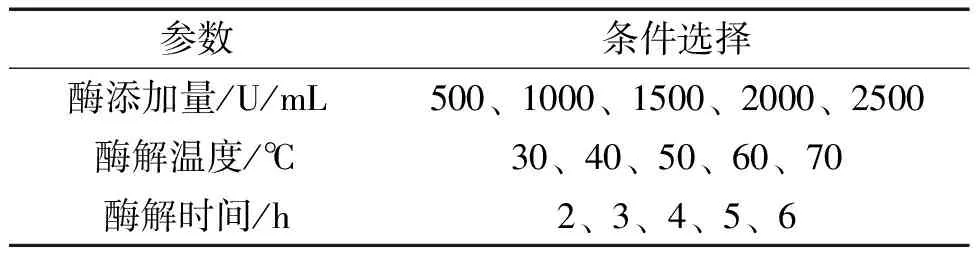
表2 生物酶解工艺参数
2.4.1 不同酶添加量制备的夏桑菊低聚糖对鼠李糖乳杆菌增殖率的影响 如图3所示,葡聚糖酶添加量在500~2500 U/mL时制备的夏桑菊低聚糖,鼠李糖乳杆菌的增殖率呈先增后减的趋势。酶添加量为1500 U/mL时,鼠李糖乳杆菌的增殖率达到了(320±17.2)%,而酶添加量大于 1500 U/mL 时,增殖率开始逐渐降低。
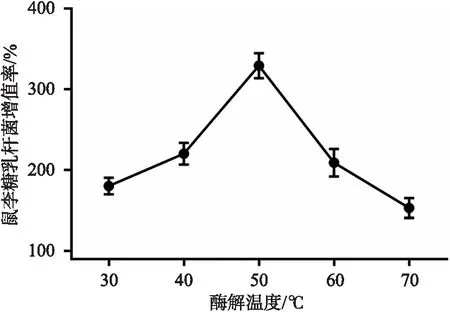
图4 酶解温度对鼠李糖乳杆菌增殖率的影响
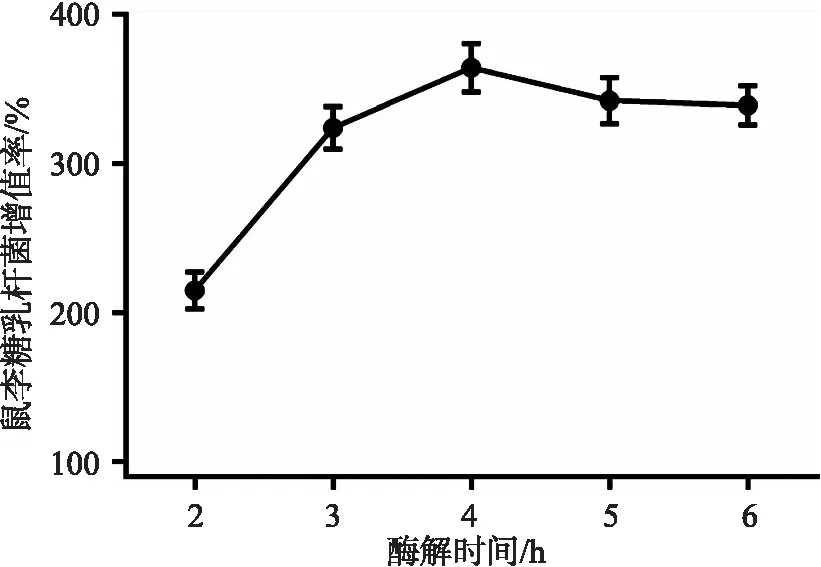
图5 不同酶解时间对鼠李糖乳杆菌增殖率的影响
2.4.2 不同酶解温度制备的夏桑菊低聚糖对鼠李糖乳杆菌增殖率的影响 葡聚糖酶添加量 1500 U/mL,夏桑菊低聚糖对鼠李糖乳杆菌增殖随着温度变化先增后降,酶解温度50 ℃时增殖率最高,为(329±15.6)%。
2.4.3 不同酶解时间制备的夏桑菊低聚糖对鼠李糖乳杆菌增殖率的影响 酶解时间为2~6 h,增殖率先直线上升后慢速下降,并在4 h时达到最高值(364±16.4)%。综上,从益生活性考虑,选取葡聚糖酶添加量1500 U/mL、酶解温度50 ℃和酶解时间4 h为较优酶解方案。
3 讨论
生物酶解法在中药提取中有广泛应用[7-10]。其原理为:通过选用一些恰当的酶类与药用植物细胞使细胞壁及细胞间质中的纤维素、半纤维素、果胶等物质降解破坏细胞壁的致密结构,引起细胞壁及细胞间质结构产生局部疏松、膨胀、崩溃等变化,减小细胞壁、细胞间质等传质屏障对有效成分从胞内向提取介质扩散的传质阻力,从传质角度促使有效成分提取率提高[11]。
在考察夏桑菊低聚糖浓度对其促鼠李糖乳杆菌生长活性的影响试验中,当低聚糖质量终浓度在2.5~40 mg/mL范围内时,鼠李糖乳杆菌的增殖率随浓度的增大呈先上升后平缓下降的趋势,当质量终浓度为10 mg/mL时增殖率达到最大值,质量终浓度继续增大时,增殖率随浓度增大而降低,可能是因为浓度过高,从而导致渗透压过高,细胞破裂,增殖率降低。
在考察不同酶添加量制备的夏桑菊低聚糖对鼠李糖乳杆菌增殖率的影响试验中,葡聚糖酶添加量在500~2500 U/mL时制备的夏桑菊低聚糖,鼠李糖乳杆菌的增殖率呈先增后减的趋势。酶添加量为1500 U/mL时,鼠李糖乳杆菌的增殖率达到最大值,而酶添加量大于1500 U/mL时,增殖率开始逐渐降低。这可能是因为低聚糖的结构影响其活性作用的发挥,具有一定分子量的低聚糖才具有促生长活性,过度酶解的低聚糖小片段,其促增殖活性不佳[12]。
在考察不同酶解温度制备的夏桑菊低聚糖对鼠李糖乳杆菌增殖率的影响的试验中,酶解温度在30~70 ℃时制备的夏桑菊低聚糖,鼠李糖乳杆菌增殖率呈先增后减趋势,温度50℃时增殖率最大。因为酶的活性与反应温度有关,且存在最适温度。制备的低聚糖对鼠李糖乳杆菌的促生长作用并不是随着酶解温度的升高而呈直线上升趋势,而是在适宜温度值时鼠李糖乳杆菌的生长活性存在较优值。
在考察不同酶解时间制备的夏桑菊低聚糖对鼠李糖乳杆菌增殖率的影响试验中,酶解时间为 2~6 h,增殖率先直线上升后慢速下降,并在4 h时达到最高值(364±16.4)%,这种情况可能是,随时间的延长,葡聚糖酶与糖链作用的机会更完全,产生更多的活性低聚糖,促进鼠李糖乳杆菌的生长繁殖;随着酶解时间的继续延长,酶解产物的组成发生改变,不利于鼠李糖乳杆菌的增殖。
实验中,8种不同生物酶酶解制备的夏桑菊低聚糖对5种益生菌都具有较好的促增殖作用,其中利用葡聚糖酶制备的夏桑菊低聚糖对鼠李糖乳杆菌的增殖效果最好;从益生活性考虑,选取葡聚糖酶添加量1500 U/mL、酶解温度50 ℃和酶解时间4 h为较优酶解方案。综上,研究提供的技术方案可为未来夏桑菊药渣的开发利用提供实验依据。
