植物甾醇酯对产蛋末期蛋鸡产蛋性能、蛋品质、肝脏抗氧化能力及卵黄前体物质合成的影响
2023-12-04杨凯丽郝隽毅邓圣庭杨伟光刘石鹏孙杰龙方热军
章 娜 杨凯丽 张 沙 郝隽毅 邓圣庭 杨伟光 刘石鹏 孙杰龙 方热军*
(1.湖南农业大学动物科学技术学院,长沙 410128;2.智慧牧业湖南省工程研究中心,长沙 410128;3.陕西省渭南市动物疫病预防控制中心,渭南 714000;4.湖南省鹏晖农牧有限公司,安化 413500;5.益阳市农业科学院,益阳 413000)
家禽产蛋过程中,卵黄前体物质主要包括极低密度脂蛋白(very low density lipoprotein,VLDL)和卵黄蛋白原(vitellogenin,VTG),其直接影响蛋禽的卵泡发育和产蛋性能。在鸡的排卵期,卵巢本身不产生内源性卵黄前体,由肝脏组装并合成的VLDL和VTG经血液循环至卵巢后,通过卵泡膜上的极低密度脂蛋白受体介导的胞吞作用被卵母细胞吸收利用,为卵泡发育提供营养物质[1-2]。高产母鸡在400日龄后通常会出现卵巢老化、产蛋量下降和蛋品质变差等问题[3]。蛋鸡在衰老过程中,雌激素作用和肝脏抗氧化机能逐渐衰退,肝脏分泌卵黄前体物质能力下降,导致产蛋性能下降[4]。近年研究发现,天然抗氧化物或植物提取物(如甜菜碱、山楂叶黄酮等)可缓解蛋鸡肝脏或卵巢氧化损伤,促进卵黄前体物质合成及相关基因的表达,从而提高蛋鸡的产蛋性能和蛋品质[5-7]。
植物甾醇(phytosterol,PS)又称植物固醇,是以环戊烷全氢菲为骨架的一类抗氧化剂,具有抗氧化、类雌激素和促生长作用,可有效调节机体氧化还原稳态失衡[8]。植物甾醇酯(phytosterol ester,PSE)由PS酯化而成,与PS相比,PSE更容易被动物肠道吸收[9]。本团队前期研究结果表明,饲粮中添加150~300 mg/kg PSE能够提高110日龄蛋鹌鹑的产蛋率,显著提高血清中总超氧化物歧化酶(total superoxide dismutase,T-SOD)、过氧化氢酶(catalase,CAT)和肝脏中谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-Px)活性,降低蛋黄中胆固醇含量[10]。PSE可否通过提高蛋鸡抗氧化应激能力,促进肝脏卵黄前体物质的生成,从而达到提高产蛋性能、改善蛋品质目的,其基于“肝脏-血液-卵巢信号轴”的产蛋机制尚未见报道。因此,本研究拟以产蛋末期(113周龄)蛋鸡为研究对象,从抗氧化的角度探究PSE对其产蛋性能和蛋品质的影响,并从卵黄前体物质VLDL、VTG合成的影响途径诠释其作用机制,以期为延长蛋鸡的产蛋周期、实现蛋鸡的“延养”、提高蛋鸡养殖经济效益提供技术方案。
1 材料与方法
1.1 试验设计
采用单因素试验设计,选用180只113周龄产蛋率[(77.22±2.50)%]相近的健康京粉六号蛋鸡,随机分为3个组,分别为CON组(饲喂基础饲粮,对照组)、100 PSE组(饲喂基础饲粮+100 mg/kg PSE)、200 PSE组(饲喂基础饲粮+200 mg/kg PSE),每组6个重复,每个重复10只鸡。预试期为7 d,正试期为56 d。试验结束后,每个重复取2只鸡屠宰。试验所用PSE产品由湖南某生物科技有限公司提供,PSE有效含量为15%。基础饲粮参照NRC(1994)、中国《鸡饲养标准》(NY/T33—2004)和《中国饲料成分及营养价值成分表(2020年第31版)》配制,其组成及营养水平见表1。
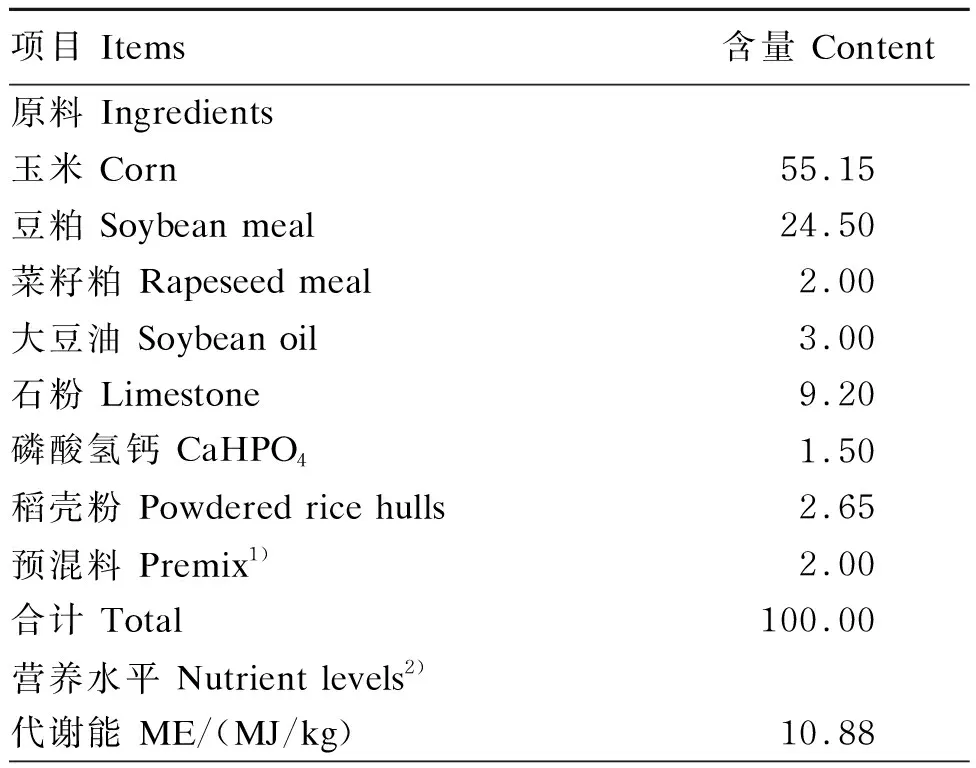
表1 基础饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of the basal diet (air-dry basis) %
1.2 饲养管理
饲养试验在湖南省益阳市鹏晖农牧有限公司进行,预试期观察蛋鸡产蛋数、采食量及死淘情况,及时调整。鸡舍为全封闭式,夏季采用湿帘和风机降温,环境温度控制在18~25 ℃。蛋鸡饲养均采用层叠式笼养模式,密度为5只/笼,2笼/重复。试验期间,每日饲喂粉料2次,使用乳头式饮水器自由饮水,按正常免疫程序进行免疫接种。
1.3 测定指标与方法
1.3.1 产蛋性能测定
以重复为单位每天记录产蛋数、蛋重、破/软壳蛋数和死亡淘汰鸡数目,按周统计各重复蛋鸡试验期间的耗料量,计算平均日采食量、产蛋率、平均蛋重、料蛋比、死淘率和破软壳率。
平均日采食量[g/(只·d)]=饲料消耗
总量/(存栏鸡只数×试验天数);
产蛋率(%)=(产蛋数/存栏鸡只数)×100;
平均蛋重(g)=产蛋总重量/总产蛋数;
料蛋比=饲料消耗总量/产蛋总重量;
死淘率(%)=(死淘数/总存栏数)×100;
破/软壳率(%)=(破/软壳蛋数/产蛋数)×100。
1.3.2 蛋品质测定
正试期每天10:00捡蛋,在试验第28天和第56天从每个重复挑选接近该重复平均重(不包括畸形、粗糙或破裂蛋)的合格鸡蛋4枚,每组共24枚。使用游标卡尺测量鸡蛋的纵径和横径,计算蛋形指数(蛋形指数=纵径/横径);使用蛋品质自动分析仪(EA-01型,以色列)测定蛋黄颜色、蛋黄直径、蛋白高度、蛋重和哈氏单位,电子天平称量蛋壳重量,计算蛋壳表面积(蛋壳表面积=4.68×蛋重2/3)。
1.3.3 血清生殖激素指标测定
采用酶联免疫吸附试验(ELISA)试剂盒(由上海酶联生物科技有限公司生产)测定血清中雌二醇(estradiol,E2)、催乳素(prolactin,PRL)、促卵泡素(follicle-stimulating hormone,FSH)、促黄体素(luteinizing hormone,LH)、孕酮(progesterone,Prog)水平,具体操作方法参照试剂盒说明书进行。
1.3.4 血清和肝脏抗氧化指标测定
采用试剂盒(由南京建成生物工程研究所生产)测定血清和肝脏中总抗氧化能力(total antioxidant capacity,T-AOC)和T-SOD、GSH-Px、CAT活性及丙二醛(malondialdehyde,MDA)含量,操作步骤严格按照试剂盒说明书进行。
1.3.5 血清和肝脏中VLDL、VTG含量测定
使用ELISA试剂盒(由上海酶联生物科技有限公司生产)对血清和肝脏中VLDL和VTG含量进行测定,操作步骤按照试剂盒说明书进行。
1.3.6 肝脏中抗氧化及卵黄前体物质合成相关基因检测
准确称取1 g肝脏组织样品置于装有液氮的研钵中充分研磨。按照Trizol试剂说明书对肝脏总RNA进行提取,使用Nanodrop 2000微量紫外分光光度计测定其浓度和纯度后,用EvoM-MLV反转录试剂盒(由湖南艾科瑞生物科技有限公司提供)合成cDNA,-20 ℃保存待测。利用Premier生物软件设计引物,引物序列由上海生工生物有限公司合成,详见表2。配制体积为10 μL的反应体系,于ABI-7900HT实时荧光定量PCR仪(Applied Biosystems,美国)上按照说明书进行一步实时荧光定量PCR。以β-肌动蛋白(β-actin)为内参基因,采用2-ΔΔCt法计算样品中目的基因mRNA的相对表达量。

表2 实时荧光定量PCR引物Table 2 Primers for real time quantitative PCR
1.4 数据处理与分析
试验数据采用SPSS 26.0统计软件进行单因素方差分析(one-way ANOVA),若有显著效应,则采用Duncan氏法进行多重比较,分析结果以平均值和均值标准误(SEM)表示。用GraphPad Prism 8软件作图。P<0.01为差异极显著,P<0.05为差异显著,0.05≤P<0.10表示差异有显著趋势,P>0.05表示差异不显著。
2 结果与分析
2.1 PSE对产蛋末期蛋鸡产蛋性能的影响
由表3可知,与CON组相比,100 PSE组和200 PSE组前期(1~4周)产蛋率分别提高6.12%和9.01%,差异显著(P<0.05);后期(5~8周)产蛋率均有所提高,但差异未达到显著水平(P>0.05);全期(1~8周)产蛋率分别提高6.37%和8.62%,差异显著(P<0.05)。100 PSE组前期和全期平均蛋重均显著高于CON组(P<0.05),其后期料蛋比显著低于CON组和200 PSE组(P<0.05)。200 PSE组后期平均日采食量较CON组提高3.56%,差异极显著(P<0.01)。各组间前期、后期以及全期死淘率、破/软壳率均无显著差异(P>0.05)。

表3 PSE对产蛋末期蛋鸡产蛋性能的影响Table 3 Effects of PSE on laying performance of laying hens during late laying period
2.2 PSE对产蛋末期蛋鸡蛋品质的影响
由表4可知,与CON组相比,100 PSE组第28天的蛋壳重量和第56天的蛋黄颜色均极显著提高(P<0.01);200 PSE组第28天和第56天的蛋黄颜色分别提高16.48%(P<0.01)和17.26%(P<0.01),蛋壳重量分别提高6.57%(P<0.01)和10.85%(P<0.05)。100 PSE组和200 PSE组第56天的蛋重和蛋壳表面积均显著高于CON组(P<0.05)。其他蛋品质指标各组间均无显著差异(P>0.05)。

表4 PSE对产蛋末期蛋鸡蛋品质的影响Table 4 Effects of PSE on egg quality of laying hens during late laying period
2.3 PSE对产蛋末期蛋鸡血清生殖激素指标的影响
由表5可知,与CON组相比,100 PSE组和200 PSE组血清中E2水平分别提高10.29%和14.77%,Prog水平提高10.15%和23.91%,差异均达到显著水平(P<0.05)。各组间血清中FSH和LH水平均存在极显著差异(P<0.01),其中200 PSE组FSH和LH含量较CON组分别提高38.46%和42.06%。

表5 PSE对产蛋末期蛋鸡血清生殖激素指标的影响Table 5 Effects of PSE on serum reproductive hormone indices of laying hens during late laying period
2.4 PSE对产蛋末期蛋鸡血清和肝脏抗氧化指标的影响
由表6可知,与CON组相比,100 PSE组和200 PSE组蛋鸡血清中T-SOD活性分别提高13.67%(P>0.05)和50.11%(P<0.05)。与CON组相比,100 PSE组和200 PSE组蛋鸡肝脏中MDA含量均存在下降趋势(P=0.078);200 PSE组蛋鸡肝脏中CAT活性呈上升趋势(P=0.067)。其他血清和肝脏抗氧化指标各组间均无显著差异(P>0.05)。

表6 PSE对产蛋末期蛋鸡血清和肝脏抗氧化指标的影响Table 6 Effects of PSE on serum and liver antioxidant indices of laying hens during late laying period
2.5 PSE对产蛋末期蛋鸡肝脏抗氧化相关基因表达的影响
由图1可知,与CON组相比,200 PSE组肝脏中Kelch样环氧氯丙烷相关蛋白1(Kelch-like epichlorohydrin-associated protein 1,Keap1)mRNA相对表达量显著下调(P<0.05);200PSE组肝脏中核转录因子E2相关因子2(nuclear factor E2-related factor 2,Nrf2)、血红素加氧酶-1(heme oxygenase-1,HO-1)、醌氧化还原酶-l(quinineoxidoreductase-1,NQO-1)mRNA相对表达量较CON组均有所提高,但差异不显著(P>0.05)。
2.6 PSE对产蛋末期蛋鸡血清和肝脏卵黄前体物质VLDL、VTG含量的影响
由表7可知,与CON组相比,100 PSE组和200 PSE组血清中VLDL含量分别提高4.51%和2.02%,但未达到统计学差异(P>0.05);200 PSE组血清中VTG含量提高25.03%,差异达显著水平(P<0.05)。与CON组相比,100 PSE组和200 PSE组肝脏中VLDL含量分别提高28.42%和37.66%,差异均达到极显著水平(P<0.01),肝脏中VTG含量分别提高21.64%和32.43%,差异均达到显著水平(P<0.05)。

表7 PSE对产蛋末期蛋鸡血清和肝脏卵黄前体物VLDL、VTG含量的影响Table 7 Effects of PSE on yolk precursors VLDL and VTG contents in serum and liver of laying hens during late laying period
2.7 PSE对产蛋末期蛋鸡肝脏卵黄前体物质合成相关基因表达的影响
由图2可知,与CON组相比,100PSE组和200 PSE组肝脏中卵黄蛋白原Ⅱ(vitellogenin Ⅱ,VTGⅡ) mRNA相对表达量显著上调(P<0.05);200 PSE组肝脏中雌激素受体-α(estrogen receptor-α,ER-α)mRNA相对表达量极显著高于CON组和100 PSE组(P<0.01),同时极低密度载脂蛋白Ⅱ(very low-density apoli-poprotein Ⅱ,ApoVLDLⅡ)mRNA相对表达量显著高于CON组(P<0.05);100 PSE组和200 PSE组肝脏中载脂蛋白B(apolipoprotein B,ApoB)mRNA相对表达量均较CON组有所提高,但差异不显著(P>0.05)。肝脏中雌激素受体-β(estrogen receptor-β,ER-β)mRNA相对表达量各组间均无显著差异(P>0.05)。

图2 PSE对产蛋末期蛋鸡肝脏卵黄前体物质合成相关基因表达的影响Fig.2 Effects of PSE on expression of genes related to yolk precursor synthesis in liver of laying hens during late laying period
3 讨 论
3.1 PSE对产蛋末期蛋鸡产蛋性能和蛋品质的影响
产蛋性能是衡量蛋禽产蛋状况和养殖经济效益的重要指标。处于产蛋后期的蛋鸡产蛋性能下降,表现为产蛋率降低、料蛋比和日耗料量增加[11]。PS在水中与脂质结合成植物激素-植物甾醇-核糖核蛋白复合体后,增加了其在动物体内分解的稳定性,从而诱导蛋白质合成,调节动物健康和生长[8]。PS有提高畜禽生产性能的趋势,PSE相较PS具有更强的脂溶性且更易被肠道吸收利用,两者生理功能相同。王龙昌等[12]在520日龄产蛋后期蛋鸡饲粮中分别添加5、10、20和40 mg/kg PS,发现饲喂5 mg/kg PS的蛋鸡全期产蛋率和平均蛋重分别提高1.05%和3.13%,且试验组死淘率均降低。本团队前期研究结果显示,110日龄日本蛋鹌鹑饲粮中PSE添加量为150和300 mg/kg时,其产蛋率相较对照组分别显著提高2.68%和4.31%[10]。本试验表明,饲粮中适量添加PSE可提高蛋鸡全期产蛋率和平均蛋重,降低后期料蛋比,改善产蛋末期蛋鸡产蛋性能受损状况。
商品蛋鸡从产蛋高峰期至产蛋后期,其生理代谢活动随周龄发生变化,导致蛋鸡产蛋性能降低,蛋品质下降[13]。鸡蛋重量随蛋鸡年龄的增长而增加,但蛋壳重量不增加,这种不成比例的变化可能导致蛋壳质量下降[14]。蛋黄颜色评分与蛋黄中类胡萝卜素含量呈线性相关[15],含类胡萝卜素的乳糜微粒经肠道吸收后转运至肝脏,在肝脏中与VLDL结合并向外周组织输送[16]。Xiao等[14]研究发现,经过5周PS(20和40 mg/kg)处理后,老龄蛋鸡的鸡蛋外观质量(蛋重、蛋壳表面积)和内部品质(蛋白高度、哈氏单位和蛋黄颜色)均呈线性提高;在试验第7周,PS线性提高蛋重和蛋壳重量。常玲玲等[17]在海兰褐蛋鸡基础饲粮中分别添加20、80、400、800 mg/kg PS后,发现PS对蛋形指数、蛋白高度和哈氏单位等蛋品质指标均无显著影响。本试验结果与上述研究结果相似,与CON组相比,饲粮添加PSE显著提高产蛋末期蛋鸡蛋重、蛋壳重量和蛋壳表面积,改善鸡蛋的外观质量。此外,PSE一定程度上改善了蛋黄颜色,可能是因为PSE增加禽类肝脏中VLDL的含量,从而促进类胡萝卜素在蛋黄中的沉积。
3.2 PSE对产蛋末期蛋鸡血清生殖激素指标的影响
下丘脑-垂体-性腺轴控制蛋鸡的生殖系统,直接调控卵泡发育、成熟和排卵,最终调节产蛋量[18],其中促性腺激素(如FSH、LH)对调节卵巢发育、类固醇激素(如E2、Prog)的产生和配子发生至关重要。FSH能刺激卵泡发育成熟,LH可诱导排卵前卵泡的生长发育并触发排卵,促进黄体形成并分泌Prog,两者协同作用下刺激卵泡成熟、排卵并提高E2的分泌量[12]。因此,血清生殖激素水平可作为指示产蛋性能的指标。老龄蛋鸡产蛋量的下降与FSH和LH分泌减少有关[19]。PS可作为类固醇激素从头生物合成的前体,在体内能表现出一定的激素活性,例如,谷甾醇和豆甾醇可能是植物中Prog的甾醇前体[8,20]。研究表明,PS对雌鼬血浆中E2和LH水平有不同程度的提高作用[21];成熟雌性湖鳟接触10或20 μg/L的PS后,其血浆中E2水平增加并刺激肝脏中VTG的产生,说明雌性排卵期延长[22]。王龙昌等[12]饲喂520日龄蛋鸡5或40 mg/kg PS后,其血清中E2水平显著提高,FSH水平未发生显著变化,与本试验结果存在差异,可能是不同种属和不同时期的动物在应答时存在不同的机制和结果。在本研究中,饲粮添加PSE显著提高老龄蛋鸡血清中FSH、LH、E2和Prog水平,从而增加卵泡数量、刺激卵泡生长成熟,这可由2个添加PSE组产蛋率提高得到证实。
3.3 PSE对产蛋末期蛋鸡肝脏抗氧化能力的影响
蛋鸡在衰老过程中,机体内活性氧(ROS)不断积聚,内源性抗氧化剂不足以中和过量的自由基和过氧化物,ROS生成和清除的平衡被打破、抗氧化防御系统失衡,产生氧化应激,致使卵泡衰竭、繁殖性能下降,从而导致老龄蛋鸡产蛋量降低[3]。在抗氧化防御系统中,超氧化物歧化酶(superoxide dismutase,SOD)可作为解毒超氧阴离子自由基的第一线,将氧自由基还原为过氧化氢(H2O2),再经CAT将H2O2转化为水(H2O)和氧气(O2),此过程中GSH-Px可与SOD协同作用,将其转化为无毒的羟基化合物,防止氧自由基堆积并清除脂质过氧化物终产物MDA的形成[13,23]。PS作为温和的自由基清除剂,其结构中的C-24位叔碳具有较强的抗氧化能力,可与ROS反应生成稳定的叔自由基,从而阻断脂肪酸链的氧化[8]。田丹丹等[24]研究证实,PS对羟自由基、DPPH自由基和超氧阴离子均有较强的清除和抑制作用,且清除率与其浓度呈正相关性。
黄志毅等[25]在清远麻鸡饲粮中分别添加100、200和300 g/t PS,提高了血清中SOD和GSH-Px活性并降低了MDA含量,增强了麻鸡的抗氧化能力。李志华等[26]发现,在罗非鱼饲料中添加20或40 mg/kg的PS后,肝脏中SOD和CAT活性均较对照组有所提高,但当添加量为160 mg/kg时两者活性较对照组有所降低。由此可推断出,PS与抗氧化酶活性存在剂量关系,适宜的添加量对抗氧化能力存在积极作用。卿一青[10]报道,饲粮中添加150~300 mg/kg PSE可显著提高蛋鹌鹑血清中抗氧化酶(T-SOD和CAT)活性,增强肝脏中GSH-Px活性。本试验结果与上述研究结果相似,饲粮添加PSE后蛋鸡血清中T-SOD和肝脏中CAT活性升高,肝脏中MDA含量下降,且200 mg/kg的效果优于100 mg/kg。
机体正常情况下,特异性受体Keap1可通过泛素-蛋白酶体途径与抗氧化应激的关键转录因子Nrf2结合,抑制其功能[27]。氧化应激导致Keap1的构象发生改变,Nrf2解离并释放,活化的Nrf2可反式定位到细胞核与抗氧化反应元件(antioxidant response element,ARE)结合,启动下游抗氧化酶基因[包括谷胱甘肽硫转移酶(glutathione S-transferase,GST)、NQO-1、HO-1等]的表达,从而缓解H2O2诱导的氧化损伤[3,27]。Gao等[28]研究表明,蛋鸡饲粮中添加染料木黄酮(20和40 mg/kg)能够激活卵巢中ER-α的表达,调节ER-α/Nrf2-Keap1抗氧化信号通路,协同提高抗氧化酶(CAT、T-SOD和GSH-Px)的活性,抑制MDA的过度积累,从而提高卵巢的抗氧化能力;Yu等[29]报道了Nrf2/HO-1通路的调控对于保护胎盘免受氧化应激至关重要。此外,研究发现金针菇茎具有抗氧化性,可通过调节Keap1-Nrf2/ARE信号通路相关蛋白的表达,增加血清、肝脏和卵巢中抗氧化酶(GSH-Px和T-SOD)活性,降低MDA含量[30]。在本研究中,与CON组相比,200 PSE组肝脏中Keap1 mRNA相对表达量显著下调,其Nrf2、HO-1和NQO-1 mRNA相对表达量较CON组均有所提高。因此,PSE能够下调Keap1 mRNA的表达,促进Nrf2的活化和释放,进一步启动ARE调控的下游抗氧化酶基因的表达。相应地,本试验发现,饲粮中添加200 mg/kg PSE后显著提高血清中T-SOD活性,同时增强肝脏中CAT活性并降低MDA含量。本试验所得部分试验结果与前人研究结果存在出入,结果的差异可能是由于动物品种、饲养阶段、PSE吸收效率和添加量不同等造成的。例如,年龄增长可引起机体ROS和蛋白质羰基水平增加,从而导致抗氧化剂的需求量更高[31]。综合上述指标,推测PSE可通过激活Keap1-Nrf2/ARE信号通路提高肝脏抗氧化能力,从而减轻产蛋末期蛋鸡肝脏氧化损伤。
3.4 PSE对产蛋末期蛋鸡卵黄前体物质合成的影响
肝脏作为氧化应激的靶向和效应器官,其氧化还原状态与卵黄前体物质的合成密切相关。蛋鸡在衰老过程中易发生氧化应激,导致肝脏生理功能衰退和抗氧化能力减弱、卵黄前体物合成受阻,进而影响产蛋性能[5]。VLDL和VTG是运输和沉积额外营养物质进入卵巢,满足卵母细胞生长需要的关键卵黄前体物质[32]。产蛋鸡体内的VLDL分为普通型VLDL和小颗粒VLDL(VLDLy),产蛋开始后E2将几乎所有的普通型VLDL产生的肝细胞脂蛋白转化为VLDLy(卵巢靶向),VLDLy是唯一能成功到达发育中的卵母细胞沉积成卵黄的物质[32-33]。VLDLy运输需2种载脂蛋白,即载脂蛋白B(ApoB)和ApoVLDLⅡ。氧化应激会使肝脏发生脂质过氧化和脂肪变性,导致ApoB降解及错误折叠,VLDLy合成受阻[34-35]。VTG是一种磷脂糖蛋白,既可作为非极性分子或酶的载体结合金属离子(如钙、锌和铁)并运输至卵母细胞,又负责供应胚胎氨基酸、葡萄糖、脂肪和磷等营养性物质[2,36]。此外,ApoVLDLⅡ和VTG的表达量均与蛋鸡的产蛋量有关[37]。
血清VLDL含量能够反映蛋鸡的产蛋能力,VLDL含量低时其产蛋率也相应降低[38]。Gao等[39]在50周龄蛋鸡饲粮中添加天然虾青素(ASTA),发现可显著提高肝脏和卵巢中T-AOC、T-SOD和GSH-Px活性,降低MDA含量,且随着ASTA添加量(50~100 mg/kg)的增加,血浆中VLDL-C含量逐渐升高,但在肝脏中VLDL-C含量无显著变化。Ali[6]用含0.5%甜菜碱的饲粮连续4周饲喂38周龄蛋鸡,结果表明,蛋鸡产蛋率和血清中VTGⅡ含量显著提高。此外,卢建军等[40]在老龄蛋鸡饲粮中添加600 mg/kg甜菜碱后,发现血清中VLDL和VTG含量显著升高,且产蛋率由76.50%提升至83.13%,料蛋比下降9.02%。本试验结果显示,饲粮中适量添加PSE一定程度上提高了老龄蛋鸡血清中VLDL含量,并且血清中VTG含量和肝脏中VLDL和VTG含量均显著提高,其中添加量以200 mg/kg为佳,其全期产蛋率也由75.33%提升至81.82%,这表明PSE可能通过增加卵黄前体物质的含量,从而改善产蛋末期蛋鸡的产蛋性能。
蛋鸡排卵时,卵巢产生的E2经血液循环至肝脏,介导肝脏上的雌激素受体(包括ER-α和ER-β,以ER-α作用为主),调控VTGⅡ、ApoB和ApoVLDLⅡ mRNA的表达,进而促进卵黄前体物质的组装合成[41]。Ali[6]进一步探究了甜菜碱对体外鸡胚原代肝细胞的影响,验证其可通过糖皮质激素受体与ER-α间的互作调控肝脏中卵黄前体物质合成相关基因(VTGⅡ和ApoVLDLⅡ)的表达,促进卵黄前体物质的合成。Amevor等[42]将槲皮素(0.4 g/kg)添加于种鸡饲粮中,发现在种鸡激素及其受体表达水平较低的情况下,槲皮素可作为植物雌激素提高E2、LH、FSH及相关受体的水平,促进卵泡发育成熟,进而显著上调VTGⅡ、ApoB和ApoVLDLⅡ等基因的表达。PS具有影响性腺合成类固醇激素的能力,并且在动物体内发挥一定的类雌激素功能。研究发现,β-谷甾醇发挥着植物雌激素的作用,且倾向与ER-β结合[43]。Tremblay等[44-45]证明β-谷甾醇可与雌激素受体结合并促进虹鳟鱼肝细胞中VTG的产生而表现出E2活性,进一步研究发现β-谷甾醇可增加血浆中VTG含量并诱导VTGmRNA表达。本试验结果表明,饲粮中添加200 mg/kg PSE可显著提高产蛋末期蛋鸡血清中E2水平和肝脏中ER-αmRNA相对表达量,上调肝脏中VTGⅡ和ApoVLDLⅡ的表达,这与Amevor等[42]的研究结果相似,表明PSE可发挥一定的类雌激素作用,诱导卵黄前体物质合成相关基因ER-α、VTGⅡ和ApoVLDLⅡ的表达。VLDL和VTG含量与产蛋性能也并非简单的相关关系,是否存在与其他调控因子(如脂质代谢)的共同作用还需进一步研究。
4 结 论
饲粮中添加适量PSE可提高产蛋末期蛋鸡血清生殖激素(FSH、LH、E2和Prog)水平和抗氧化酶(血清中T-SOD、肝脏中CAT)活性,降低肝脏中MDA含量和Keap1 mRNA相对表达量,增强肝脏的抗氧化能力,同时诱导肝脏中卵黄前体物质合成相关基因ER-α、VTGⅡ和ApoVLDLⅡ的表达,促进肝脏组装合成卵黄前体物质VTG和VLDL,从而改善产蛋末期蛋鸡因氧化损伤而导致的产蛋性能和蛋品质下降状况,且本试验中以200 mg/kg效果较好。
