降香黄檀根瘤以及根瘤菌形态和超微结构特征
2023-12-04邓家珍叶绍明林铭业蓝雅惠樊容源潘彩玲
邓家珍,叶绍明,林铭业,蓝雅惠,燕 羽,樊容源,潘彩玲
(广西大学林学院,广西森林生态与保育重点实验室,广西 南宁 530004)
豆科植物的根与根瘤菌(rhizobia)共生是常见的一种生物固氮(biological nitrogen fixation,BNF)方式,在环境保护、全球氮循环和农林生产等方面发挥着重要作用[1]。根瘤菌能够将大气中的氮转化为豆科植物可利用的氨,而豆科植物则为根瘤菌提供生存所需的营养物质[2]。豆科植物与根瘤菌形成的互惠共生关系导致了根瘤(root nodule)的形成。根瘤是重要的共生固氮场所,不仅能促进宿主和共生体之间的代谢物交换,还能将固氮产物输送给植物的地上部。根瘤的特殊结构和生理环境,不但保护了固氮酶而且提供了合适的固氮条件[3-4]。目前,关于豆科植物根瘤的结构已有大量研究,尤其关注根瘤的形成发育过程和固氮生理[5-7]。已有学者从超微水平研究豆科植物根瘤的结构,对根瘤侵染细胞的特征、侵入线的结构以及多磷酸盐积累的特征等展开研究[8-10],但多集中于草本豆科植物,如豌豆(Pisumsativum)、田菁(Sesbaniacannabina)、大豆(Glycinemax)等[11-13],关于木本豆科植物研究较少,特别是对木本豆科植物根瘤形态及超微结构特征的研究尚鲜见报道,研究木本豆科植物根瘤的形态及超微结构特征对揭示豆科植物与根瘤菌共生关系具有重要意义。
降香黄檀(Dalbergiaodorifera)是蝶形花科黄檀属(Dalbergia)常绿乔木,国家二级保护植物,其木材珍贵且经济价值高[14]。降香黄檀根系发达能形成根瘤,可起到改良土壤、减少氮肥使用的作用,具有良好的生态效益。目前已有学者对降香黄檀的心边材成分、药理作用、叶的活性成分、栽培技术和资源分布调查等[15-18]进行了研究,但鲜有关注降香黄檀与根瘤菌的共生固氮关系,尤其是降香黄檀用来进行固氮作用的器官——根瘤。仅发现谭小明等[19]对降香黄檀根瘤进行过解剖学研究,而关于其超微结构方面目前暂鲜见相关报道。鉴于此,本研究以降香黄檀根瘤为材料,制作石蜡切片、超薄切片和扫描电镜样品,利用光学显微镜(optical microscope,OM)、透射电镜(transmission electron microscope,TEM)和场发射扫描电镜(field emission scanning electron microscope,FESEM)研究降香黄檀根瘤和瘤内根瘤菌的形态及其超微结构,以期在细胞学水平上更深层次厘清降香黄檀根瘤及根瘤菌的特征,为探究降香黄檀与根瘤菌的共生固氮机制提供理论依据。
1 材料与方法
1.1 试验地概况和根瘤样品采集
采样地位于广西大学林学院苗圃实习基地(108°17′14″E,22°51′20″N),于2021年1月在基地塑料大棚内种植降香黄檀。供试苗木为1年生降香黄檀(幼苗无根瘤),每盆种植2株,盆口径40 cm、高45 cm。供试土壤取自良凤江国家公园,清除石粒及杂质后混入珍珠岩以保持透水性,泥土与珍珠岩的体积比为25∶1,经过灭菌处理后待用。待植株服盆(14 d)后,每15 d给幼苗根部浇灌根瘤菌菌液100 mL直至结瘤,根瘤菌菌株为费氏中华根瘤菌(Sinorhizobiumfredii),购自广东省微生物菌种保存中心。采用酵母甘露醇琼脂固体培养基(yeast mannitol agar,YMA)培养活化保存的根瘤菌菌株,菌株活化后,接入YMA液体培养基中,28 ℃下摇床培养5~6 d,用分光光度计检测菌液吸光度OD600(OD600表示根瘤菌培养液在600 nm处的光吸收值)值达0.7时即可浇灌。该接菌结瘤试验设置6个重复,每个重复设置10盆,共60盆,120株苗。接菌后每隔5 d在每个重复中随机选取1盆观察幼苗根系结瘤情况。本研究以肉眼观察或借用放大镜辅助观察能见根瘤的当天作为计算瘤龄的第0天,之后在第1、3、5、7、10、20、60天时采集根瘤。每次采样于6个重复内随机选取4个重复,每个重复采集1盆,共收获4盆,8株幼苗。
采集根瘤时,将植株基部的枯枝落叶和0~5 cm的表层土清除,然后收集土层中降香黄檀幼苗根系的全部根瘤,并放入装有冰袋的泡沫盒中带回实验室。将样品用蒸馏水冲洗干净,吸走表面水分后,使用放大镜观察降香黄檀根瘤的外部形态特征,记录根瘤形态、颜色等指标。使用游标卡尺(精度0.01 mm)测量根瘤的长径。将取下的根瘤按照50%、25%、25%的数量比例分别用于石蜡切片的制备以及扫描电镜样品的制备、超薄显微切片的制备、豆血红蛋白含量测定。
1.2 降香黄檀石蜡切片、超薄切片及扫描电镜样品制作
石蜡切片制样[20]。根瘤于酒精醋酸福尔马林混合固定液(FAA)中固定24 h,采用乙醇梯度浓度(体积分数)30%(2 h)、50%(2 h)、70%(2 h)、90%(2 h)、100%(1 h,重复1次)进行脱水,利用TO型生物制片透明剂进行透明,浸蜡包埋(熔点58~60 ℃),切片机(LEICA SM 2010 R)切片(厚度为8 μm),展片,将完全晾干的载玻片浸入TO透明剂中脱蜡30 min,重复2次。之后每隔30 min分别浸入1/3 100%乙醇+2/3 TO透明剂、1/2 100%乙醇+1/2 TO透明剂、2/3 100%乙醇+1/3 TO透明剂、100%乙醇、90%乙醇、70%乙醇、50%乙醇、纯水浸泡,进行样品组织的复水。采用番红-固绿染色法进行染色,中性树胶封片,在光学显微镜(LEICA ICC50 W)下观察降香黄檀根瘤组织结构。
超薄切片制样参照Wang等[21]的方法并做相应改良:将根瘤切成大小1 mm(长)×1 mm(宽)×3 mm(高),浸入2.5%(体积分数)的戊二醛溶液4 ℃固定过夜后用pH 7.0的磷酸缓冲液漂洗样品,1%(体积分数)锇酸溶液固定样品,取出锇酸废液后用pH 7.0的磷酸缓冲液漂洗,乙醇梯度脱水,纯丙酮处理20 min,再用包埋剂与丙酮的混合液(体积比为1∶1)处理样品1 h,后用包埋剂与丙酮的混合液(体积比为3∶1)处理样品3 h,将经过渗透处理的样品包埋,70 ℃加热过夜,即得到包埋好的样品。样品在超薄切片机(LEICA EM UC 7)中切片,厚度为70~90 nm,切片经柠檬酸铅染液和醋酸双氧轴染液各染色5 min后晾干即可上透射电镜(JEM 1200 EX)观察。
扫描电镜样品制备[22]。根瘤在FAA固定液中固定24 h后取出,用pH为7.0的磷酸缓冲液每隔15 min冲洗1次,共冲洗3次。将冲洗过的样品每隔10 min依次浸入50%、70%、80%、90%、100%(体积分数)的乙醇中进行脱水。将脱水后的样品自然风干。将风干后的样品分别粘台、编号,离子溅射仪(Cressington sputter coater 408,英国)喷金,放置于场发射扫描电镜(FEI Quattros)载物台后观察、拍照。每个样品的拍摄视角不少于15个。
1.3 降香黄檀不同时期根瘤豆血红蛋白含量测定
利用牛血红蛋白作为标准品,称取1 mg粉末溶于0.1 mol/L磷酸缓冲液(PBS,pH 6.8)中配置成1 mol/L标准母液。使用移液枪分别吸取1、2、3、4、5和6 mL母液至10 mL容量瓶中后用上述磷酸缓冲液定容,获得浓度0.1、0.2、0.3、0.4、0.5和0.6 mol/L的溶液。使用紫外分光光度计(Ultrospec 2100 pro)在540 nm处测定吸光值,绘制以牛血红蛋白浓度为横坐标,以OD540值为纵坐标的标准曲线。根瘤冻存于-80 ℃超低温冰箱,分别取出瘤龄为1、3~5、7~10、11~20、21~60 d的根瘤于液氮中研磨成粉末,准确称取0.2 g粉末样品,加入8 mL的低温(4 ℃)PBS。悬浊液在4 ℃、1 000 r/min下离心15 min,吸取上清液继续在4 ℃、12 000 r/min条件下离心20 min。以磷酸缓冲液为空白对照,在540 nm处测定上清液吸光值。利用标准曲线计算豆血红蛋白含量[23]。
1.4 数据统计及分析
数据采用平均值±标准误差表示,使用Excel 2010进行数据整理。采用单因素方差分析法(One-way ANOVA)对豆血红蛋白含量数据进行显著性差异分析,显著性水平设置为α=0.05。其中,单因素方差分析在R 4.1.1中调用“agricolae”和“tidyverse”包完成[24]。
2 结果与分析
2.1 降香黄檀不同时期的根瘤形态及豆血红蛋白含量变化
连续观察降香黄檀根部的变化发现,幼苗浇菌后的第20~25天开始显瘤,随发育进行,根瘤的形状由圆形逐渐分化为椭圆形、杠铃形及不规则椭圆形等,颜色由黄白色向黄褐色转变,着生部位由侧根和基部逐渐向主根扩散(图1、表1)。
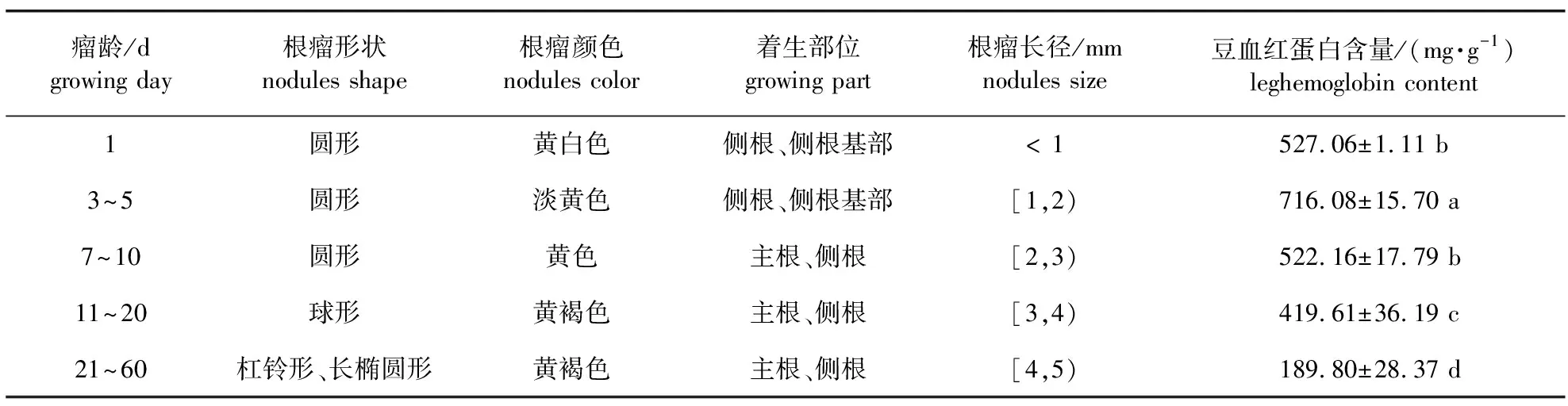
表1 不同时期降香黄檀根瘤形态及豆血红蛋白含量

a—g.分别为瘤龄1、3、5、7、10、20、60 d时的根瘤形态。These are morphology of root nodules of 1,3,5,7,10,20 and 60 d respectively. h.根瘤解剖the anatomy of root nodules。 图1 不同时期的降香黄檀根瘤形态Fig. 1 The root nodules morphology of Dalbergia odorifera in different stages
随着根瘤变大,其中心部分由鲜明的橙红色逐渐变淡直至灰白且伴随凹陷(图1h)。此外,豆血红蛋白(leghemoglobin)含量呈先升后降的趋势,并在瘤龄为3~5 d的根瘤(长径为1~2 mm)中达到最高(表1)。
2.2 降香黄檀根瘤组织学特点
通过连续石蜡切片显微观察发现(图2),降香黄檀根瘤从外到内由周皮、皮层、内皮层、维管束和侵染组织构成。侵染区域几乎由侵染细胞填满,不同侵染细胞区域间隔2至多层薄壁细胞。侵染细胞周围薄壁细胞的细胞核较皮层细胞的细胞核发达,且能明显看到胞内物质增多(图2a)。在皮层与内皮层之间分布着多个维管束,其切片面积、形状和排列方向各不相同。维管束内有环纹导管以端壁首尾相连,组成连通的长管道,外有1层厚壁细胞环绕(经番红-固绿染色后细胞壁颜色加深)。维管束内的细胞体积较周围薄壁细胞小,但细胞核明显(图2b)。
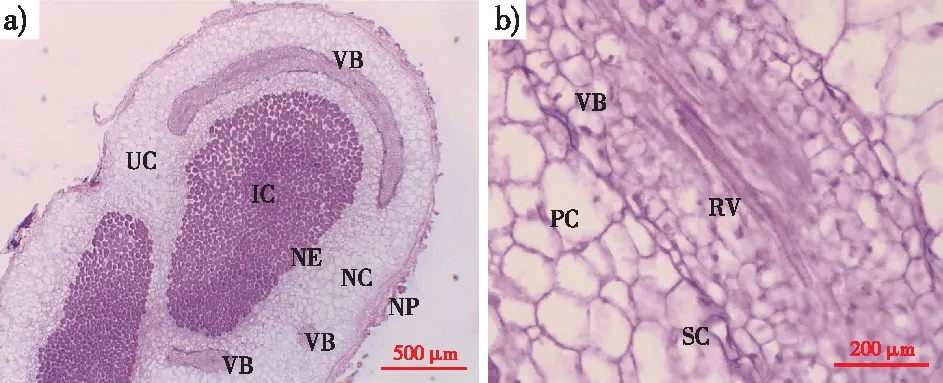
IC.侵染细胞 infected cell;NC.根瘤皮层 nodule cortex;NE.根瘤内皮层 nodule endodermis;NP.根瘤周皮 nodule periderm;PC.薄壁细胞 parenchymatous cell;RV.环纹导管 ringed vessel;SC.厚壁细胞 sclerenchymatous cell;UC.非侵染细胞 un-infected cell;VB.维管束 vascular bundle。图2 降香黄檀根瘤组织结构Fig.2 The histological structure of D. odorifera root nodules
2.3 降香黄檀根瘤的超微结构
观察发现,降香黄檀根瘤的侵染细胞与非侵染细胞在组成上存在明显差别(图3a),侵染细胞区域存在少量非侵染细胞(图3b)。非侵染细胞高度液泡化,细胞器被挤压至细胞壁周围,细胞壁厚薄不均,淀粉粒和一些质体围绕细胞核存在。液泡内含有少量游离的纤维状或颗粒状物质(图3c、3d黑色箭头所示),可能是被液泡里的水解酶降解形成。两个邻近非侵染细胞的细胞壁存在胞间连丝(图3 c白色箭头所示)。侵染细胞内无中央液泡,细胞被形状不一的泡囊填满且泡囊都拥有膜结构,膜内包被着1至多个根瘤菌类菌体。泡囊膜的生成活动剧烈,数量不等的小泡贴附于泡囊膜外侧(图3e、3l)。在部分侵染细胞内,泡囊膜处于分离破碎的状态,根瘤菌被释放至细胞质中成为独立的个体,这可能预示细胞的凋亡(图3f、3k)。侵染细胞的细胞核与细胞膜系统组装活动有关,可明显看到侵染细胞的核仁、核膜及染色质(图3g);有的侵染细胞的细胞核则显示出核凸起,可能与核内多轮DNA复制相关[25](图3h)。侵染细胞内的线粒体、内质网等细胞器由于根瘤菌数量的增加被挤到细胞壁边缘(图3i、3j、3k)。
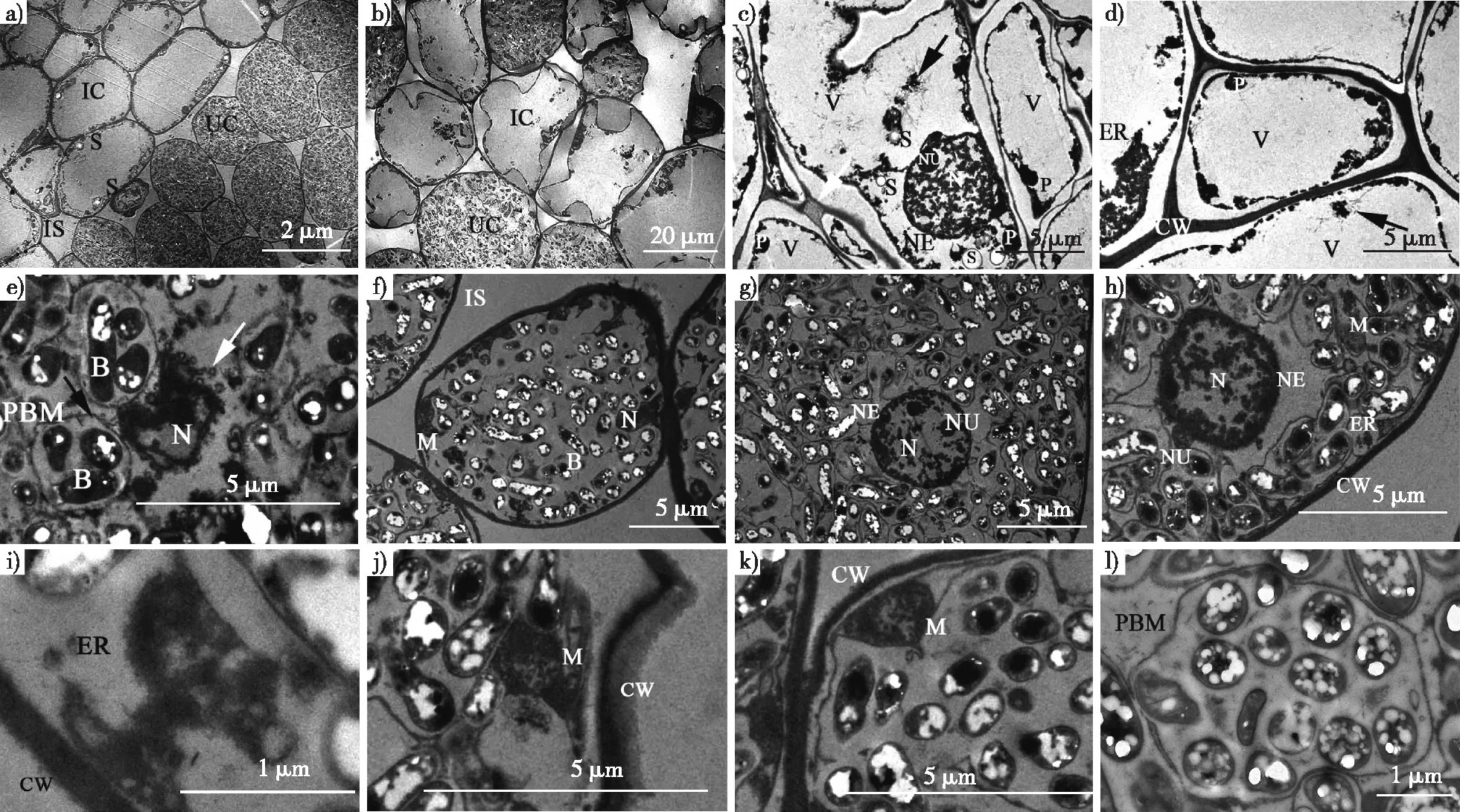
B.类菌体 bacteroid;CW.细胞壁cell wall;ER.内质网 endoplasmic reticulum;IC.侵染细胞 infected cell;IS.细胞间隙 intercellular space;M.线粒体 mitochondria;N.细胞核 nucleus;NE.核膜 nucleus envelope;NU.核仁 nucleolus;P.质体 plastid;PBM.泡囊膜 peribacteroid membrane;S.淀粉粒 starch grain;UC.非侵染细胞 un-infected cell;V.液泡 vacuole。图3 降香黄檀根瘤超微结构Fig. 3 Ultrastructure of D. odorifera root nodules
2.4 降香黄檀根瘤内根瘤菌的超微观察
通过透射电镜观察可见,降香黄檀根瘤细胞内类菌体附近出现一些絮状的膜物质(图4a箭头所示),可能是由膜小泡聚集融合形成。发现泡囊膜解体现象,由于膜的破裂,膜碎片及一些颗粒物质存在于细胞质中(图4b)。根瘤菌侵入根瘤细胞后会分化成类菌体,降香黄檀根瘤侵染细胞内含大量长短不一,粗细不均的根瘤菌类菌体,其形状多样,有杆形、梨形、“Y”形、哑铃形等(图4d、4e、4f),并具有细胞壁和细胞膜(图4c)。类菌体不具备完整的细胞核,而是拥有一个相当于细胞核的拟核区,在拟核区的附近存在明显的黑色圆形小颗粒(图4b、4c、4d箭头所示),此黑色颗粒为多磷酸盐(polyphosphate,PP)颗粒。除了多磷酸盐颗粒外,根瘤菌类菌体内还存在大量电子密度大的白色斑点,为聚-β-羟基丁酸盐(poly-β-hydroxybutyrate,PHB)颗粒。

B.类菌体 bacteroid;CW.细胞壁 cell wall;PBM.泡囊膜 peribacteroid membrane;PP.多磷酸盐颗粒 polyphosphate particle;PHB.聚-β-羟基丁酸盐颗粒 poly-β-hydroxybutyrate。图4 降香黄檀根瘤内根瘤菌的超微观察Fig. 4 Ultraobservation of rhizobium in D. odorifera root nodules
2.5 降香黄檀根瘤扫描电镜超微观察
与透射电镜相比,场发射扫描电镜呈现的图像为立体三维图,能更直观和清晰地了解降香黄檀根瘤内外样貌。从观察到的视野来看,降香黄檀根瘤表面凹凸不平,褶皱多,周皮呈破碎状(图5a)。非侵染区域和侵染区域分区明显,非侵染细胞与侵染细胞最大的区别是前者不含根瘤菌。非侵染细胞内的物质呈现颗粒或者团状,分布不均,可见圆球状的淀粉粒(图5b、5c);侵染细胞饱满膨大,内含大量长杆状根瘤菌类菌体交织缠绕,细胞壁与胞间隙明显。多个泡囊膜贴合在一起呈现连续状(图5c箭头所示),把类菌体包围堆叠起来形成紧密的膜系统(图5d)。扫描电镜观察到的类菌体形态多为长杆状,少见圆球形的类菌体。每个侵染细胞所含的类菌体数量不一,刚被侵染的细胞根瘤菌数量稀少,随着根瘤菌的分裂,类菌体的数量不断增加直至充满整个细胞(图5e、5f)。

B.类菌体 bacteroid;CW.细胞壁 cell wall;IC.侵染细胞 infected cell;PBM.泡囊膜 peribacteroid membrane;RN.根瘤root nodule;UC.非侵染细胞 un-infected cell。图5 降香黄檀根瘤的扫描电镜观察Fig. 5 Observation of D. odorifera root nodules by scanning electron microscope
3 讨 论
豆科植物的根瘤可分为无限型根瘤(indeterminate nodule)和有限型根瘤(determinate nodule)[26]。无限型根瘤产生根瘤原基皮层为内皮层,其形状通常不规则且保留顶端分生组织;有限型根瘤产生根瘤原基皮层为外皮层,形状通常为圆形,但分生组织失去活性,只能有限生长[27]。此外,无限型根瘤由顶端到底部可分为顶端分生组织区、侵染区、过渡区、固氮区、衰老区和胞内腐生区几个年龄梯度不同的部分[28]。本研究中,降香黄檀根瘤呈现圆形或椭圆形,不似刺槐(Robiniapseudoacacia)、银合欢(Leucaenaleucocephala)等无限型根瘤出现多重分叉、形态多样的现象,说明降香黄檀根瘤形成时,根瘤分生组织的分裂活动便停止,不再向四周分裂,只保持细胞的伸长[29]。且根据石蜡切片观察结果,降香黄檀根瘤的组织结构并不具有无限型根瘤所表现的侵染年龄分区[30],因此降香黄檀根瘤属于有限型根瘤,这与草本豆科植物大豆、落花生(Arachishypogaea)等根瘤类型相同[31]。豆血红蛋白是根瘤高效固氮的衡量特征[23,32],本研究发现不同瘤龄的降香黄檀根瘤内豆血红蛋白含量差异显著,这与Wang等[33]研究豆血红蛋白基因Lb1/2/3在百脉根(Lotuscorniculatus)根瘤发育的不同时期表达情况一致,均呈现先上升后下降的趋势。由此可见,降香黄檀根瘤在不同生长时期的固氮能力存在差异。
降香黄檀根瘤含有2~3个侵染区域,合理的解释为在根瘤发育过程中,根瘤菌为扩大侵染面积或响应根瘤皮层细胞的分裂速度,侵染距离相近的外皮层薄壁细胞,形成新的侵染区域[34-35],进而提高根瘤固氮作用面积,这是降香黄檀与根瘤菌共生固氮的特征。维管束连接着根瘤、根系和植物的其他部分,对根瘤进行代谢具有重要作用[36]。降香黄檀根瘤的侵染区周围分布着维管束,能够给根瘤进行固氮作用提供水分和无机盐支持[37]。依据透射电镜和场发射扫描电镜的观察结果,降香黄檀根瘤非侵染细胞与侵染细胞的组成截然不同,说明根瘤菌的入侵改变了根部细胞原有的组成。值得注意的是,降香黄檀根瘤侵染细胞内的液泡已不复存在,这与前人对沙冬青(Ammopiptanthusmongolicus)、银合欢等根瘤侵染细胞的观察结果不一致[38-39],上述植物的根瘤侵染细胞都含大液泡,这可能暗示不同豆科植物根瘤侵染细胞维持胞内环境平衡作用机理不一[40]。此外,降香黄檀根瘤的非侵染细胞之间存在胞间连丝,这与韩善华[41]观察大豆根瘤非侵染细胞特征研究结果相似,说明降香黄檀根瘤的非侵染细胞之间存在信息交流现象,再次证实了非侵染细胞拥有物质运输的功能。
根瘤菌进入宿主根部的皮层细胞定殖后分化成具有强固氮活性的类菌体。类菌体利用宿主植物提供的苹果酸或琥珀酸进行代谢[42],维持自身生理生化反应和固氮反应供能,并与宿主植物进行物质交换和信号转导[43-44]。降香黄檀根瘤内存在大量的根瘤菌类菌体,说明瘤内的固氮代谢活动强烈,有助于根瘤固定空气中的氮。多磷酸盐(PP)颗粒在微生物体内可以用来合成氨基酸、碱基、糖类、脂肪等生命必需物质[45],聚-β-羟基丁酸(PHB)颗粒是一种存在于微生物细胞中并作为营养和能量储存物质参与细胞代谢的天然产物[46]。降香黄檀根瘤菌类菌体内同时含有PP和PHB颗粒,说明其类菌体的固氮活动拥有良好的储能条件作为支持。但这两种颗粒的含量及分布在类菌体内均不一致,导致差异的原因可能与类菌体的发育相关,有关它们产生的时期和途径还有待深入探讨。
泡囊膜,又称类菌体周膜,可以保护类菌体和寄主植物细胞,保证二者之间能进行正常的物质运输、能量代谢和信息传递[47]。根瘤菌从侵入线释放到宿主细胞后,都以膜包裹的类菌体形式存在,否则将引起宿主细胞的防御反应[48]。本研究中降香黄檀根瘤侵染细胞内拥有多个泡囊,这与豌豆[11]、田菁[12]、肯氏相思(Acaciacunninghamia)[49]的根瘤情况类似。Bergerson等[50]最早在大豆根瘤发现类菌体被泡囊膜包围,何一等[51]在白车轴草(Trifoliumrepens)根瘤内发现相邻细菌周膜融合的现象。本研究发现,降香黄檀根瘤侵染细胞内存在泡囊膜的合成状态,其合成可能是以下过程:由内质网或者质膜合成膜小泡释放至细胞质,而后膜小泡移动至类菌体的附近融合成絮状膜,最后通过融合反应变成泡囊膜,包裹着根瘤菌类菌体。由于泡囊膜的面积大小直接影响侵染细胞与类菌体物质交换的量,所以泡囊膜的合成和解离与固氮作用的强弱有一定联系,这也是降香黄檀与根瘤菌互惠共生关系的重要表现。
