鸡传染性支气管炎病毒感染HD11细胞的转录组分析
2023-11-30常新宇沈嘉忱沈瑞廷赵振华侯晓林
陈 璐,常新宇,沈嘉忱,沈瑞廷,赵振华,侯晓林
(北京农学院动物科学技术学院,北京 102206)
鸡传染性支气管炎(infectious bronchitis,IB)是由IB病毒(infectious bronchitis virus, IBV)引起的一种急性、高度传染性呼吸道疾病,于20世纪30年代在美国首次被发现,它是造成全球家禽养殖业经济损失的最重要原因之一[1-2]。IBV主要引起禽类呼吸道疾病,此外,它也会导致肾炎、产蛋量和产蛋品质下降,甚至死亡[3]。由于基因突变和重组,不同的IBV毒株表现出不同的特性,在致病机制、毒力、年龄取向以及受体特异性方面存在差异,迄今为止,还没有针对IBV感染有效药物报道,接种疫苗也不足以提供完全的保护[4],因此IBV的防控还存在巨大挑战[5]。
研究表明,宿主先天免疫在抵抗IBV中发挥重要作用,是机体保护的第一道防线。IBV进入宿主细胞,免疫系统识别产生抗病毒免疫。IBV天然免疫应答的研究受到越来越多的关注,了解感染早期宿主细胞对抗IBV发挥的作用具有重要研究意义。
本研究使用IBV Beaudette株感染鸡巨噬细胞HD11,进行转录组学检测分析,将差异表达基因按照功能进行分类,分析其参与的信号通路,了解IBV感染早期宿主细胞产生的抗病毒反应和免疫应答,为探究IBV的致病机制提供理论依据,从而为疫苗开发奠定基础。
1 材料与方法
1.1 细胞和病毒
鸡巨噬细胞HD11购自中国科学院细胞库(上海),IBV Beaudette株由四川农业大学提供,现保存于北京农学院基础兽医实验室。
1.2 细胞培养与病毒感染
将对数生长期的HD11细胞铺于6孔板中,当细胞汇合率达到80%左右时,感染100 TCID50的IBV,37 ℃、5%CO2孵育1 h后,弃去病毒液,加入2%维持培养基,2 h后收集细胞。病毒感染组命名为IBV-1、IBV-2、IBV-3,未感染病毒的细胞为对照组,分别命名为NC-1、NC-2、NC-3。
1.3 转录组测序
Trizol法提取细胞总RNA,检测合格后将RNA纯化、片段化并反转录成cDNA,使用Illumina TruseqTM RNA sample prep Kit进行文库构建、Illumina NovaSeq6000上机测序分析[6]。
1.4 差异表达基因和模式识别受体的RT-qPCR验证
为了验证测序结果的准确性,随机选择10个与天然免疫、抗病毒反应相关的差异表达基因和3个模式识别受体基因进行RT-qPCR验证(以β-Actin为内参基因,引物由上海生工生物工程有限公司合成,详见表1)。扩增条件:95 ℃预变性10 min;95 ℃变性15 s,60 ℃退火/延伸1 min,共40个循环;熔解曲线分析:95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s,60 ℃ 15 s。GraphPad Prism 8软件统计分析。将得到的Ct值按照2-△△Ct法计算相对表达量,GraphPad Prism 8软件统计分析。
2 结 果
2.1 差异表达基因筛选
以P<0.05,|log2FC|≥1为标准筛选差异表达基因,共检测到2 016个差异表达基因,其中1 363个基因表达上调,653个基因表达下调。
2.2 GO注释分析
GO注释分析显示,差异表达基因共分为细胞组分(cellular component,CC)、生物学过程(biological process,BP)、分子功能(molecular function,MF)三大类,图1为前20个显著富集的二级条目。在CC分类中占比较大的为细胞成分、细胞器、细胞膜;BP分类中占比较大的为细胞过程、生物调节和代谢过程;MF分类中结合、催化活性和分子功能调节剂数量较多。
2.3 KEGG富集分析
KEGG 信号通路富集分析显示,癌症通路、细胞因子和细胞因子受体之间的相互作用、MAPK信号通路是富集到差异表达基因最多的前3个信号通路(图2)。与免疫相关的MAPK信号通路中,主要有促分裂原活化蛋白激酶激酶激酶5(mitogen-activated protein kinase kinase kinase, MAP3K5)、TNF 受体关联因子6(TNF receptor-associated factor 6, TRAF6)、双调蛋白(amphiregulin, AREG)等,这些结果可以进一步为细胞抗IBV的天然免疫应答的研究提供理论基础。
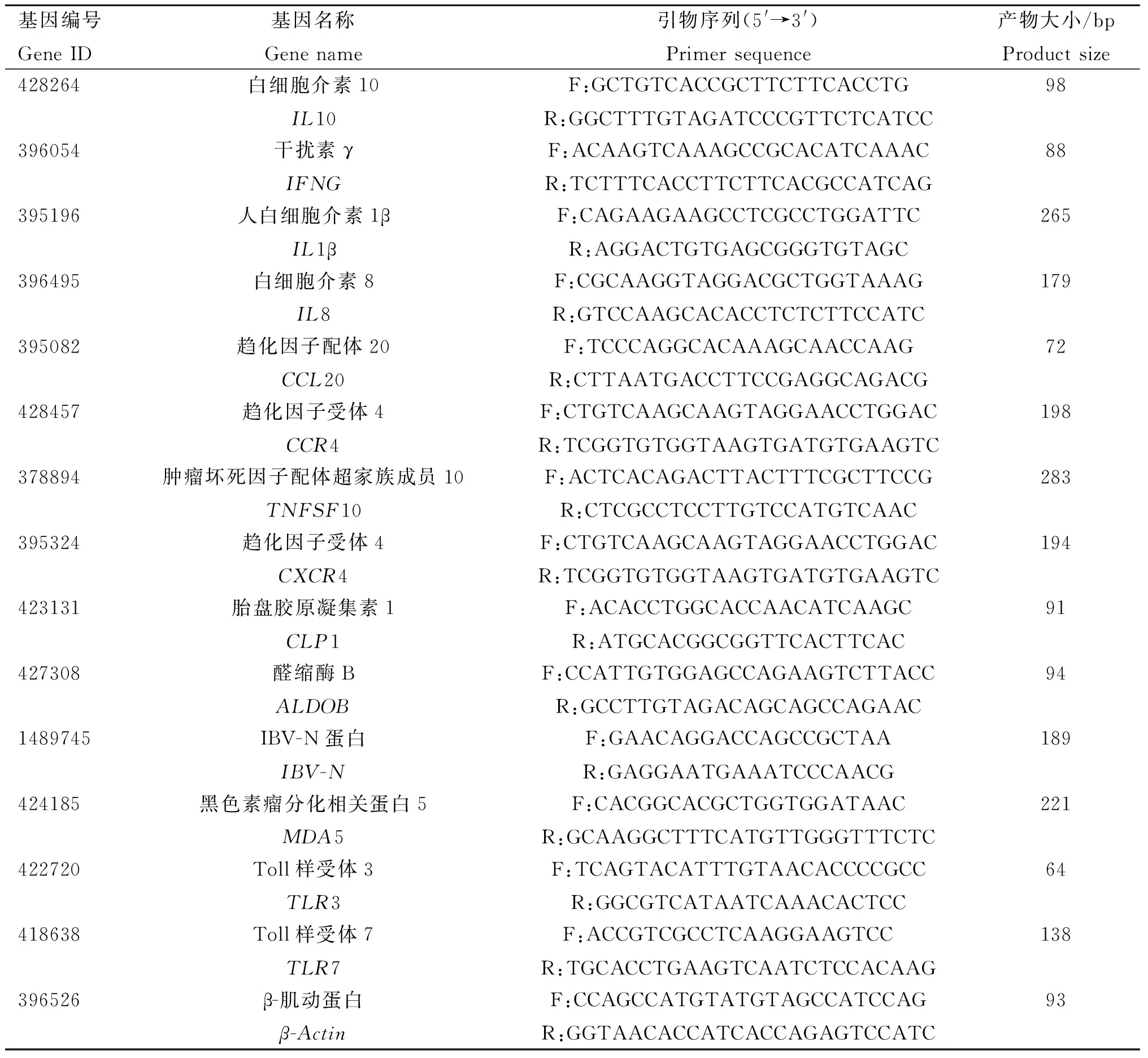
表1 差异表达基因RT-qPCR引物设计Table 1 Design of RT-qPCR primers for differentially expressed genes
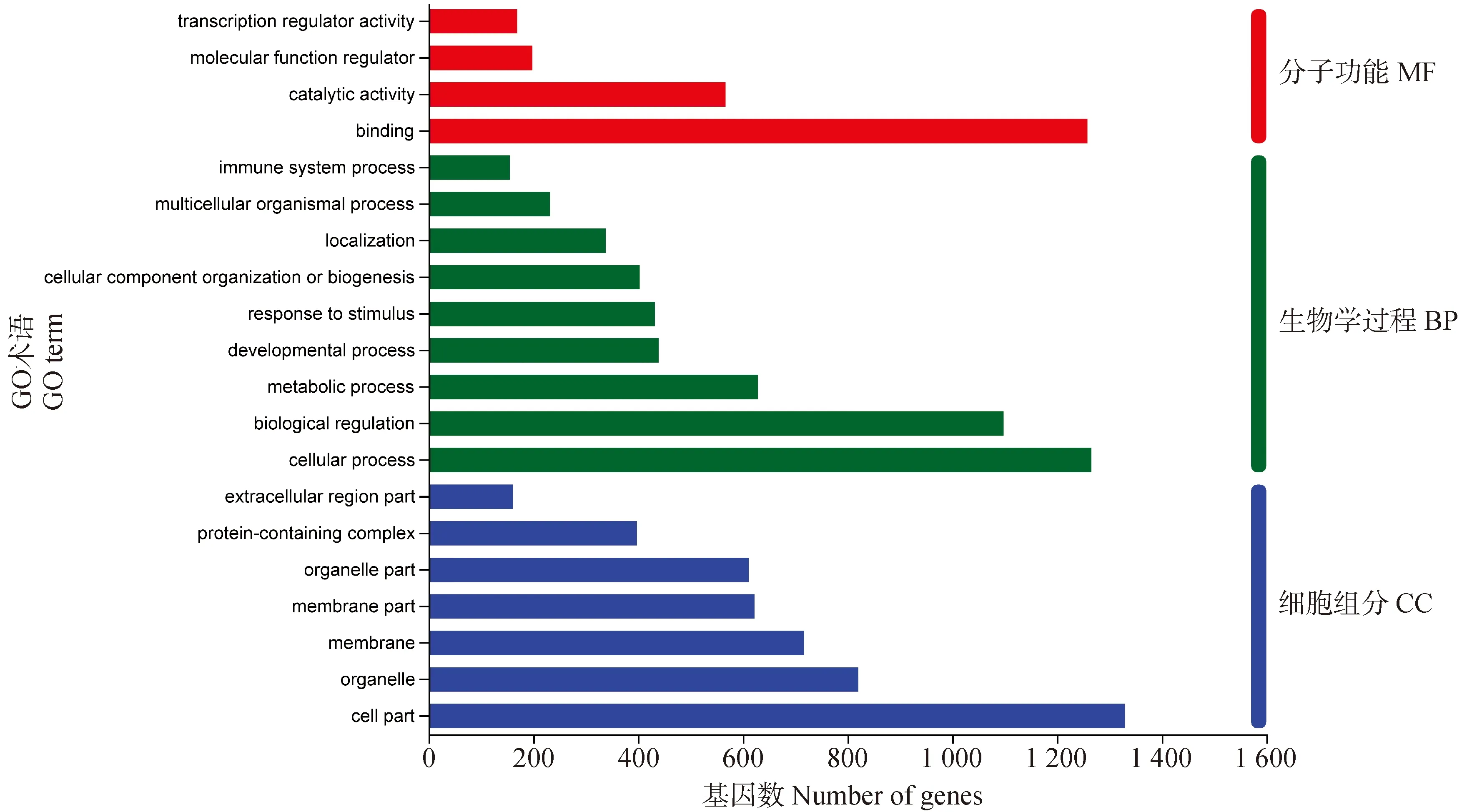
图1 差异表达基因GO注释分析Fig.1 GO annotation analysis of differentially expressed genes
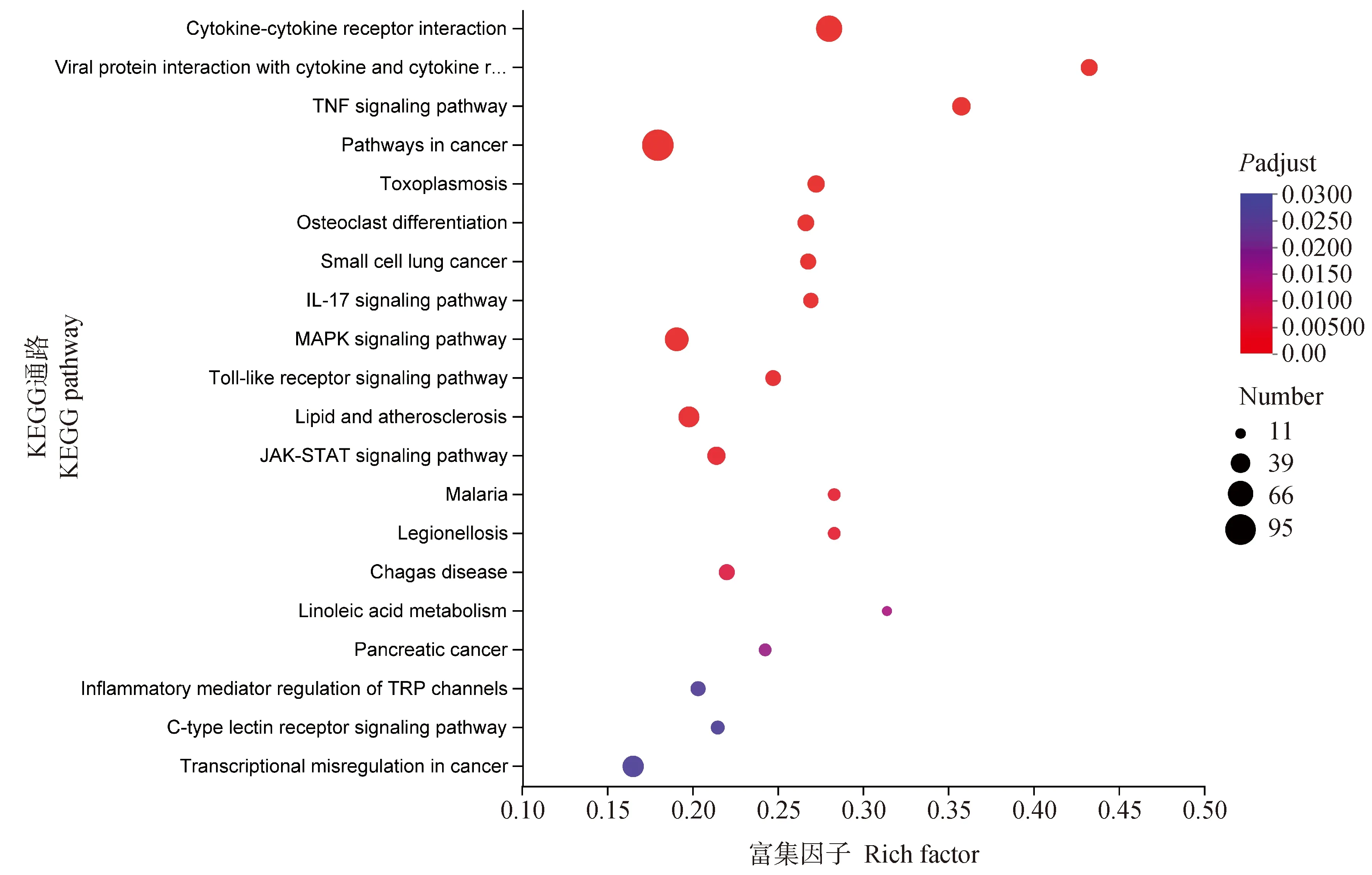
图2 差异表达基因KEGG富集分析气泡图Fig.2 Bubble map of KEGG enrichment analysis of differentially expressed genes
2.4 RT-qPCR验证
为了验证测序结果的可靠性,任意选取10个与天然免疫、抗病毒反应有关的差异表达基因,以β-Actin为内参基因,RT-qPCR检测其相对表达水平。结果如图3A所示,RT-qPCR的变化趋势与转录组测序结果一致,IL10、IFNG、IL8、IL1β、CCL20上调;CXCR4、CLP1、ALDOB、TNFSF10、CCR4下调。
本次转录组测序结果中,模式识别受体基因MDA5、TLR3、TLR7转录水平与对照组相比并无显著性差异(图3B)。
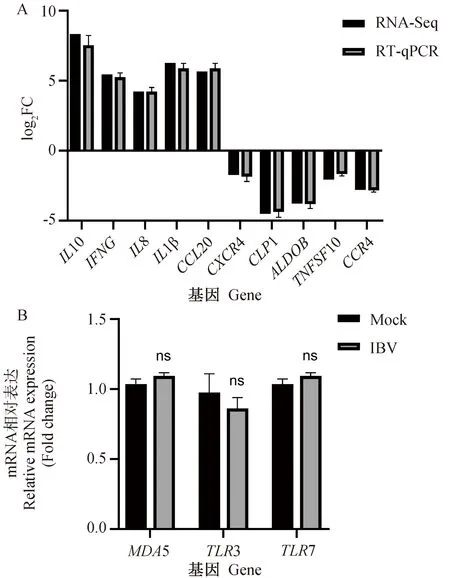
A. 差异表达基因的RT-qPCR验证;B. 模式识别受体基因,ns. P>0.05A. Differentially expressed genes; B. Pattern recognition receptors genes, ns. P>0.05图3 差异表达基因和模式识别受体基因的RT-qPCR验证Fig.3 Verification of differentially expressed genes and pattern recognition receptors genes by RT-qPCR
3 讨 论
模式识别受体(pattern recognition receptors, PRRs)是宿主先天免疫的重要组成部分,例如MDA5、TLR3、TLR7等,它们可以识别IBV复制产生的双链RNA和单链RNA,然后通过信号转导产生干扰素、促炎细胞因子,抑制IBV的复制[7-8]。本次转录组测序结果中,MDA5、TLR3、TLR7转录水平与对照组相比并无显著性差异(图3B),推测可能由于感染时间过短,IBV还未处于复制阶段,PRRs没有开始识别病毒。
通过对差异表达基因进行GO和KEGG分析发现,差异表达基因富集最显著的信号通路是细胞因子和细胞因子受体之间的相互作用,其中包括CCR4、IL8、IL1β、CCL20等参与免疫反应的基因,其中CXCR4在协调免疫反应方面发挥突出作用,它可以调节白细胞的运输和分布,与CCR5共同促进免疫突触的形成和稳定来支持T细胞启动[9-11]。TNF信号通路、MAPK信号通路也发生了显著的变化,TNF是炎症和免疫反应的关键介质和调节因子,它能够与受体TNF-R1相互作用激活不同的信号转导途径[12],富集到的TNF信号通路有34个差异表达基因,其中包括IRF1、TNFRSF1A、TRAF1、TRAF3等。MAPK能够将细胞表面的信号传递给细胞核内部,MAPK信号转导作为一种高度保守的信号通路参与各种生物事件[13]。TRAF6是一种连接蛋白,它可以直接激活MAPK信号通路[14]。本研究中,TRAF6在IBV感染2 h后显著上调,提示它在参与细胞的免疫反应中发挥了重要作用。
此次分析发现,大多数差异表达基因与免疫调控有关,表明细胞感染后通过调控免疫因子发挥抗病毒效应。例如JAK-STAT调节代谢过程信号通路几乎介导所有的免疫调节过程[15]。Toll样信号通路能够识别保守的微生物结构,哺乳动物的Toll样受体还诱导多种效应分子,直接破坏微生物病原体[16-17]。
综上所述,本研究采用转录组测序技术分析了IBV感染HD11细胞后参与免疫应答、抗病毒的相关基因,这些差异表达基因可能与IBV的致病机制或细胞的抗病毒反应有关。这将为研究IBV的防治提供一定的理论基础。
4 结 论
IBV感染HD11细胞后,感染组筛选出1 363个上调基因,653个下调基因,进一步GO和KEGG分析,发现IBV感染HD11细胞早期,差异表达基因大多与免疫反应、抗病毒作用有关,富集到的信号通路主要集中在细胞因子和细胞因子受体之间的相互作用、病毒蛋白与细胞因子或细胞因子受体之间的相互作用、TNF信号通路、MAPK信号通路等。本研究有助于探究IBV致病机制和感染早期宿主细胞免疫反应。
