初生及成年牦牛大脑皮质的转录组分析
2023-11-30余四九何俊峰潘阳阳
张 倩,崔 燕,余四九,何俊峰,潘阳阳,王 萌
(甘肃农业大学动物医学院,兰州 730070)
近年来,多项研究采用转录组学探究生物体发育阶段的基因调控,并把从转录组分析中获得的相关基因与转录本的功能表征结合[1-2],深入探究机体生长发育过程与遗传和环境间的互作关系。大脑皮质是感觉、自主运动和认知功能的中枢,高度有序的结构功能及发育依赖于基因在时空上精确的调控,其发育的异常会导致多种神经系统疾病的发生[3]。前期有研究对不同年龄人类[4]及小鼠[5]大脑皮质进行转录组分析,结果显示随着年龄增长突触活性和免疫功能上升。
牦牛是青藏高原高寒牧区的特有畜种,也是当地牧民获取奶、肉、皮等畜产品的重要来源,高寒、低氧、强辐射是青藏高原最显著的环境特征,牦牛各器官对低氧形成了其独特的形态结构和生理机制,尤其体现在呼吸和心血管系统[6-7]。梁林[8]报道,高原低氧牦牛脑组织对低氧产生了适应性,其大脑皮质分层数和神经元类型与其他哺乳类动物大体相同,发育特点与其他哺乳类动物也相似,皮质厚度随着年龄的增长逐渐增加,大脑皮质微血管密度均体现出随年龄增长而逐渐上升的趋势。此外,黄兴[9]运用转录组技术对牦牛大脑低氧适应性进行了研究,结果显示随着海拔升高,牦牛的转录本数量逐渐增加,表达网络更加复杂,具有更多差异显著的低氧适应相关通路,而低海拔牛的表达调控相对单一。近年来,学者们除了关注牦牛脑组织的低氧适应性,也开始关注在高原环境下牦牛各器官的生长发育机制,目前关于不同发育阶段牦牛转录组学研究仅限于瘤胃[10]、肝脏[11]、肺脏[12]和肌肉[13]的发育机制。基于此,本研究对初生和成年牦牛大脑皮质进行转录组分析,旨在深入挖掘影响牦牛大脑皮质生长发育的因子,最终为进一步探究高原动物大脑的生长发育机理提供一定依据。
1 材料与方法
1.1 试验材料
试验动物分为初生(1~7日龄,NYB)和成年(3~4岁,AYB)组的健康雄性青海高原牦牛,每组3 个生物学重复。2021年7 月采自青海省西宁市,放血处死,快速采集初生及成年牦牛大脑皮质额叶组织置于液氮,之后转至-80 ℃保存备用。
1.2 牦牛大脑皮质转录组测序
选取初生和成年牦牛各3个大脑皮质组织样品,使用TRIzol 试剂(Invitrogen)提取总RNA,通过琼脂糖凝胶电泳检测RNA的完整性,利用Nano-Drop ND-1000 (Thermo scientific)和2100 Bioanalyzer(Agilent)分别对RNA的量与纯度进行质控,之后使用 Illumina Hiseq xten测序平台进行高通量测序。
1.3 牦牛大脑皮质转录组测序分析
运用FastQC和 Trim-Glore软件对原始数据进行质控和过滤,之后借助HISAT2 软件 (https://daehwankimlab.github.io/hisat2/,版本为hisat2-2.0.4)[14]将得到的Clean Data比对到牦牛参考基因组上(版本为 BosGruv3. 0),然后使用Htseq-count[15]软件对Clean Reads进行计数,再用String Tie软件 (版本为1.3.4d.Linux_x86_64)[16]进行FPKM定量。使用R包DESeq2对样本进行差异显著分析[17],将P<0.05和|log2(fold change)|≥1作为筛选条件,之后获得差异表达基因(differentially expressed genes, DEGs),并用ClusterProfiler[18]软件对DEGs进行GO (Gene ontology)和KEGG (Kyoto encyclopedia of genes and genomes)富集分析。
1.4 qRT-PCR差异验证
为了验证转录组数据的可靠性,随机选出9个DEGs,通过qRT-PCR 技术进行验证分析,采用Primer 5.0软件设计引物(表1),以β-actin作为内参,采用2-ΔΔCT法进行分析[19],通过SPSS 24.0 统计软件对数据进行ANOVA分析。

表1 RT-qPCR 引物序列信息Table 1 Primers information for RT-qPCR
2 结 果
2.1 测序数据质控分析
本研究分别采集初生和成年的牦牛大脑皮质进行转录组测序,RNA 样品量>18 μg,纯度检测值(OD260 nm/OD280 nm)>2.0,RNA 完整性检测(RNA intergrity number, RIN)>9.0,说明所有 RNA 样本质检合格,符合建库要求。各样本过滤后的序列数据不少于4.75×107条,所测数据的碱基长度达到6.41 G以上,Q20 均大于 99.89%,Q30在97.26%以上,并且GC 含量在48.00%~51.00% (表2),比对到参考基因组的 Reads 与 Clean Reads 的比率在 86.53%以上,表明测序所得的数据准确可靠,可用于后续生物信息学分析要求。
2.2 初生及成年牦牛脑组织的DEGs筛选
在成年组相对初生牦牛的大脑皮质转录组分析中共获得4 790个DEGs,其中包括2 670个上调表达的DEGs,2 120个下调表达的DEGs(图1)。
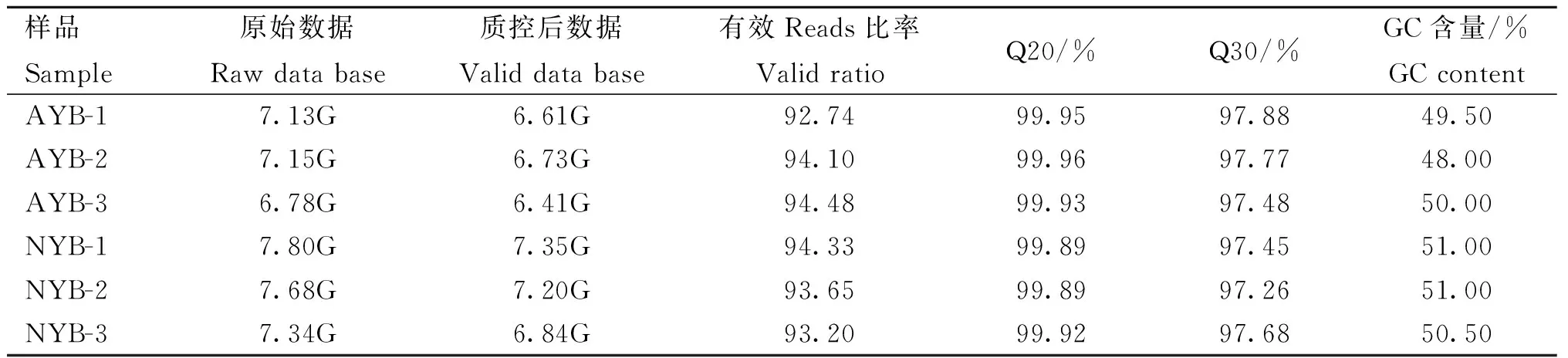
表2 转录组测序数据质量检测分析
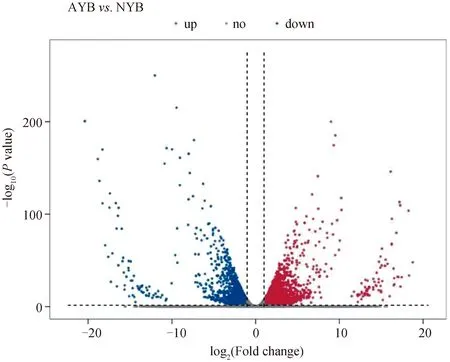
虚线左侧代表显著下调基因,虚线右侧代表显著上调基因。NYB代表初生组;AYB代表成年组 The left side of the dotted line represents significantly down-regulated genes, and the right side of the dotted line represents significantly up-regulated genes. NYB stands for the newborn group; AYB stands for the adult group图1 不同年龄牦牛大脑皮质发育中DEGs火山图(AYB vs. NYB)Fig.1 Volcano map of differentially expressed genes in the yak cerebral cortex at different age (AYB vs. NYB)
2.3 差异表达基因GO功能聚类
对在成年组和初生组牦牛大脑皮质 (AYBvs. NYB) 中4 790个DEGs进行GO 功能分类富集,共富集到 8 751个GO条目,包括845个细胞组成 (cellular component, CC),1 734个分子功能 (molecular function, MF),6 172个生物过程 (biological process, BP)。前50条显著富集到的GO条目如图2所示,在BP分类中,其中占比例最大的3个二级条目从高到低为转录调控(255个基因),RNA聚合酶II正调控转录(237个基因),蛋白质磷酸化(166个基因)。在CC分类中,细胞质(598个基因)占比例最多,其次为质膜(564个基因)和细胞核(554个基因)。在MF分类中,其中占比例最大的3个二级条目为蛋白结合(806个基因),相同的蛋白结合(274个基因)以及ATP结合(272个基因)。
2.4 差异表达基因KEGG富集分析
KEGG富集分析的结果表明,在成年组相对初生组牦牛大脑皮质筛选出的DEGs主要在325条信号通路,图3中列出了前20条显著富集到的通路,分别为PI3K-Akt信号通路(129个基因)、MAPK信号通路(119个基因)、HIF信号通路(90个基因)和轴突导向(85个基因)等。
2.5 qRT-PCR 验证
为了进一步检测 RNA-Seq 所识别 DEGs 的可靠性,9个DEGs被随机筛选进行 qRT-PCR验证,如图 4 显示,以初生牦牛为对照,成年牦牛VEGFA、PDGFA、IGF1、AKT1、ERK1、BDNF、NGF及Slit2显著上调;HIF1α显著下调,上述挑选的DEGs表达模式均与 RNA-Seq 数据一致。
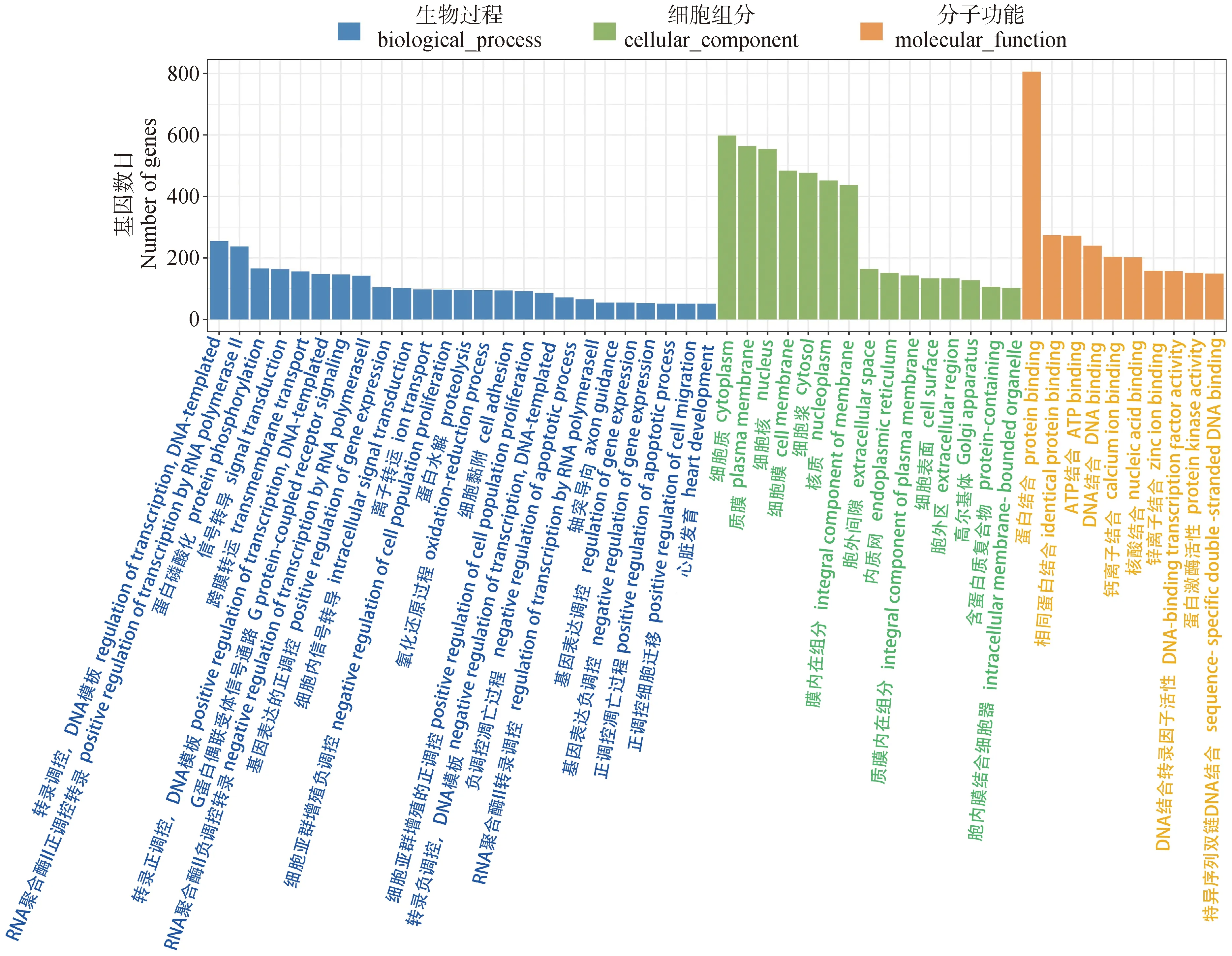
图2 不同年龄牦牛大脑皮质发育中DEGs富集分析中前50个GO条目 (AYB vs. NYB)Fig.2 Top 50 GO items in DEGs enrichment analysis of cerebral cortex development in yaks at different ages (AYB vs. NYB)
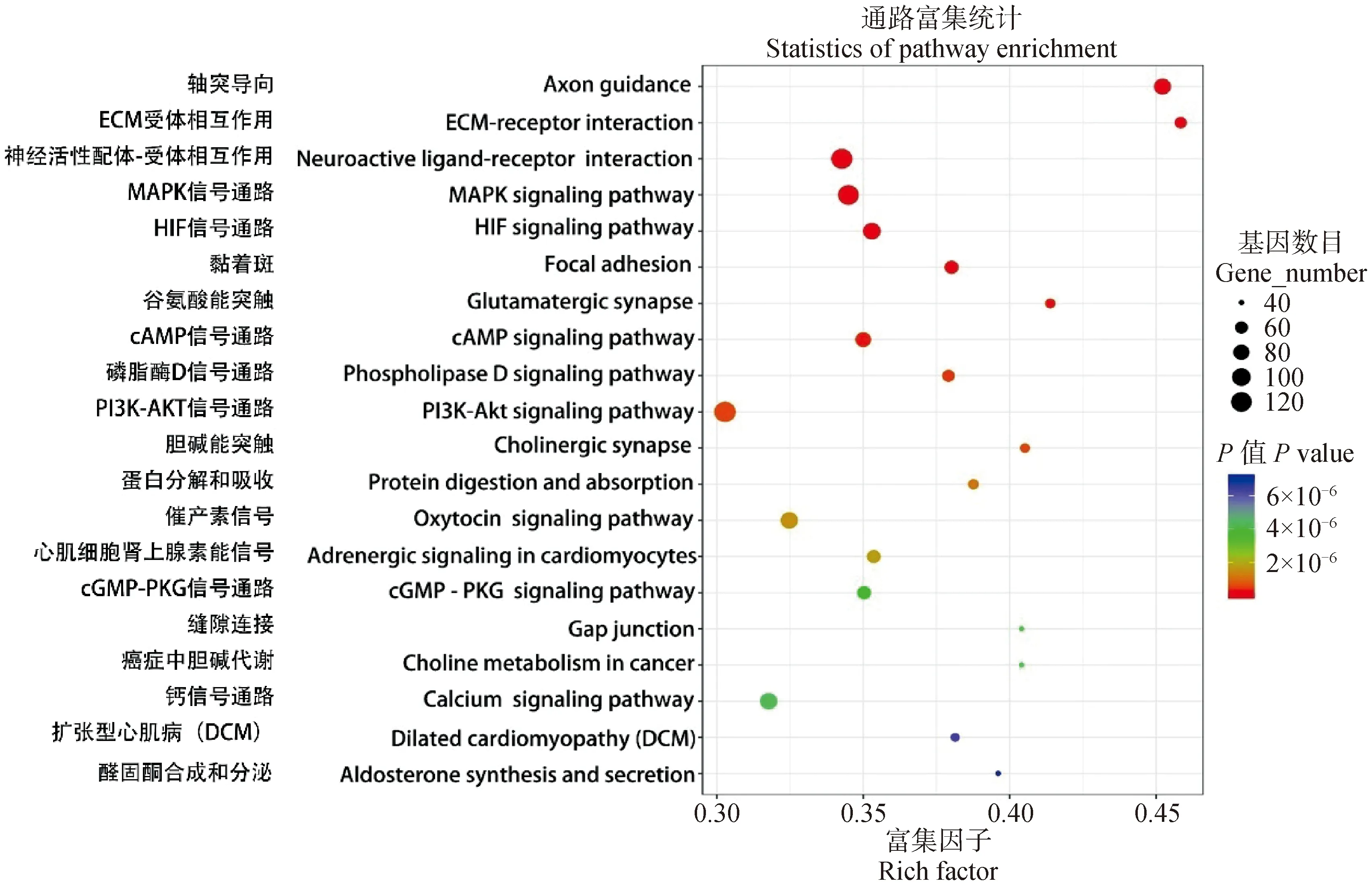
图3 不同年龄牦牛大脑皮质发育中DEGs富集分析中前20条KEGG通路 (AYB vs. NYB)Fig.3 Top 20 KEGG pathways in DEGs enrichment analysis of cerebral cortex development in yaks at different ages (AYB vs. NYB)
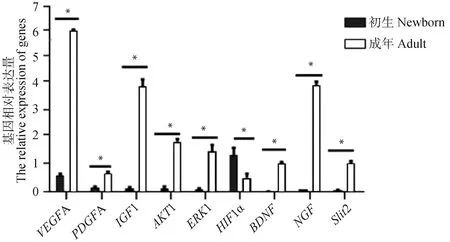
*. P<0.05图4 qRT-PCR检测9个DEGs在初生及成年牦牛大脑皮质的表达模式Fig.4 qRT-PCR verification of the expression tendency of 9 DEGs in the cerebral cortex of newborn and adult yaks
2.6 调控大脑皮质发育的候选基因筛选
通过筛选KEGG富集到的信号通路,共得到与神经元神经突发育及可塑性、突触囊泡传递、胶质细胞增殖、免疫防御及低氧适应相关的13个DEGs,其中11个上调基因,2个下调基因(表3、图5)。

表3 参与调控大脑皮质发育的信号通路及基因Table 3 Signaling pathway and genes realated to the development of yak cerebral cortex
3 讨 论
本研究利用RNA-seq技术对初生和成年牦牛大脑皮质进行了转录组测序,筛选出了2 670个上调表达的差异基因,2 120个下调表达的差异基因,并对DEGs进行了GO功能分类富集,分析发现占比最多的条目是转录调控、RNA聚合酶II 正调控转录、蛋白结合和ATP结合,大部分都与能量产生、转录和翻译过程紧密相关,说明成年牦牛大脑皮质细胞的蛋白质合成和能量利用较为活跃,提示成年牦牛神经递质生成、释放等生理活动较旺盛。
KEGG富集分析得到了DEGs的主要富集通路,包括PI3K-Akt、神经活性配体-受体相互作用、 MAPK、轴突导向、钙信号、磷脂酶D信号、突触囊泡运输、Toll样受体等与大脑发育相关的通路,这与前人对其他动物大脑增龄性转录组分析结果相似[3-5],但有意思的是,本研究显示HIF信号通路也在TOP通路中,这与其他动物不一样,可能与牦牛常年生活在高原低氧环境相关,有研究报道低氧诱导因子通路是重要的缺氧适应性调节因子信号,在缺氧条件下HIF触发多种基因的表达,这些基因能够启动红细胞生成、血管生成、糖代谢及转运、细胞存活等,可在低氧脑组织起到保护作用[20]。Wang等[21]和Zhang等[22]报道,PI3K-AKT 是神经细胞增殖、生长、存活和代谢的正向调节信号,可以将细胞外刺激信号转导至细胞核,激活核内转录因子,调控应激蛋白基因的表达,促进有关蛋白质的合成,如神经营养因子生长及神经细胞表面受体等,完成对胞外刺激的反应蛋白。神经活性配体-受体相互作用信号通路主要存在于突触小泡、轴突末端和树突的胞浆中,促进突触传递[4]。轴突导向因子信号不仅能够调控轴突的生长,还能定向细胞迁移,帮助在大脑损害后起到抗凋亡、诱导细胞的迁移、调节轴突方向形成作用[23]。钙信号在突触传递和改变神经突动力学方面发挥功能[24]。磷脂酶D信号能够参与神经递质传递和星型胶质细胞增殖[25]。突触囊泡运输信号能够决定神经递质运输的特异性[26]。MAPK信号具有广泛的神经发育和生理功能,在神经发育过程中调节神经组织的细胞增殖和分化是至关重要的,尤其能够促进神经胶质细胞增殖及迁移,有利于损伤修复[27]。上述信号通路被富集,说明成年牦牛大脑皮质中突触发育及可塑性、神经递质传递、胶质细胞增殖、免疫防御、低氧适应相关的功能非常活跃,即成年牦牛大脑皮质发育可能更完善。
在KEGG富集排名较前且符合大脑发育通路中,挑选显著上调或下调P值最小的基因,筛选出与牦牛大脑皮质生长发育相关的13个候选基因,有研究表明这些基因均与大脑发育相关。其中SEMA5A和Slit2在成年牦牛大脑皮质高表达,Carulli等[28]报道SEMA5A有两个功能相反的结构域TSP和SEMA,通过与细胞外基质中特定蛋白多糖的互作介导轴突延伸或抑制过程;Slit2可以引导轴突选择正确途径,从而成功到达靶区,促进神经元、轴突与靶器官之间形成精准联系[29],以上提示成年牦牛可能轴突调节较活跃以及突触传递较准确。另外,FGF18在成年牦牛大脑组织内表达显著上调,Klimaschewski和Claus[30]研究发现,FGF18在胶质细胞形成过程中起着神经元源性胶质细胞生长因子的作用,对星形胶质细胞和小胶质细胞都具有有丝分裂活性,说明成年牦牛先天免疫能力和血脑屏障可能更完善,具有调节损伤和修复的能力。同样,作为表达量显著上调的BDNF,它在维持神经元的生长及分化过程中至关重要,尤其是对多巴胺能神经元具有神经保护和神经恢复作用,还可以调节神经突向外生长和维持突触连接稳态,从而维持神经元的正常功能和突触可塑性[31],此基因的高表达提示其可能与大脑皮质发育密切相关。Rab3a大量分布在囊泡前神经末端,与突触囊泡膜结合,通过调节囊泡运输过程的定向和捆绑过程,来决定囊泡转运过程的特异性[26],本研究发现Rab3a在成年牦牛大脑皮质显著上调,说明成年牦牛可能具有更为精准的囊泡运输。VEGFA能促进血管生成发挥营养作用,在发育和成年期可以刺激神经突生长和成熟,以及星型胶质细胞增殖、迁移和存活,成年牦牛大脑皮质VEGFA表达量上升,表明成年牦牛的星型胶质细胞可能发育更完善,有利于血脑屏障的形成及稳定[32]。NGF在成年牦牛大脑皮质显著上调,Rocco等[33]报道NGF对中枢神经系统胆碱能神经元的神经突生长、发育和存活至关重要,有研究报道在胎儿生长过程中,NGF合成异常会导致中枢神经系统发育异常,长期影响神经元连接及信息传递,提示成年牦牛脑组织内胆碱能神经元神经突的发育更为完善。CaMK2A在大脑皮质的突触后致密部含量丰富,通过和 N-甲基-D-天冬氨酸受体,或其他靶结构相结合补充到突触后致密区,增加了突触部位α-氨基-3-羟基-5-甲基-4-异唑丙酸的锚定地点,从而增强了突触传递;另外,在轴突的生长锥中有CaMK2A 蛋白的表达,而CaMK2A具有 F-actin 的结合区域,CaMK2A可能通过调节 F-actin 达到影响神经突延伸、影响神经突树枝状化及改变神经突动力学的目的[24],因此本研究中成年牦牛CaMK2A高表达可能提示其突触可塑性和突触传递功能更完善。PLD1在成年牦牛大脑皮质显著上调,研究表明它们在大脑发育中可以促进突触和髓鞘的形成,同时神经递质中PLD促进突触囊泡融合以及参与囊泡运输[25],说明成年牦牛神经突形成及递质传递更完善。Toll样受体是机体一线先天防御反应的一部分,它构成了一个早期预警系统,刺激T淋巴细胞和抗体介导的适应性反应,神经元、小胶质细胞和星形胶质细胞也表达各种TLR,它们有助于大脑皮质的的免疫保护[34],成年牦牛大脑皮质TLR1、TLR4表达较高,可能提示其调节损伤和免疫防御的能力强于初生牦牛。HIF家族是重要的低氧适应调控因子,HIF1α和HIF2α在缺氧时表达量会显著上升,促进多种生存途径,包括神经保护、血管生成和神经营养素,减少细胞死亡[20,35],而本研究发现这两个因子在成年牦牛大脑皮质显著下调,说明成年牦牛脑组织可能已经适应了低氧。
4 结 论
综上,本研究利用RNA-seq对初生及成年牦牛大脑皮质发育的转录组进行了分析,筛选到了13个与大脑皮质发育相关的候选基因,其中Slit2、SEMA5A、BDNF、Rab3a、FGF18、NGF、PLD1、VEGFA、CaMK2A、TLR1、TLR4在成年牦牛大脑皮质显著上调,可能提示成年牦牛轴突可塑性、突触传递、免疫防御等生理活动更完善,另外与低氧适应调控相关的HIF1α和HIF2α在成年牦牛显著下调,可能表明成年牦牛大脑皮质对低氧适应性更强,以上结果为进一步研究高原动物大脑发育的分子机制奠定了理论基础。
