低水温下鳜短期饥饿的生理动态变化
2023-11-22于俊琦张皓迪李洪琴刘天骥刘
于俊琦 陶 鹏 张皓迪 李 虹 李洪琴 罗 浩 刘天骥刘 匆 郑 轲 罗 莉
(1.西南大学水产学院,淡水鱼类资源与生殖发育教育部重点实验室,水产科学重庆市重点实验室,重庆 400715;2.重庆市水产技术推广总站,重庆市生态渔产业技术体系,重庆 400400;3.四川新希望六和科技创新有限公司,成都 610000;4.江西天之佳生物科技有限公司,南昌 331700)
鳜(Siniperca chuatsi)俗称“桂花鱼”,隶属于鲈形目,是我国特有的名优鱼类,因其肉质鲜美被大众所喜爱。据2022年渔业统计年鉴显示,鳜养殖产量已经达到37.4万吨[1],是近几年来在我国发展迅速的名优养殖品种之一。在鳜传统养殖过程中,养殖户在冬季通常一次性放入活饵料鱼,保证鳜安全越冬,但因鳜存塘量估算缺乏精准性,则可能存在饵料不足处于饥饿状态。为响应水产绿色健康养殖五大行动之一的配合饲料替代冰鲜杂鱼,本研究团队在重庆各地区对于饲料鳜健康养殖进行示范推广,并实现了冬季低水温饲料鳜健康养殖新模式[2]。在冬季饲料鳜养殖过程中如果面临短期饥饿,则会导致鱼体消瘦、易患病[3,4],对养殖生产造成隐形损失。所以本课题组对鳜在低水温(13±1)℃下短期(15d)饥饿和摄食的生理状态进行比较,摄食相较于饥饿增重量差值达到15.93%,表明低水温下鳜摄食可以维持体重的增长,饥饿则会使鳜体重和消化、免疫、抗氧化能力降低[3]。
为进一步探究在低水温下,15d饥饿胁迫对鳜生理指标造成负面影响的动态变化过程,本研究依据鳜主产区之一的广东省冬季的低温期水温(10—16)℃,在(13±1)℃水温条件下,进行15d的饥饿实验,以采样时间分组D0、D3、D6、D9、D12和D15。研究低水温下短期饥饿对鳜形体指标、肌肉组成、血液生理生化指标、消化能力、肝脏和鳃抗氧化能力、肝脏脂质代谢水平等指标的动态生理变化,旨在探究鳜在低水温下短期饥饿的生理状态变化,丰富鳜生理学研究内容,为其在越冬期科学合理采用配合饲料的饲养管理提供参考依据。
1 材料与方法
1.1 试验前期驯化饲料
根据鳜的营养需求,前期驯化饲料配方及主要营养成分如表1所示。

表1 试验前期驯化饲料配方及主要营养成分Tab.1 Formula and main nutrients of acclimated feed in the early stage of the experiment
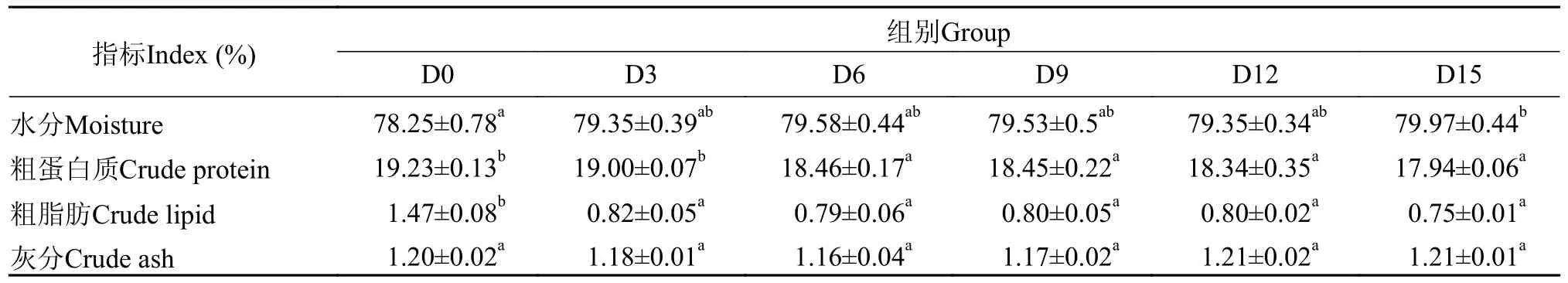
表2 低水温下短期饥饿对鳜肌肉组成的影响(鲜样基础)Tab.2 The effect of short-term starvation on the muscle composition of mandarin fish under low water temperature (fresh matter basis)
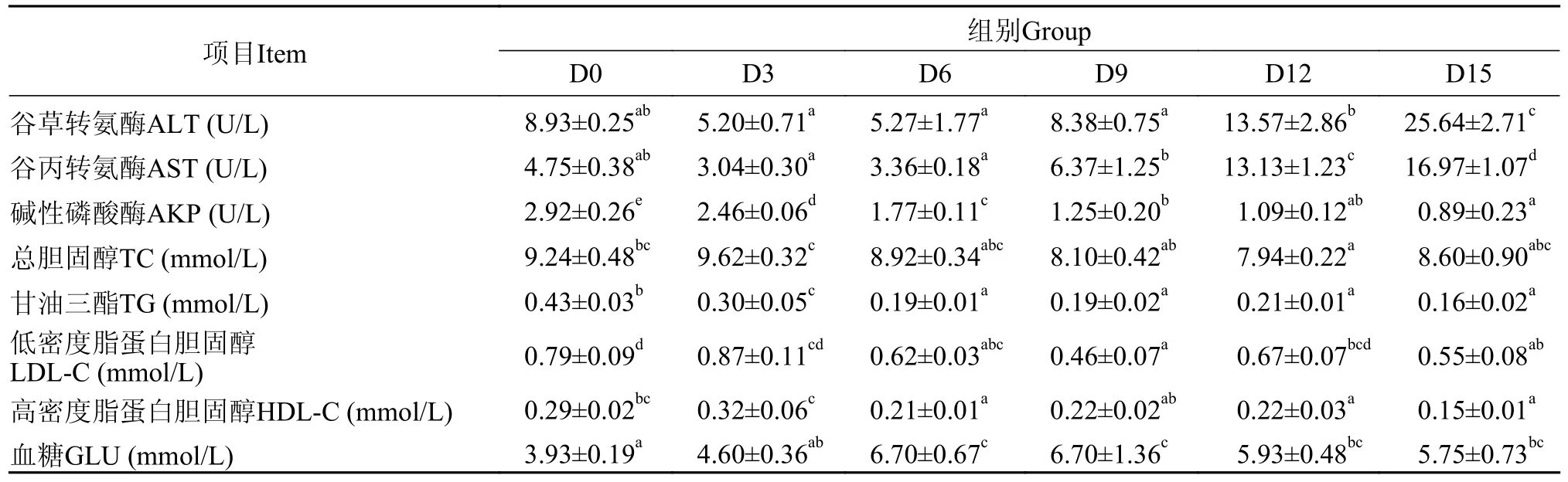
表3 低水温下短期饥饿对鳜血液生化指标的影响Tab.3 The effect of short-term starvation on blood biochemical indexes of mandarin fish at low water temperature
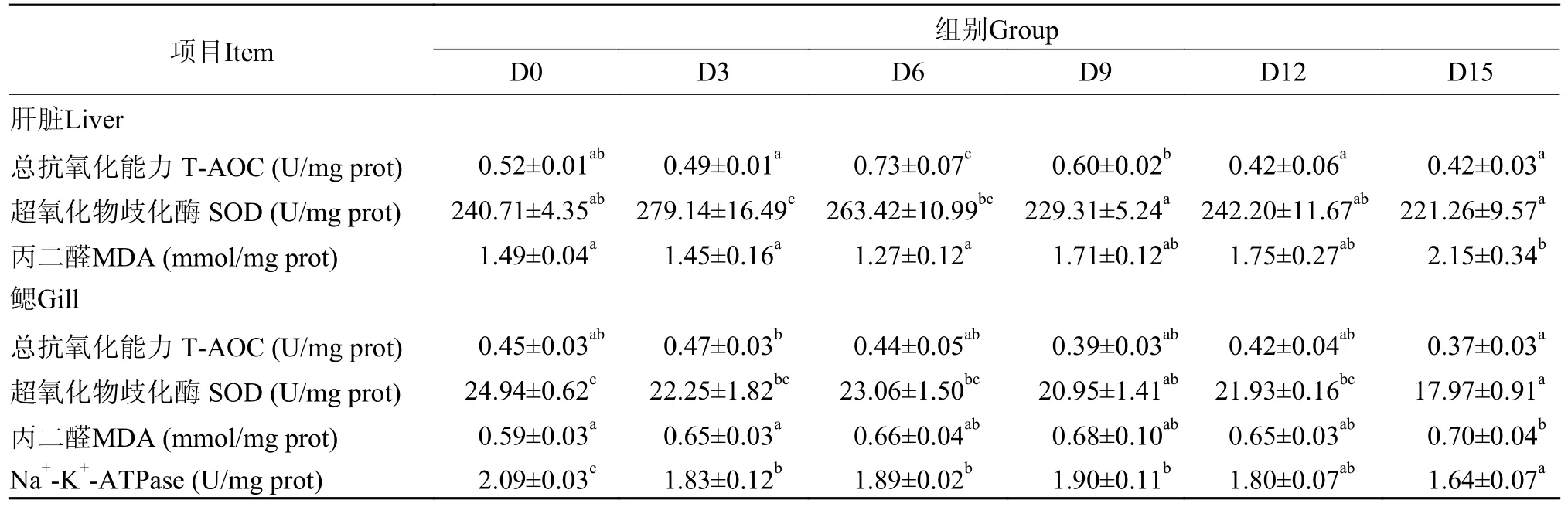
表4 低水温下短期饥饿对鳜肝脏和鳃抗氧化能力的影响Tab.4 The effects of short-term starvation at low water temperature on the antioxidant capacity of mandarin fish liver and gills
1.2 试验鱼及饲养管理
试验所需鳜购于重庆市潼南区兴水渔场,共1000尾,运回实验室先静养2d,再用1.0%的NaCl溶液浸泡消毒15min,饥饿2d后放置于3个5 m3的养殖缸内驯养30d,期间投喂自配鳜专用配合饲料(配方见1.1),驯化15d达到90%的鳜转食配合饲料后,每天投喂2次(上午9: 00,下午17: 00)进行15d饱食投喂。取转食饲料后的健康鳜270尾用于试验,均重(84.13±0.14) g。实验设计D0、D3、D6、D9、D12和D15共6个组,分别饥饿0、3d、6d、9d、12d和15d,每组3个重复,每个重复15尾,共计270尾,饲养于西南大学水产学院国家示范中心循环水养殖系统的18个圆形养殖缸(0.5 m3)中,正式试验时间为15d,在饥饿试验开始后,每天吸污1次,以确保水质清洁,光周期为自然周期,水温(13±1)℃,溶解氧≥6.0 mg/L,氨氮含量≤0.50 mg/L,亚硝酸盐含量≤0.03 mg/L,pH 6.5—7.5。
1.3 样品采集
分别在鳜饥饿的0、3d、6d、9d、12d和15d,每组每个重复随机选取5尾鳜用50 mg/L的MS-222麻醉,称重后用1 mL一次性无菌注射器在尾静脉处取血,用罗氏活力型血糖仪测定血糖后,其余血液转移至离心管中4500 r/min离心10min制备血浆放入液氮罐速冻后,转入-80℃冰箱内保存,用于血浆生化指标测定。取血后将鳜冰上解剖,取鳃、内脏团和背部肌肉,将肝脏、胃、脾脏和肠道分离并称重,液氮速冻放入-80℃冰箱保存备用。指标测定前,取组织按1∶9 (g/mL)或1∶4 (g/mL)加入生理盐水,迅速转入高速分散器匀浆(匀浆过程在冰水浴中进行),后将匀浆液离心(4℃,4000 r/min,10min),取上清液制得粗酶液,分装存于-80℃冰箱待用。
1.4 指标测定
形体指标: 饥饿实验结束后,准确称量各组鳜体质量、内脏重及肝重,计算公式如下:
脏体比(Viscerosomatic index,VSI)=Wv/W×100;
肝体比(Hepatosomatic index,HSI)=Wl/W×100;
肥满度(Condition factor,CF,g/cm3)=W/BL3×100;式中,W为每组鱼体质量(g);Wv为内脏重(g);Wl为肝脏重(g);BL为鱼体长(cm)。
肌肉常规营养成分: 水分含量测定采用105℃烘箱干燥法(GB/T 6435-2006)、粗蛋白测定采用凯氏半微量蒸馏定氮法(GB/T 6432-1994)、粗脂肪测定采用索氏抽提法(GB/T 6433-1994)、粗灰分采用550℃灼烧法(GB/T 6438-1992)。
血液生化指标均采用南京建成生物工程研究所试剂盒进行测定。其中谷丙转氨酶(ALT)采用赖式法(货号: C009-1),谷草转氨酶(AST)采用比色法(货号: C010-1),碱性磷酸酶(AKP)采用比色法(货号: A059-1),甘油三酯(TG)采用GPO-PAP法(货号:A110-2),胆固醇(TC)采用COD-PAP法(货号: A111-2),高密度脂蛋白胆固醇(HDL-C)和低密度脂蛋白胆固醇(LDL-C)采用直接法(货号分别为A112-2、
A113-2)。
消化指标均采用南京建成生物工程研究所生产的试剂盒测定,其中胃蛋白酶采用比色法(货号:A080-1);胰蛋白酶采用紫外比色法(货号: A080-2);脂肪酶采用比色法(货号: A054-1);淀粉酶采用淀粉-碘比色法(货号: C016-1)。
抗氧化指标均采用南京建成生物工程研究所试剂盒进行测定,其中总抗氧化能力(T-AOC)采用比色法(货号: A015-1);总超氧化物歧化酶(T-SOD)采用羟胺法(货号: A001-1);丙二醛(MDA)采用TBA法(货号: A003-1)。
脂质代谢指标均采用上海优选生物科技有限公司Elisa试剂盒进行测定。肝脏肉碱脂酰转移酶-1(CPT-1)和乙酰辅酶A羧化酶(ACC)采用酶联免疫吸附法(货号分别为YX-22445F、YX-22438F)。
H+-K+-ATP酶、Na+-K+-ATP酶均采用南京建成生物工程研究所试剂盒(定磷法)进行测定(货号分别为A069-1和A016-2)。
1.5 数据处理
试验结果用SPSS 22.0对数据进行方差齐性检验和单因子方差分析(One-way ANOVA,LSD),用Tukey多重比较分析组间差异显著性程度,显著水平为(P<0.05)。数据用平均值±标准差(mean±SD)形式表示。
2 结果
2.1 低水温下短期饥饿对鳜形体指标的影响
低水温下短期饥饿能显著影响鳜的形体指标(P<0.05),由图 1可知随着饥饿时间的延长,BW、CF、VSI和HSI都呈现明显的下降趋势(P<0.05),但下降程度不一。其中BW在饥饿3d、9d和12d时出现显著性下降,CF在饥饿9d时出现显著性下降,HSI和VSI都在饥饿6d时显著性下降(P<0.05)。
2.2 低水温下短期饥饿对鳜肌肉组成的影响
由表 1可知,在低温和短期饥饿的胁迫下,鳜肌肉组成有显著性差异(P<0.05),肌肉粗脂肪含量则在饥饿3d时显著下降(P<0.05),比初始下降44.22%,而3—15d保持稳定,无显著变化(P>0.05)。肌肉蛋白质含量分别在饥饿6d、9d、12d和15d 时下降4.00%、4.10%、4.63%和6.71% (P<0.05)。肌肉粗灰分含量在饥饿15d内相对稳定(P>0.05)。
2.3 低水温下短期饥饿对鳜血液生化指标的影响
由表 2可知,低水温短期饥饿15d内,鳜血浆ALT、AST活性随饥饿时间的延长呈先下降后上升的趋势,而AKP活性、TG、TC、LDL-C、HDL-C含量呈整体下降趋势。其中血浆ALT、AST活性都在饥饿3d时降到最小值,AKP活性和TG含量逐渐下降,TC、LDL-C含量分别在饥饿9d、12d时降到最小值,HDL-C含量在饥饿6d时显著下降(P<0.05),之后保持稳定,直到15d时再次降低。GLU在饥饿6d时显著上升(P<0.05),然后维持在一定的水平范围。
2.4 低水温下短期饥饿对鳜消化能力的影响
由图 2可知,鳜胃H+-K+-ATP酶活性随饥饿时间的延长前3d上升,但与初始无显著差异(P>0.05),3d后呈持续下降的趋势,15d时显著性下降到最小值(P<0.05);胃蛋白酶活性在3d、6d时出现显著性下降(P<0.05),并在15d时显著性下降到最小值(P<0.05),两者分别比初始下降88%和39%。在图 3和图 4 中,鳜肠道和幽门盲囊的胰蛋白酶、脂肪酶、淀粉酶活性都随饥饿时间的延长呈整体下降趋势,这些消化酶的酶活性都在饥饿15d时达到最小值。其中肠道和幽门盲囊的胰蛋白酶活性都在饥饿前3d无显著性差异(P>0.05),肠道和幽门盲囊的淀粉酶活性分别在饥饿前9d、6d无显著差异(P>0.05),肠道和幽门盲囊脂肪酶活性在饥饿6—12d无显著性差异(P>0.05)。
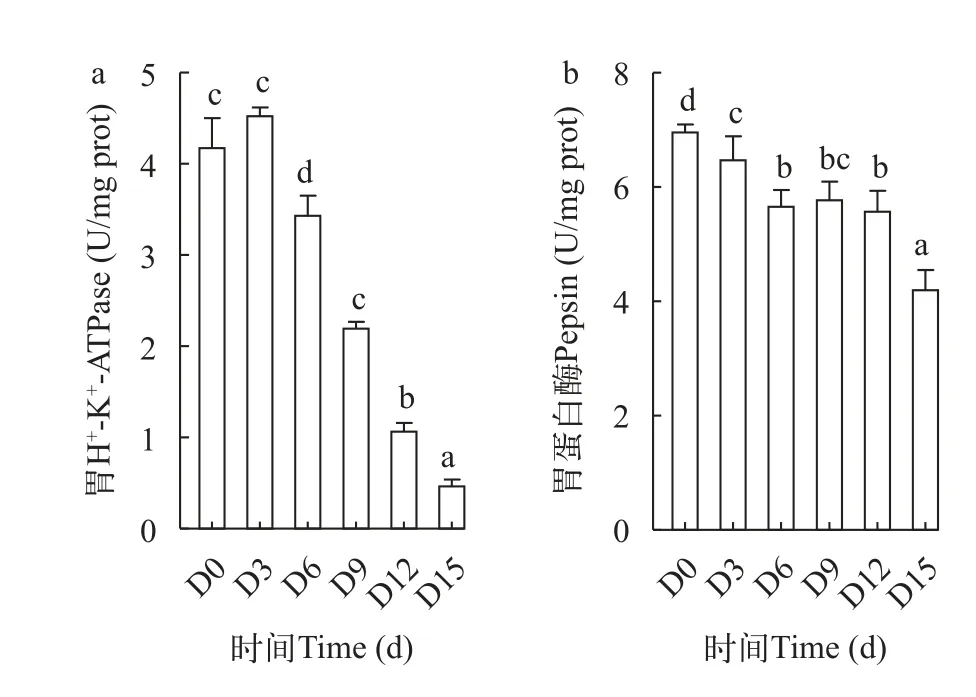
图2 低水温下短期饥饿对鳜胃H+-K+-ATPase(a)和胃蛋白酶(b)的影响Fig.2 The effects of short-term starvation on H+-K+-ATPase (a)and pepsin (b) in mandarin fish at low water temperature
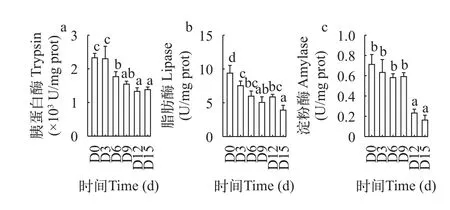
图3 低水温下短期饥饿对鳜肠道胰蛋白酶(a)脂肪酶(b)和淀粉酶(c)的影响Fig.3 The effects of short-term starvation at low water temperature on intestinal trypsin (a) lipase (b) and amylase (c) of mandarin fish
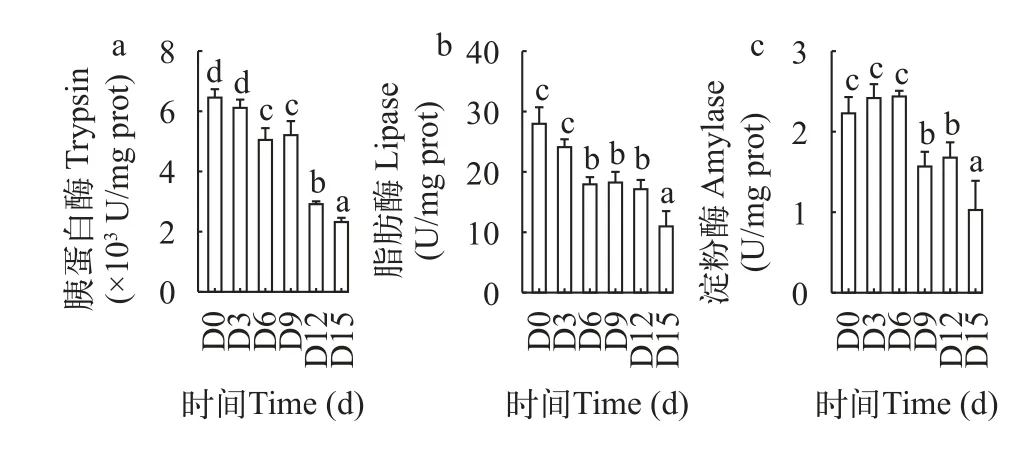
图4 低水温下短期饥饿对鳜幽门盲囊胰蛋白酶(a)、脂肪酶(b)和淀粉酶(c)的影响Fig.4 The effects of short-term starvation on trypsin (a),lipase(b) and amylase (c) in the pyloric blind sac of Mandarinfish at low water temperature
2.5 低水温下短期饥饿对鳜肝脏和鳃抗氧化能力的影响
由表 3可知,在低水温下,短期饥饿对鳜肝脏和鳃抗氧化能力及鳃Na+-K+-ATP酶活性有显著影响(P<0.05),随饥饿时间的延长,鳜肝脏T-AOC含量、T-SOD活性出现先上升后下降趋势,分别在饥饿6d和3d时达到最大值,两者都在饥饿15d时达到最小值。肝脏MDA含量、鳃T-AOC含量和T-SOD活性都在饥饿前期无显著性变化(P>0.05),并维持在一定的水平范围,饥饿后期肝脏MDA含量显著性升高(P<0.05),到15d时达到最大值,比初始0d上升44.30%,而鳜鳃T-AOC含量和T-SOD活性则显著性下降(P<0.05),到15d时下降到最小值。此外,鳃MDA含量在饥饿期间逐渐上升,到15d时显著性上升到最大值(P<0.05),而Na+-K+-ATP酶活性则在饥饿3d时显著性下降(P<0.05),之后到饥饿前12d都无显著性差异(P>0.05),直到饥饿15d时,Na+-K+-ATP酶活性显著性下降到最小值(P<0.05)。
2.6 低水温下短期饥饿对鳜肝脏脂质代谢的影响
由表 4可知,在低温条件下,15d短期饥饿对鳜肝脏脂质代谢有显著性影响(P<0.05)。随着饥饿时间的延长,鳜肝脏肉碱脂酰转移酶呈整体下降趋势,在饥饿3d时显著性下降(P<0.05),在15d时显著性降低到最小值(P<0.05)。乙酰辅酶A羧化酶活性呈整体上升趋势,在饥饿6d时显著性上升(P<0.05),在15d时显著性上升到最大值(P<0.05;表 5)。
3 讨论
3.1 低水温下短期饥饿对鳜形体指标及肌肉组成的影响
鱼类在饥饿条件下,由于没有外源能量的补充,只能不断消化自身含有的营养物质来维持生命活动所需的能量代谢,从而造成鱼体外部形态特征和内部解剖性状产生某些特定变化,如鱼体质量出现负增长、肠管变细、肝胰脏缩小等[5],鱼体通过改变自身的形体指标来适应这种环境的变化,以保证形体结构和机能相协调[6]。在本实验中,当饥饿15d时,鳜体质量较初始时显著下降,这与罗非鱼(Oreochromis niloticus)[6]、彭泽鲫(Carassius auratusvar Pengze)[7]、虹鳟(Oncorhynchus mykiss)[8]上的研究结果类似。内脏和肝脏都是鱼类存储营养物质的部位,在饥饿胁迫时,动用肝脏和内脏中的脂肪、蛋白和糖原,导致相应形态指标的下降。钟金香等[9]在斑点叉尾鮰(Ietalurus punetaus)和鹿王成志等[10]在俄罗斯鲟(Acipenser gueldenstaedti)上的研究表明,随着饥饿程度的加深,水产动物的肥满度、肝体比和脏体比会显著下降,本实验得到相近的实验结果,鳜的脏体比在饥饿6d时显著下降,而肝体比在饥饿9d时才显著下降,推测可能是鳜饥饿过程中先动用非肝脏的其他内脏,如肠系膜脂等储备的能量,当消耗到一定程度后,才开始动用肝脏的能量储备。
鱼类体内的三种主要营养物质(蛋白质、脂肪和碳水化合物),在饥饿或食物不足时,鱼体代谢内源性营养物质以维持生命活动[11]。一般鱼类的主要贮能物质为脂肪和糖原,只有当长期饥饿状态下才会将蛋白质作为能量代谢[12]。在本实验中,肌肉的水分随饥饿天数的增加显著性上升,而肌肉粗脂肪和粗蛋白则随饥饿时间延长显著下降,在瓦氏黄颡鱼(Pelteobagrus vachelli)[13]上有相同的结果,本研究中鳜肌肉粗蛋白随饥饿时间延长逐渐下降,而肌肉粗脂肪在饥饿3d时即下降44.22%,说明在饥饿过程中,脂肪首先被鳜动用来维持基本生命活动,当脂肪含量过低时就开始消耗肌肉蛋白供能。
3.2 低水温下短期饥饿对鳜血液生化指标的影响
本实验中鳜血糖含量升高之后保持着高水平血糖,与草鱼(Ctenopharyngodon idellus)在饥饿状态下血糖降低后维持在较低水平不同[14],其中血糖的上升幅度和甘油三酯下降幅度几乎一致,推测原因是鳜血液的糖脂代谢转换,血液中甘油三酯转换成血糖。血液中谷草转氨酶和谷丙转氨酶活性的变化可以反映鱼体内肝细胞损伤的程度,肝细胞损伤后,细胞膜的通透性增加,这两种酶能从肝细胞进入血液,导致这两种酶在血液中的活性增加[15]。本实验中的谷草、谷丙转氨酶随饥饿时间延长先下降后上升,说明短时间的饥饿能改善鳜肝功能,但饥饿时间过长,会加剧肝损伤,同时碱性磷酸酶活性也会不断下降,免疫力减弱。
鱼体饥饿期间脂肪大量分解供能,主要通过提高脂肪分解酶活性、加速脂肪在体内的氧化和释放脂肪酸三种途径来实现[14]。在本实验中,血浆中甘油三酯、总胆固醇和高、低密度脂蛋白胆固醇含量在饥饿前3d上升,之后开始明显下降,说明鳜在面临饥饿胁迫时,前3d并不会分解血液中的酯类,主要是消耗肌肉中的粗脂肪,3d后鳜开始分解血液中的总胆固醇和甘油三酯,而高、低密度脂蛋白胆固醇作为总胆固醇和甘油三酯的在机体内的运载工具,也随它们含量的改变发生相应的改变。此外,可以发现,较甘油三酯含量的下降幅度,总胆固醇含量下降并不多。这是因为血浆中甘油三酯含量主要依赖于外源性甘油三酯的吸收,但动物体的几乎所有组织都能合成总胆固醇,饥饿或禁食使肝脏中胆固醇合成大幅度下降,但肝外组织中的合成量减少不多[16]。
3.3 低水温下短期饥饿对鳜胃肠消化能力的影响
鳜胃由黏膜层、黏膜下层、固有层和肌层组成,其中在黏膜层基部中的胃腺细胞,能够同时分泌胃蛋白酶原和胃酸,属于泌酸胃酶细胞。胃蛋白酶原是胃蛋白酶的无活性前体物质,需要在胃酸的作用下激活为有活性的胃蛋白酶。胃H+-K+-ATP酶,也称胃质子泵,是胃蛋白酶原激活过程中分泌胃酸的关键性酶[17]。胃蛋白酶和胃H+-K+-ATP酶对于鱼体的胃部消化起着重要的作用。在本实验中,鳜随着饥饿时间的延长,胃蛋白酶和胃H+-K+-ATP酶分别比初始时下降39%和88%,饥饿使胃的泌酸和泌酶的能力降低,其中胃H+-K+-ATP酶相较于胃蛋白酶下降幅度较大,推测可能是由于饥饿的反馈性抑制,从而也对胃黏膜起到一定的保护作用。
鱼类在遭遇饥饿胁迫过程中,通过调节鱼体各种酶的活性来提高机体储能物质的利用效率,根据鱼体各种酶的活力变化可分析鱼体自身代谢情况和营养状况,但饥饿对鱼体消化酶活性的影响会随鱼的种类、饥饿程度、规格大小不同而存在一定的差异[18]。在本实验中,随着饥饿时间的延长,鳜肠和幽门盲囊消化器官的各类消化酶活性均呈不同程度的下降,其中胰蛋白酶和脂肪酶比淀粉酶下降迅速,可能是鳜作为肉食性鱼类主要利用动物蛋白,对碳水化合物利用能力较差。此外,幽门盲囊的脂肪酶、消化酶和淀粉酶活性等条件下均高于肠道,说明幽门盲囊也是鳜非常重要的消化器官,其地位不亚于肠道。
3.4 低水温下短期饥饿对鳜抗氧化能力的影响
外界环境的变化,例如饥饿会引起水产动物体内抗氧化防御系统发生变化,从而产生大量的自由基,这些自由基能够加速老化,加大局部缺血性损伤,导致氧化应激反应[19]。而SOD和T-AOC是动物体酶类抗氧化系统的重要组成部分,能够清除体内多余自由基,保护自身免受氧化损伤。在生物体内,脂质和自由基发生过氧化反应,其最终产物便是丙二醛,具有很强的毒性[20]。在本实验中,鳜SOD活性在饥饿3d时达到最大值,然后逐渐下降,T-AOC在饥饿6d时达到最大值,然后逐渐下降,而MDA含量在饥饿6d时下降到最小值,然后逐渐上升。本课题组在大口黑鲈(Micropterus salmoides)的饥饿实验当中发现了相似的趋势,而董学兴等[21]在异育银鲫(Carassius auratus gibelio)中发现饥饿8d后SOD活性显著下降,可见饥饿对不同条件,不同鱼种的鱼类的肝脏抗氧化能力影响不同。另外,在本实验中,鳜鳃的抗氧化酶随饥饿时间延长有一定下降,但总体上看,下降不明显,说明饥饿对鳜鳃抗氧化能力影响较弱。此外,Na+-K+-ATP 酶是鳃组织泌氯细胞及细胞器的膜上存在的一种蛋白酶,在鱼体渗透调节过程中起着非常重要的作用[22]。在本实验中,鳜鳃Na+-K+-ATP酶随饥饿时间延长有下降趋势,这与在虹鳟[23]上的研究结果相似。
4 结论
在本实验条件下,鳜15d的短期饥饿会出现体质量下降、体型变瘦和肝脏相对变小现象;饥饿过程中前3d优先利用肌肉脂肪供能,后期优先利用肌肉蛋白质供能;适宜饥饿时间(6d)能改善鳜抗氧化能力,但饥饿时间过长时,会严重影响鳜的消化和抗氧化能力。
