摄食水平对大鲵幼体游泳能力和力竭运动后代谢的影响
2023-11-22侯起淼付世建黄悌基李秀明
侯起淼 付世建 黄悌基 李秀明
(重庆师范大学进化生理与行为学实验室,重庆市动物生物学重点实验室,重庆 401331)
由于自然界中食物资源的时空异质性及种间和种内竞争等因素影响,动物并不能随时获得充足的食物达到饱足。在不同摄食水平下动物往往表现出明显的生理、生态和行为差异,这些表型适应策略对其生存具有重要的生态学意义[1,2]。摄食和运动是保证动物存活和繁衍的两种重要生理活动,在其生活史过程中经常同时发生。由于消化系统和运动系统功能的发挥都依赖于动物呼吸和循环系统对氧气和能源物质的运输,因此摄食活动对动物的运动功能往往具有显著的影响,其效果可能与其摄食水平大小和摄食习性密切相关[3—6]。
运动能力与动物日常觅食、逃避捕食者和求偶繁殖等生理活动密切相关,是其生长、生存和种群繁衍的重要保障。感应流速(Induced velocity)能够一定程度反映水生动物的游泳特性,是其趋流性研究的重要指标之一[7—9]。暴发游泳(Burst swimming)是一种持续时间较短的高耗能运动方式,与水生动物伏击捕食、逃避天敌和穿越激流等活动密切相关[10,11]。此外,水生动物在无氧运动后经常会达到力竭状态,此后的耗氧率会显著高于运动前水平,称为力竭运动后过量耗氧(Excess post-exercise oxygen consumption,EPOC)[12,13]。由于EPOC相关参数具有重要的生理生态学意义,因此也受到研究者的广泛关注[1,10]。比如力竭运动后代谢峰值和过量耗氧分别与动物的最大有氧代谢能力和无氧代谢能力密切相关[1,14]。EPOC持续时间能够反映出动物在无氧运动后的恢复速度,直接影响着无氧运动的再次进行[15,16]。水生动物的运动功能除了受到各种外在环境因素的影响,还可能与自身因素相关,比如个体大小和饥饿过程中营养状态等[17—19]。以往有关摄食水平影响水生动物运动功能的研究主要采用鱼类作为研究对象[1,6,20],两栖动物的相关研究还未见报道。
大鲵(Andrias davidianus)隶属于两栖纲(Amphibia),有尾目(Caudata),隐鳃鲵科(Cryptobrachidae),大鲵属(Andrias),是我国二级保护动物,喜欢生活在溪流环境中。大鲵幼体生活在水中,用外鳃呼吸,通过四肢在水底爬行,趾间有蹼,尾部侧扁,也适应在水中游动[21]。由于栖息地丧失和野生个体不可持续的过度开发,该物种野生种群数量下降严重[22]。目前,有关大鲵的研究主要涉及其繁殖发育[23]、养殖技术[24]、饲料营养[25]、种群生态[26]、生理生化[27]和遗传保护[28]等方面。但是有关大鲵运动生理及其影响因素的研究还鲜有报道。本研究以大鲵幼体为实验对象,考察不同摄食水平对大鲵幼体游泳能力和EPOC的影响,探讨其在不同营养状态下的运动生理适应策略,丰富两栖动物生理生态学资料,为大鲵野外栖息地保护和增殖放流实践提供一定的参考数据。
1 材料与方法
1.1 实验动物和摄食驯化
从湖南省临武县珠江源大鲵养殖合作社购买获得大鲵幼体,养殖在室内循环水养殖水槽中。养殖水体溶氧和温度分别控制在(7.0±0.5) mg/L和(20.0±0.5)℃,光照周期设置为10D∶14L。用摇蚊幼虫作为饵料每两天饱足喂养1次,及时清理残饵和粪便,并适当更换养殖水体以保持水体清洁。驯养周期为4周。
挑选大小相似的大鲵幼体80尾(表1),随机分为4个组,即对照组、1%摄食水平组(摄入1%体重的饵料)、4%摄食水平组(摄入4%体重的饵料)和10%摄食水平组(摄入10%体重的饵料),每组20尾(其中10尾用于游泳能力测定,另外10尾用于力竭运动后代谢测定)。为了使大鲵幼体在实验环境中更好地摄食,将单独的个体放置于自制的“循环密闭式代谢测定仪”(150 mL,图 1)中,每天饱足投喂摇蚊幼虫进行摄食驯化1周。
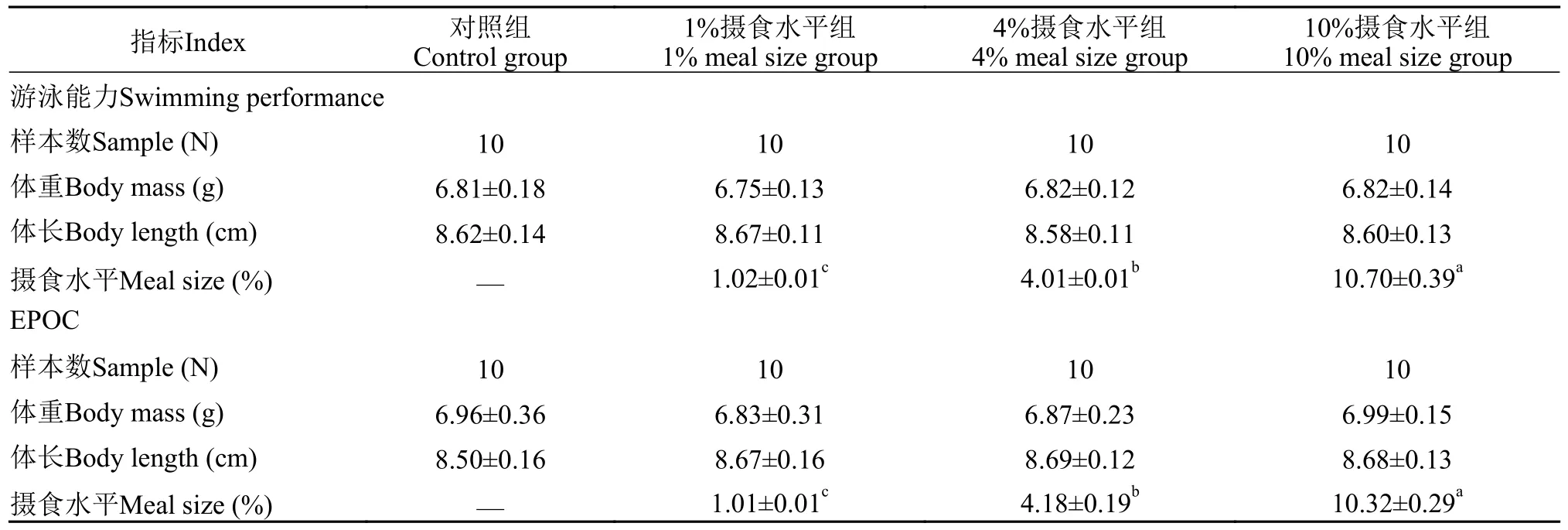
表1 大鲵幼体体重、体长和摄食水平(平均值±标准误)Tab.1 The body mass,body length and meal size of juvenile Chinese giant salamander (Andrias davidianus) in the present study(mean±SE)
1.2 实验方法
感应流速和暴发游泳速度测定摄食驯化周期结束后,在“循环密闭式代谢测定仪”中根据大鲵幼体体重分别投喂相应摄食水平(1%、4%或10%体重)的摇蚊幼虫,对照组没有投喂食物,3h后将其转移到“水生动物游泳代谢测定仪”[29](3.5 L)中适应1h,适应期间水流速度为4 cm/s (约0.5倍体长/秒,bl/s),随后测量其感应流速和暴发游泳速度[4,10,30]。此时为大鲵幼体摄食后4h,其摄食代谢率到达峰值[31]。
“水生动物游泳代谢测定仪”的水流速度由连接的电脑程序控制。在适应期结束以后,水流速度以0.167 cm/s2的加速度持续增加,当大鲵幼体表现出身体离开水底、四肢紧贴身体两侧且尾部左右摆动的游泳状态时,此时的水流速度记为该个体的绝对感应流速(Absolute induction velocity),将绝对感应流速除以大鲵幼体体长记为该个体的相对感应流速(Relative induction velocity)。随着水流速度的进一步增大,大鲵幼体将表现出力竭状态(停留在游泳管末端筛板20s以上不再继续游泳),此时是水流速度记为该个体的绝对暴发游泳速度(Absolute burst swimming speed),将绝对暴发游泳速度除以大鲵幼体体长记为该个体的相对暴发游泳速度(Relative burst swimming speed)[10]。
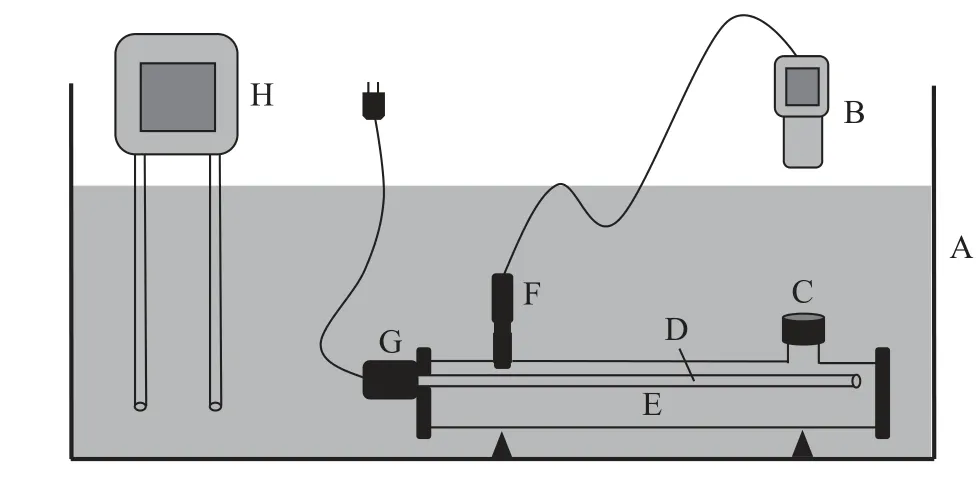
图1 循环密闭式代谢测定仪模式图Fig.1 Circulatory closed respirometer
力竭运动前后代谢测定本实验中大鲵幼体的代谢率在自制的“循环密闭式代谢测定仪”中采用溶氧测定仪(HQ30,Hach Company)测定。代谢率测定前,大鲵幼体单独放置在“循环密闭式代谢测定仪”呼吸管中禁食48h,随后每10s测定1个溶氧值,通过1min内呼吸管内溶氧值随时间变化的斜率计算1个耗氧值 [mg O2/(kg·h)],连续测定30min(30个耗氧值),平均值作为大鲵幼体静止代谢率。将大鲵幼体取出,用同样方法连续测定10min空白呼吸管(细菌耗氧)的溶氧值,平均值作为空白呼吸管斜率。大鲵幼体代谢率通过以下公式计算:
式中,MO2[mg O2/(kg·h)]为大鲵幼体代谢率,St为1min期间溶氧值随时间变化的斜率,S0为空白呼吸管(细菌耗氧)的溶氧值变化斜率,V为代谢测定仪体积(0.15 L),60为60min。W表示大鲵幼体的体重(g)。
在静止代谢率测定结束后,在“循环密闭式代谢测定仪”呼吸管中投喂大鲵幼体相应摄食水平(1%、4%或10%体重)的摇蚊幼虫,对照组不投喂食物,4h后测定其代谢率。连续测定20min(20个耗氧值),平均值作为大鲵幼体的力竭运动前代谢率。此时,1%、4%或10%摄食水平组代谢率为其摄食代谢峰值。随后将大鲵幼体从呼吸管中取出放入环形力竭运动装置(水深10 cm,直径50 cm)中,通过戴手套手指触碰大鲵幼体尾部驱赶其快速持续运动,致使其2min内达到力竭状态,其判断标准为对手指触碰刺激无明显应激反应。随后立即将大鲵幼体放回呼吸管中测定力竭运动后耗氧率,连续测定60min(计算60个耗氧值)。
EPOC相关参数包括静止代谢率(Rest metabolic rate)、运动前代谢率(Pre-exercise metabolic rate)、运动后代谢峰值(Peak post-exercise metabolic rate)、峰值比率(Factorial scope)、代谢率增量(Increment of metabolic rate)、运动后恢复时间(Duration)和过量耗氧(EPOC magnitude)。各个参数具体计算方法参见相关参考文献[1,14]。
1.3 统计分析
本实验所有数据用平均值±标准误(mean±SE)表示,运用Excel 2003进行常规计算后采用SPSS 17.0软件进行统计分析。摄食水平对所有参数的影响采用单因素方差分析(ANOVA),若差异显著则进行多重比较(LSD法),显著性水平规定为P<0.05。
2 结果
2.1 感应流速和暴发游泳速度
摄食水平对大鲵幼体绝对感应流速(F3,39=0.340,P=0.797)、相对感应流速(F3,39=0.413,P=0.745)、绝对暴发游泳速度(F3,39=0.342,P=0.795)和相对暴发游泳速度(F3,39=0.380,P=0.768)都没有显著性影响(表2)。大鲵幼体绝对感应流速和相对感应流速平均值分别为21.94 cm/s和2.56 bl/s,大鲵幼体绝对暴发游泳速度和相对暴发游泳速度平均值分别为33.50 cm/s和3.90 bl/s(图2和图 3)。
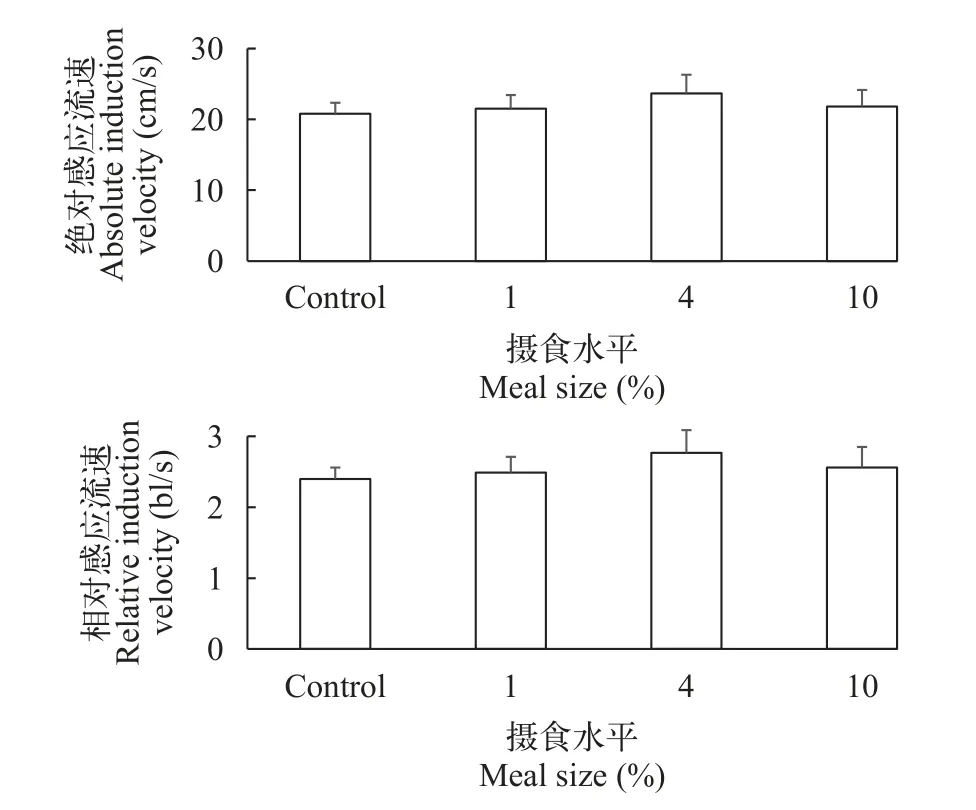
图2 摄食水平对大鲵幼体感应流速的影响(平均值±标准误,样本数=10)Fig.2 The effect of meal size on induction velocity in juvenile Chinese giant salamander (Andrias davidianus;mean±SE,n=10)
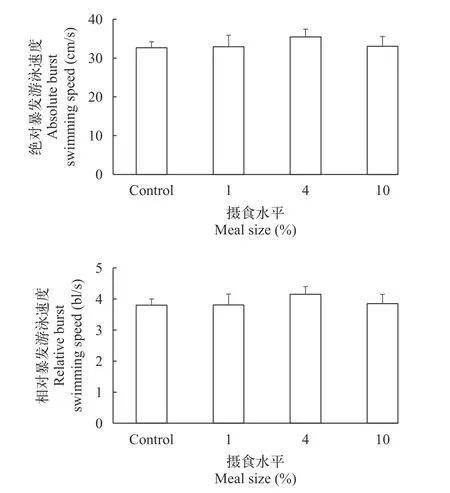
图3 摄食水平对大鲵幼体暴发游泳速度的影响(平均值±标准误,样本数=10)Fig.3 The effect of meal size on burst swimming speed in juvenile Chinese giant salamander (Andrias davidianus;mean±SE,n=10)
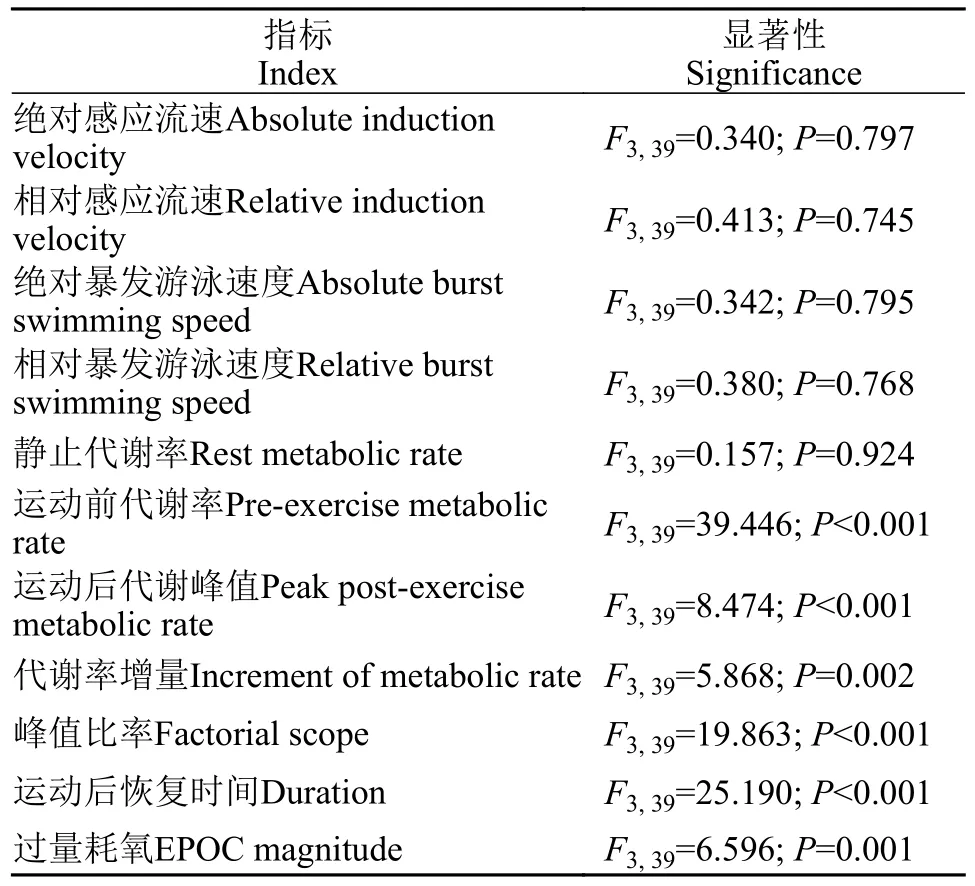
表2 摄食水平对大鲵幼体游泳能力和力竭运动后代谢参数的单因素方差统计分析表Tab.2 The effect of meal size on several variables related to swimming performance and EPOC in juvenile Chinese giant salamander (Andrias davidianus) based on the results of One-way analysis of variance (ANOVA)
2.2 力竭运动后代谢
所有组大鲵幼体在力竭运动后,其代谢率都快速升高达到峰值,随后逐步降低至力竭运动前代谢水平(图4)。所有组大鲵幼体的静止代谢率之间都无显著性差异(F3,39=0.157;P=0.924;表 2和图 5)。随着摄食水平的增加,大鲵幼体运动前代谢率都显著增加(F3,39=39.446;P<0.001),4%摄食水平组运动前代谢率显著高于1%摄食水平组和对照组,但是显著低于10%摄食水平组(表2和图 5)。4%和10%摄食水平组运动后代谢峰值无显著性差异,但是显著高于对照组和1%摄食水平组(F3,39=8.474;P<0.001;表 2和图 5)。4%摄食水平组代谢率增量显著低于对照组,但是显著高于10%摄食水平组(F3,39=5.868;P=0.002;表 2和图 5)。1%、4%和10%摄食水平组峰值比率都显著低于对照组,4%和10%摄食水平组峰值比率之间无显著性差异(F3,39=19.863;P<0.001;表 2和图 5)。4%摄食水平组运动后恢复时间显著小于对照组和1%摄食水平组,但是显著大于10%摄食水平组(F3,39=25.190;P<0.001;表 2和图 5)。4%摄食水平组过量耗氧显著低于对照组,但是显著高于10%摄食水平组(F3,39=6.596;P=0.001;表 2和图 5)。

图4 摄食水平对大鲵幼体力竭运动后代谢反应的影响 (样本数=10)Fig.4 The effect of meal size on excess post-exercise oxygen consumption (EPOC) response in juvenile Chinese giant salamander(Andrias davidianus;n=10)
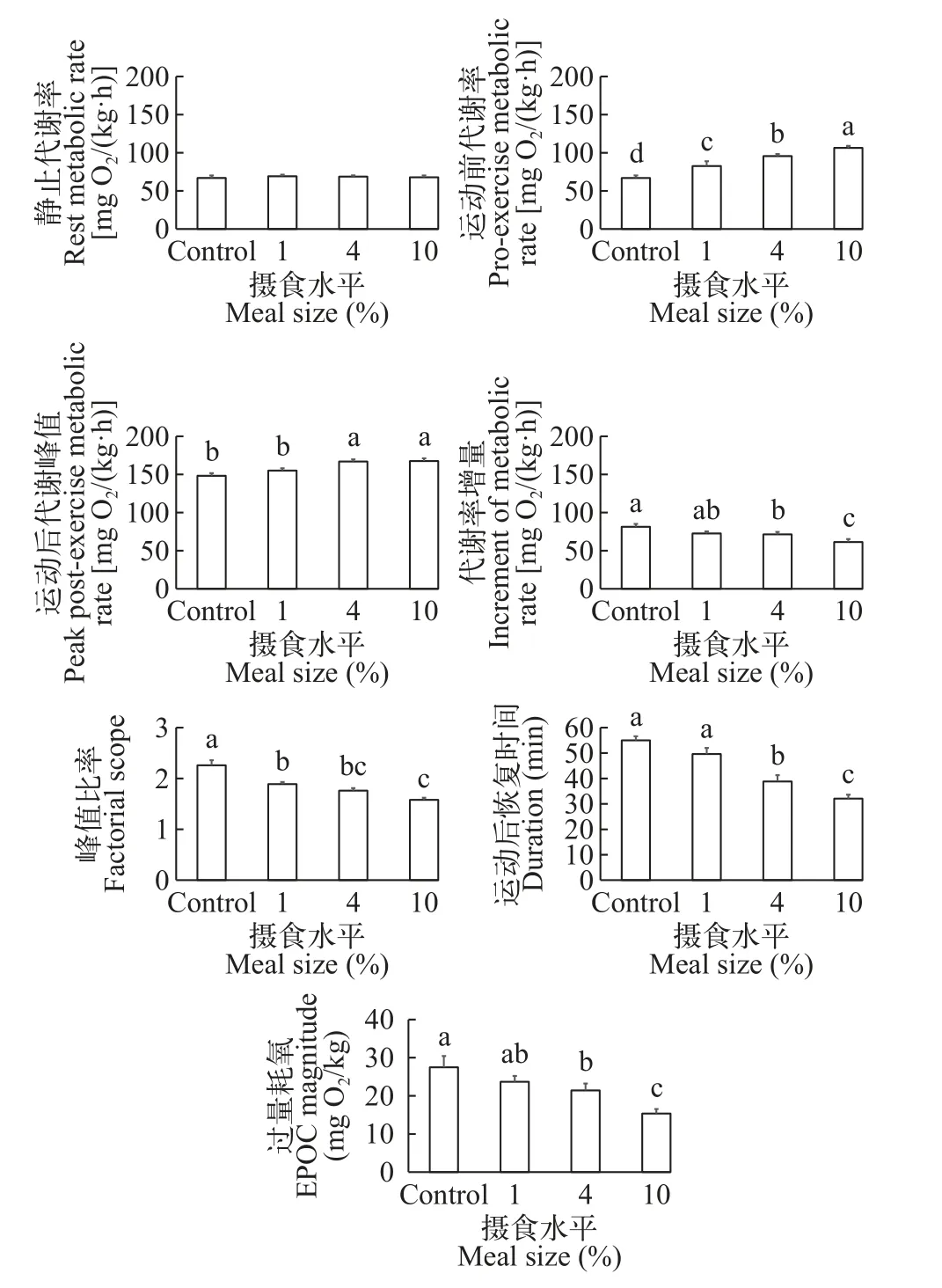
图5 摄食水平对大鲵幼体力竭运动后代谢参数的影响(平均值±标准误,样本数=10)Fig.5 The effects of meal size on several parameters of excess post-exercise oxygen consumption (EPOC) response in juvenile Chinese giant salamander (Andrias davidianus;mean±SE,n=10)
3 讨论
3.1 摄食水平对大鲵幼体感应流速和暴发游泳速度的影响
水生动物一般具有逆流游泳的习性,在一定流速下能够自发产生逆向水流运动的行为反应,此时的流速称为感应流速,是水利设施修建和物种保护的重要参考数据[7,30]。以往研究主要对大鲵游泳行为进行定性描述,但其游泳特征的定量研究还未见报道[21]。本研究发现,大鲵幼体在水流速度约为21.9 cm/s(约2.6 bl/s)时才表现出明显的自发游泳行为,并且摄食水平并未对大鲵幼体的感应流速产生显著性影响,其内在原因还有待进一步探究。在大鲵栖息地保护及增殖放流生境流速选择等实践中,大鲵幼体的这些游泳特征应当予以考虑。
两栖动物幼体阶段主要生活在水中,其游泳能力对其觅食和避敌活动具有重要的意义,因此受到研究者的广泛关注[19,32—34]。以往的研究发现黑斑侧褶蛙(Pelophylax nigromaculatus)、灰树蛙(Hyla chrysoscelis)和南方豹蛙(Rana sphenocephala)蝌蚪的最大游泳速度分别约为7、12和15 cm/s (分别约16、17和15 bl/s)[32,35]。而东方蝾螈(Cynops orientalis)幼体的最大游泳速度约为25 cm/s (约6 bl/s)[34]。本研究发现,大鲵幼体绝对暴发游泳速度约为33 cm/s(约4 bl/s)。由此表明,相比于无尾两栖类动物幼体而言,有尾两栖类幼体具有较高的绝对游泳能力和较低的相对游泳能力,这可能与其相对较长的体长有关。
以往研究发现,南方鲇(Silurus meridionlis)的临界游泳速度(Critical swimming speeds)在低摄食水平(2%—8%)下没有显著变化,但在较高摄食水平(12%—20%)下显著下降。由于南方鲇幼鱼在12%—20%摄食水平下的摄食代谢峰值约为其静止代谢率的4.7倍,研究者认为高摄食水平下游泳能力的降低可能与其较高的消化能力相关[2]。不过本研究发现,大鲵幼体暴发游泳速度并没有受到摄食水平(1%—10%体重)的显著影响,暗示其消化功能并未对运动能力产生显著的影响,这可能与其相对较低的消化代谢能力有关(摄食代谢峰值仅为其静止代谢率的2.2倍)[31]。这种结果与草鱼(Ctenopharyngodon idellus)和瓦氏黄颡鱼(Pelteobagrus vachelli)的研究结果相似,这两种鱼类在1%—8%摄食水平下其游泳速度也并没有显著变化,其最大摄食代谢峰值分别为其静止代谢的1.5和2.2倍[2]。大鲵幼体的游泳能力在不同摄食水平下的“维持”策略可能有利于继续捕获猎物和逃避捕食者,提高其复杂溪流生境下的生存适合度。
3.2 摄食水平对大鲵幼体力竭运动后代谢特征的影响
由于动物呼吸系统和循环系统功能的限制,动物往往具有一个潜在的最大有氧代谢率,它与其静止代谢率之间的差值即为动物执行各项生理活动所能利用的有效代谢空间[36—38]。因此,当消化和运动功能同时进行时,可能出现两种不同的代谢模式: 一种是“添加模式”,动物的有效代谢空间足够大,能够同时满足其消化和运动功能;另一种是“竞争模式”,动物的有效代谢空间不足,优先维持其中一种生理功能而牺牲另一种生理功能[4,5,39]。以往研究发现,一些水生动物在禁食状态下诱导出的代谢率与消化期间运动诱导出的代谢率相当,表明单一的运动能够诱导出最大有氧代谢,当摄食和运动功能同时进行时可能采用代谢“竞争模式”,即优先维持一种生理功能,例如中华倒刺鲃(Spinibarbus sinensis)、岩原鲤(Procypris rabaudi)和草鱼[1,6,40]。本研究发现,大鲵幼体10%摄食水平组力竭运动后的代谢峰值[即消化期间运动诱导的最大有氧代谢,约为167.63 mg O2/(kg·h)]显著高于对照组力竭运动代谢峰值[即单一运动诱导的最大有氧代谢,约为148.21 mg O2/(kg·h)]和10%摄食水平组力竭运动前代谢峰值[即单一摄食消化诱导的最大有氧代谢,106.32 mg O2/(kg·h)]。由此表明,单独的消化或者运动生理活动均不能诱导出大鲵幼体的最大耗氧率,当摄食和运动功能同时进行时可能出现一定程度的代谢“添加模式”。这一结果与印度蟒蛇(Python molurus)、南方鲇、瓦氏黄颡鱼和锦鲫(Carassius auratus)的研究相似[1,41]。
动物的各项生理功能在力竭运动后恢复的快慢直接影响着下一次力竭运动的进行,对其捕食和避敌活动具有重要的生态学意义。以往研究发现,水生动物力竭运动后恢复时间通常依赖于摄食水平的高低[1]。低摄食水平对其力竭运动后恢复时间没有显著影响,在高摄食水平下恢复时间通常会缩短,例如南方鲇、岩原鲤、锦鲫和青鱼(Mylopharyngodon piceus)[1,6,20]。与以往研究结果相似,本研究发现,1%摄食水平组大鲵幼体力竭运动后恢复时间与对照组无显著差异,但是在4%和10%摄食水平下力竭运动后恢复时间显著缩短。力竭运动后恢复时间是指动物力竭运动恢复过程中代谢率回落到与运动前代谢率无显著性差异所对应的时间。由于高摄食水平组的力竭运动前代谢较高(即摄食代谢峰值较高),所以力竭运动后恢复到这个较高运动前代谢率所需时间可能相应缩短。因此,大鲵幼体在较高摄食水平下(4%和10%)力竭运动后恢复时间显著缩短可能与其力竭运动前代谢相对较高有关[1]。从力竭运动中快速恢复的代谢特征将有利于大鲵幼体下一次捕食和避敌等力竭运动的进行。
EPOC大小是评价动物无氧代谢能力的重要指标之一,其大小受到各种因素的影响[1,14,40,42,43]。摄食对动物EPOC大小的影响可能与其营养等级和摄食习性有关。有研究发现,一些营养等级较低且频繁摄食的草食或杂食性种类(如草鱼和岩原鲤),其EPOC大小并未受到摄食活动的影响,研究者认为摄食后无氧代谢能力的保持将有利于频繁的觅食和避敌活动[1,6]。本研究发现,大鲵幼体在1%摄食水平下的EPOC大小与对照组无显著差异,但是在4%和10%摄食水平下的EPOC大小显著低于对照组。由此表明,大量食物的摄入对大鲵幼体的无氧代谢能力产生了负面影响。不过,这并没有导致大鲵幼体暴发游泳速度的显著下降。这种无氧代谢能力和无氧运动能力非正相关的现象在鱼类相关研究中也有发现[10]。EPOC是一个复杂的生理过程,其内在影响因素众多,比如磷酸肌酸和糖原等能量物质的分解与恢复、乳酸等代谢废物的积累与清除以及渗透平衡的调节等[44—46],EPOC大小与无氧运动能力之间的关联还有待进一步研究。由于在摄食消化期间大鲵幼体的游泳能力并未下降,加之其具有躲藏消化的习性,因此无氧代谢能力的下降可能对其生存适合度的影响有限。这种现象在一些营养等级较高,具有伏击捕食、隐匿消化习性的肉食性种类中也有发现,比如南方鲇和鲇(Silurus asotus)[1,42]。
综上所述,随着摄食水平的增加,大鲵幼体的感应流速和暴发游泳速度没有显著变化,但却表现出有氧代谢能力的增加、恢复速率的加快和无氧代谢能力的降低等能量代谢特征,这可能与其较低的消化代谢能力和隐匿消化的习性有关,有助于在复杂的溪流生境中进行捕食和避敌活动。
