广西外来入侵植物豚草的分布调查
2023-11-20黄小荣何峰彭玉华陈始贵
黄小荣 何峰 彭玉华 陈始贵
黄小荣,何 峰,彭玉华,等. 广西外来入侵植物豚草的分布调查[J]. 杂草学报,2023,41(2):29-34.
doi:10.19588/j.issn.1003-935X.2023.02.0004
摘要:2019—2022年,对广西111个县(市、区)的豚草分布进行样带调查。结果表明,横州、兴宾、上林、武宣、象州、鹿寨、宾阳、灌阳和隆林9个县(市、区)有豚草发生;除了在兴宾区持续蔓延之外,横州市和上林县成为豚草的新增暴发热点,新热点的传播风险更大。装卸场是豚草发生率和归化率最高的生境,车辆传播和污染泥土传播是广西豚草扩散的主要渠道。环境的物种丰富度与豚草盖度呈负相关,但与豚草发生率和归化率均无显著关联。桂中平原周边的崇山峻岭对豚草传播起到隔离带作用。南部沿海地区的北海、钦州和防城港均未发现豚草,这种情况可能与冬季气候温和有关。
关键词:豚草;外来入侵物种;广西;分布;春化
中图分类号:S451;Q948.2 文献标志码:A 文章编号:1003-935X(2023)02-0029-06
Distribution of Alien Invasive Species Ambrosia artemisiifolia in Guangxi Area
HUANG Xiao-rong,HE Feng,PENG Yu-hua,CHEN Shi-gui
(Guangxi Academy of Forestry Sciences,Nanning 530002,China)
Abstract:A field survey was conducted in 111 counties of Guangxi area to investigate distribution of Ambrosia artemisiifolia. The results showed that A. artemisiifolia occurred in 9 counties,including Hengzhou,Xingbin,Shanglin,Wuxuan,Xiangzhou,Luzhai,Binyang,Guanyang and Longlin. In addition to the continued spread in Xingbin County,Hengzhou and Shanglin became the other two new outbreak hotspots of A. artemisiifolia,and the risk of transmission in the new hotspots was even greater. Loading yard was the habitat with the highest occurrence rate and naturalization rate of A. artemisiifolia,and vehicle and contaminated soil transport were the main channels of spread for A. artemisiifolia in Guangxi area. Species richness in the environment was negatively correlated with the coverage of A. artemisiifolia,but was not correlated with occurrence rate or naturalization rate. Towering and steep mountains around Guizhong Plain acted as a natural barrier to prevent the spread of A. artemisiifolia. No A. artemisiifolia was found in Beihai,Qinzhou and Fangchenggang of the southern part coastal areas due to warm winter climate there.
Key words:Ambrosia artemisiifolia;alien invasive species;Guangxi;distribution;vernalization
收稿日期:2023-03-07
基金項目:国家林业局生物安全项目(编号:KJZXSA2018012、KJZXSA2019010、KJZXSA202008)。
作者简介:黄小荣(1964—),女,广西富川人,硕士,高级工程师,主要从事外来植物调查与研究。E-mail:huangxr2004@sina.com。
豚草(Ambrosia artemisiifolia L.)为雌雄同株、风媒传粉的一年生草本植物,原产北美洲。近几十年来,豚草在全球迅速传播,已经扩散至80多个国家和地区,其中25个国家将其列为外来入侵物种[1]。在欧洲的豚草严重暴发区,豚草造成作物大量减产,高致敏花粉引发公共卫生问题[2-3]。
豚草20世纪40年代传入我国[4-5],现在在上海、广州[6]、新疆、黑龙江等地区均有分布[7]。豚草是一种喜热植物[8]。Essl等发现,入侵奥地利的豚草主要分布在海拔低于400 m的低海拔地区[9]。据Bassett等报道,在恒温条件下培养28 d的豚草种子萌发率很低(<8.7%),10 ℃ 16 h和30 ℃ 8 h交替温度培养28 d的种子萌发率最高(75%)[10]。这表明豚草在大陆性气候下生长最好[11]。广西大部分内陆地区适合豚草生长发育。豚草于20世纪80年代传入广西来宾市兴宾区[12-14],广西其他地区未见豚草分布的报道。2019—2022年,笔者对广西全区111个县(市、区)进行了林业外来物种普查,分析了广西豚草发生区和归化区的生境特征,可以为清除零星豚草提供信息,有助于防范其蔓延扩散。
1 材料与方法
1.1 样地概况与调查方法
在广西111个县(市、区)进行实地调查,侧重调查桂中地区来宾市兴宾区及相邻的县(市、区),以每个县(市、区)的城区为中心,向东南西北4个方向沿公路行进至县边界,每个方向选3~4个地段作样带(宽10~50 m,长500~1 500 m)进行调查,样带大小以连续行走5 min无新增物种为度,共调查样带250个,样带间距在5 km以上。记录样带所在地理位置和生境特征,以及样带内所有维管植物的种类、丰度、豚草盖度和发生情况。样带内豚草数量大于50株,或零星块状分布出现3次的记为归化;低于该标准的记为偶发或无[15]。公路边的停车装卸点、修车场、散货露天堆场、木楞场等生境类型都归入装卸场。
1.2 豚草识别
豚草植株高5~250 cm,茎直立,上部分枝被糙毛状柔毛;茎下部叶对生,上部互生,2~3回羽状分裂;头状花序单性,雌雄同株;雄性头状花序在枝顶排列成总状花序[16]。雌头状花序2~5组簇生于雄头状花序下部叶腋处[11];总苞闭合,倒卵形,顶端有围裹花柱的圆锥状嘴部,在顶部以下有5~8个尖刺[17],每个总苞只有1朵小花[11];瘦果倒卵形,藏于坚硬的总苞中[17]。豚草每年11月底开始枯死,雄花脱落;次年3月种子萌发;营养生长期的豚草与伴生的青蒿、艾蒿极形似,但豚草叶片被毛、叶背青绿色;6月底开始顶生雄花穗现蕾,辨识变得容易。
1.3 数据处理
数量分析是为了揭示环境因子与豚草分布的关系,因此剔除了未发现豚草的102个县(市、区),只对本次调查发现豚草的9个县(市、区)进行统计分析,共计82个样带。运算采用R 4.1.3的二元广义线性模型(GLM)和混效模型,响应变量为豚草归化时,用二元变量1和0分别代表归化和其他;响应变量为豚草发生时,二元变量的1包括归化和偶发[18]。解释变量包括生境类型、县(市、区)、道路类型、纬度、经度、海拔和物种个数,后4个变量先行对数转换使其在同一数量级、权重相同。
2 结果与分析
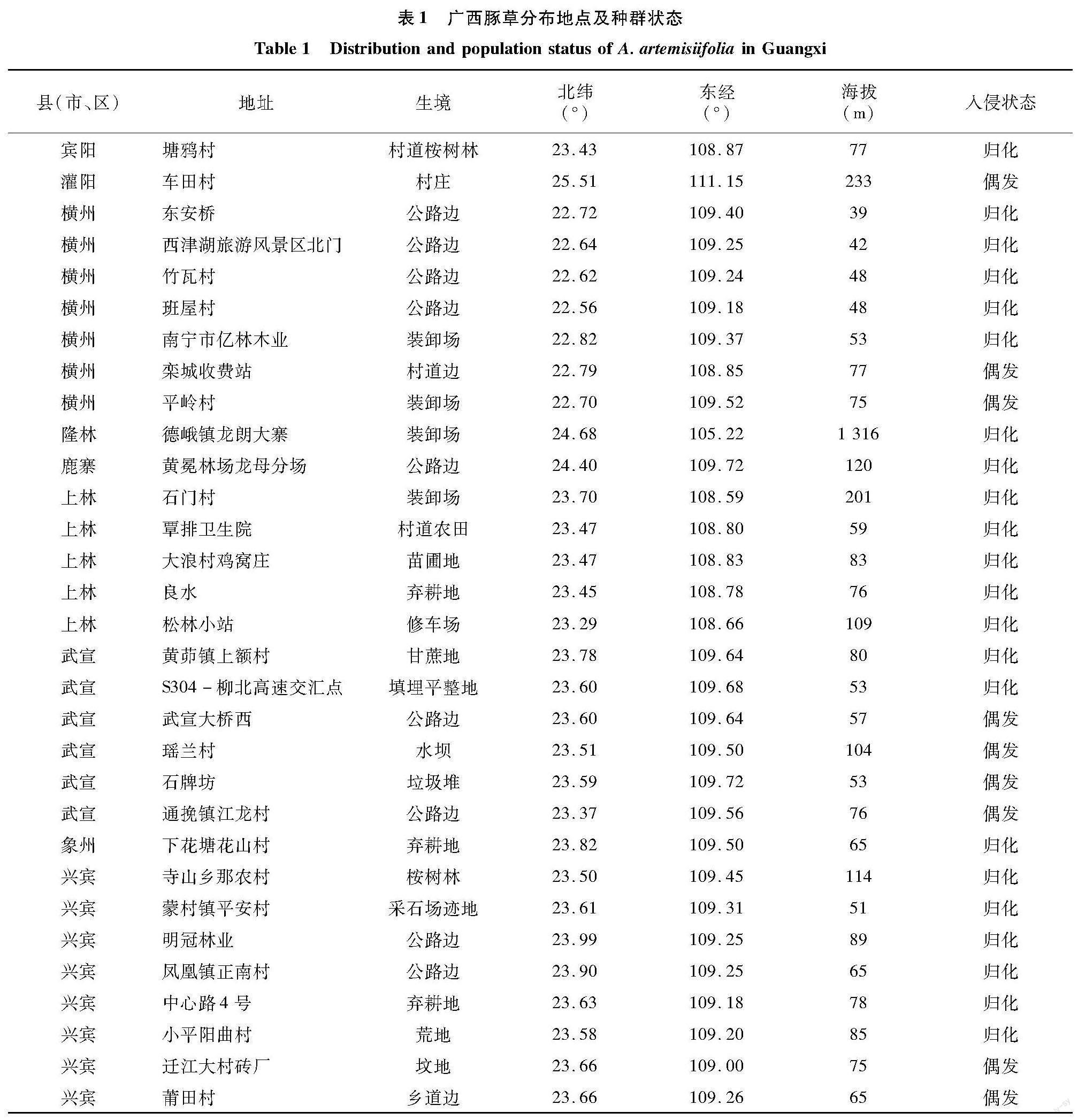
2.1 广西豚草分布地点及种群状态
发现豚草的9个县(市、区),共计82个样带,其中31个样带发现豚草,分布在兴宾、上林、横州、武宣、象州、鹿寨、宾阳、灌阳和隆林9个县(市、区)(表1)。来宾市兴宾区东南部仍为豚草危害重灾区;蒙村镇、寺山乡尤为严重,部分地点豚草盖度超过50%;周边的武宣县、象州县均受影响;兴宾区西部豚草分布较少。上林县和横州市形成了新的豚草暴发热点;上林县大浪村鸡窝庄的一个黄花风铃苗圃豚草生长极为繁茂,豚草盖度达70%,密度约120株/m2;横州市东安桥路边豚草单优群落连片约100 m2,高达2.8 m。本研究发现的大多数豚草种群分布在桂中平原,海拔低于100 m,但在隆林海拔1 316 m的石山区公路边台地上(装卸场)也发现归化的豚草(表1)。
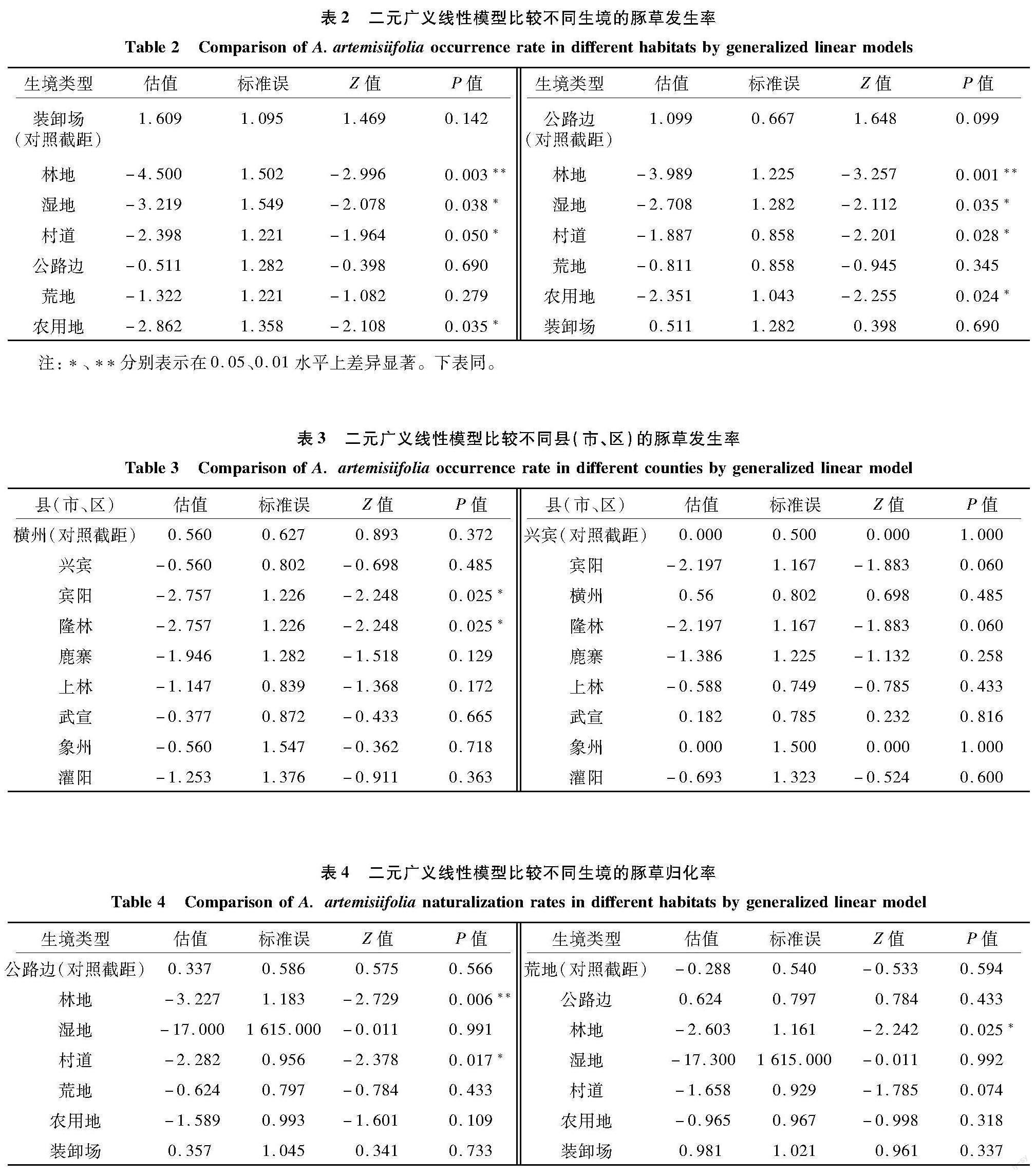
2.2 不同生境及不同区域豚草发生率
9个县(市、区)的82个样地,不同生境的豚草发生率差异显著,豚草发生率分别为装卸场5/6,公路边9/12,荒地8/14,村道5/16,农用地2/9,湿地1/6,林地1/19;装卸场和公路边的豚草发生率显著或极显著高于村道、农用地、湿地和林地,表2是以装卸场和公路边为对照时与其他生境豚草发生率比较结果。横州的豚草发生率(7/11)显著高于宾阳(1/10)和隆林(1/10);武宣(6/11)和兴宾(8/16)的豚草发生率高于宾阳和隆林,但未达0.05显著水平。表3是以横州和兴宾区为对照时不同县(市、区)之间豚草发生率比较结果。
2.3 不同生境豚草归化率
调查结果表明,豚草归化率分别为装卸场4/6,公路边7/12,荒地6/14,农用地2/9,村道2/16,林地1/19,湿地0/6,装卸场归化率最高,装卸场和公路边的豚草归化率极显著高于林地,荒地豚草归化率显著高于林地。表4是以公路边和荒地为对照的不同生境豚草归化率比较结果。象州和横州样地的豚草归化率(1/2、5/11)略高于宾阳和隆林(1/10);其他县(市、区)豚草归化率之间无明显差异。
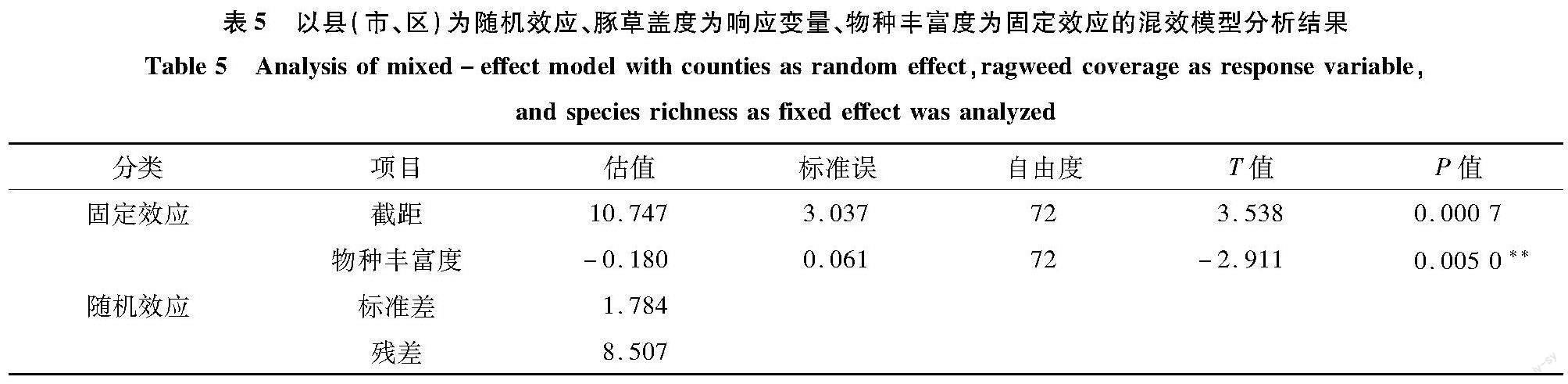
2.4 物种丰富度固定效应的混效模型分析
物种丰富度与豚草盖度呈负相关(r=-0.926);在响应变量为豚草盖度、县(市、区)为随机效应、物種丰富度为固定效应的混效模型(表5)中,物种丰富度的斜率为-0.180,物种丰富度每增加10,豚草盖度降低1.80%。但是,物种丰富度与豚草发生和豚草归化变量均无显著关联。
3 讨论与结论
3.1 豚草主要由人类活动传播
豚草瘦果中央末端突喙可达2 mm[10],顶端有4~7个刺状突起[19],附着力较弱[20];豚草种子较重,中国种群的平均质量为4.65 mg[11],只能靠重力传播[20];大多数种子落在母株附近1 m之内[11];但豚草的土壤种子库持久,被豚草种子污染的土壤可通过车轮实现远距离迁移。在隆林,豚草发生在偏僻的桂西石漠化山区,海拔很高,且远离来宾豚草高发地区;附近的石山是一个造林示范点。豚草种子和幼苗很容易随树苗一起传播[15],林木苗圃很可能是隆林豚草的传播源。
装卸场是豚草发生率和归化率最高的生境。装卸场是货物暂时堆放、受各种运输工具频繁干扰的场所,为豚草种子随车辆远距离传播创造了机会;横州和上林的豚草发生区大都与河沙、泥土、砖瓦等建材和农林收获物的运输有关。有研究认为,豚草传播的主要途径是车辆,轮胎、篷布等携带种子沿交通线路传播[8,20]。
来宾市兴宾区是广西最早报道豚草入侵的地区,20世纪80年代该地区从新西兰引进牧草种子时带入[12];目前,豚草在兴宾区的多个乡镇归化,最大种群出现在兴宾区东南部的寺山乡和蒙村镇。在兴宾区城区及周边5 km内的道路绿化带、公园等多个地点均未发现豚草。
频繁的人类活动干扰和当地豚草管理观念淡薄共同导致横州和上林成为新的豚草暴发热点。横州的豚草发生率(7/11)和归化率(5/11)最高,广西形成了横州、兴宾、上林三大豚草传播中心。豚草新热点比旧热点的传播风险更大,因为民众往往对新热点缺乏防范意识。宾阳县北边毗邻兴宾区,西边紧连上林县;靠近兴宾区的宾阳农户多知晓邻乡有豚草,须严防传入。本研究在宾阳的10个调查样带中有9个没有豚草,只在宾阳塘鸦村Y035乡道旁发现豚草,该乡道直通上林大浪村鸡窝庄;宾阳成功地阻挡了兴宾区方向的传入,但豚草已从上林悄然入境。
3.2 山嶺的物理阻隔作用明显
大多数豚草种群分布在桂中平原,即驾桥岭—大瑶山—莲花山以西、都阳山—大明山以东的地区。在紧邻武宣县、兴宾区的覃塘区、港北区和桂平市,均未发现豚草。莲花山突兀横亘在武宣与港北区之间,对豚草传播起到物理阻隔作用。笔者在大明山西坡的马山县也进行了密集调查,未发现豚草。大明山东坡是新增豚草肆虐地区上林县。崇山高岭往往意味着经济活动和人为干扰较少,限制了豚草随人类活动的扩散。桂北和桂西除灌阳和隆林有零星短暂分布外,其他地方未发现豚草。
3.3 气候限制豚草向桂南沿海地区扩散
广西南部沿海地区北海、钦州、防城港均没有豚草发生,这种情况可能与气候密切相关。豚草种子需要长时间的寒冷春化才能打破休眠。4 ℃是豚草最佳春化温度[21]。-5 ℃春化效果最差,10 ℃处理效果中等,-5 ℃、10 ℃春化种子连续春化15周才能达到最大萌发率,而4 ℃春化种子12周即达到最高萌发率[22]。豚草种子萌发率随着春化天数呈线性增加,4 ℃春化7、14、21、28 d后 25 ℃ 温育催芽的萌发率约为14%、21%、29%、38%[23]。从天气网(www.tianqi.com)查询2012—2022年间冬季3个月的平均低温,北海、钦州和防城港的11年冬季低温均值高于12 ℃,来宾、上林、横州的11年冬季低温均值分别为9.2、9.6、10.2 ℃。沿海地区冬季气候温和,不利于豚草种子打破休眠,限制了豚草在沿海地区的扩展。
目前,笔者发现豚草入侵了广西桂中平原和横县平原的9个县(市、区),形成了横州、兴宾和上林3个暴发热点。鉴于豚草的恶性生态影响和花粉过敏症的危害,在向当地农林部门及时反馈的同时,应加大科普宣传,让当地人们了解该入侵物种的危害性,群防群治,以有效控制其在广西的继续蔓延。
参考文献:
[1]Montagnani C,Gentili R,Smith M,et al. The worldwide spread,success,and impact of ragweed (Ambrosia spp.)[J]. Critical Reviews in Plant Sciences,2017,36:139-178.
[2]Schindler S,Bayliss H R,Essl F,et al. Effectiveness of management interventions for control of invasive common ragweed Ambrosia artemisiifolia:a systematic review protocol [J]. Environmental Evidence,2016,5:11.
[3]Basky Z,Ladányi M,Simonc[DD(-1*2/3]ˇic[DD(-1*2/3]ˇ A. Efficient reduction of biomass,seed and season long pollen production of common ragweed (Ambrosia artemisiifolia L.)[J]. Urban Forestry and Urban Greening,2017,24:134-140.
[4]梁巧玲,陆 平. 新疆伊犁河谷发现外来杂草——三裂叶豚草和豚草[J]. 杂草科学,2014,32(2):38-40.
[5]谢云珍,王玉兵,谭伟福. 广西外来入侵植物[J]. 热带亚热带植物学报,2007,15(2):160-167.
[6 ]周 伟,徐瑞晶,赵 倩,等. 广州市花都区豚草种群监测调查[J]. 杂草科学,2010,28(3):9-13.
[7]邓贞贞,赵相健,赵彩云,等. 繁殖体压力对豚草(Ambrosia artemisiifolia)定殖和种群维持的影响[J]. 生态学杂志,2016,35(6):1511-1515.
[8]Galzina N,Baric[DD(-1*2/3]' K,c[DD(-1*2/3]'epanovic[DD(-1*2/3]' M,et al. Distribution of invasive weed Ambrosia artemisiifolia L. in Croatia[J]. Agriculturae Conspectus Scientificus,2010,75(2):75-81.
[9]Essl F,Dullinger S,Kleinbauer I.Changes in the spatio-temporal patterns and habitat preferences of Ambrosia artemisiifolia during its invasion of Austria[J]. Preslia,2009,81(2):119-133.
[10]Bassett I J,Crompton C W.The biology of Canadian weeds.11. Ambrosia artemisiifolia L. and A. psilostachya DC.[J]. Canadian Journal of Plant Science,1975,55(2):463-476.
[11]Essl F,Biro K,Brandes D,et al. Biological flora of the British Isles:Ambrosia artemisiifolia[J]. Journal of Ecology,2015,103(4):1069-1098.
[12]周忠實,陈红松,郑兴汶,等. 广聚萤叶甲和豚草卷蛾对广西来宾豚草的联合控制作用[J]. 生物安全学报,2011,20(4):267-269.
[13]刘晓亮,何金富,李克敌,等. 豚草卷蛾在广西来宾建群状况及其对作物的安全性调查[J]. 生物安全学报,2013,22(2):136-139.
[14]林春华,唐赛春,韦春强,等. 广西来宾市外来入侵植物的调查研究[J]. 杂草科学,2015,33(1):38-44.
[15]Skálová H,Guo W Y,Wild J,et al. Ambrosia artemisiifolia in the Czech Republic:history of invasion,current distribution and prediction of future spread[J]. Preslia,2017,89(1):1-16.
[16]万方浩,刘全儒,谢 明,等. 生物入侵:中国外来入侵植物图鉴[M]. 北京:科学出版社,2012.
[17]中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京:科学出版社,1993.
[18]Pinke G,Kolejanisz T,Vér A,et al. Drivers of Ambrosia artemisiifolia abundance in arable fields along the Austrian-Hungarian border[J]. Preslia,2019,91(4):369-389.
[19]周业盛,吴永健,超材踞,等. 豚草的扩展规律和综防技术的研究[J]. 杂草科学,1992(4):35-38.[HJ2.3mm]
[20]Vitalos M,Karrer G. Dispersal of Ambrosia artemisiifolia seeds along roads:the contribution of traffic and mowing machines [C]//Pyek P,Pergl J. Biological invasions:towards a synthesis. NeoBiota,2009,8:53-60.
[21]Baskin J M,Baskin C C.Ecophysiology of secondary dormancy in seeds of Ambrosia artemisiifolia[J]. Ecology,1980,61(3):475-480.
[22]Willemsen R W. Effect of stratification temperature and germination temperature on germination and the induction of secondary dormancy in common ragweed seeds[J]. American Journal of Botany,1975,62(1):1-5.
[23]Dinelli G,Marotti I,Catizone P,et al. Germination ecology of Ambrosia artemisiifolia L. and Ambrosia trifida L. biotypes suspected of glyphosate resistance[J]. Central European Journal of Biology,2013,8(3):286-296.
