巨菌草分子研究及生态修复应用的进展
2023-11-15史志丹郭呈宇房永雨伊风艳刘思博皇甫海燕孙红霞丁海君
史志丹,郭呈宇,房永雨,伊风艳,刘思博,皇甫海燕,孙红霞,丁海君
(内蒙古自治区农牧业科学院草原研究所,内蒙古 呼和浩特 010031)
巨菌草(Cenchrus fungigraminus)是单子叶纲禾本科狼尾草属多年生直立丛生型植物,2006 年由福建农林大学菌草研究所从南非夸祖鲁奈塔尔省引入我国进行培育[1],是适宜在热带、亚热带、温带地区生长的高产优质饲草[2]。 在前人的不断探索下引种至北方, 但是由于温度等因素只能是一年生[3]。 目前,在海南、福建、浙江、宁夏、陕西、甘肃、新疆、内蒙古等地均有栽培种植[4]。 巨菌草产量大,且含有丰富的粗蛋白和粗纤维。处在幼嫩期的巨菌草营养丰富,适口性好,是饲草的优良选择。巨菌草植株高大直立、丛生,根系发达,在较深土层中仍发育良好[5],因其吸附雨水和对土壤的防冲能力要强于森林和农作物, 而成为环境保护和修复的最好植物之一[6]。 目前已在生态脆弱的砒砂岩地、滩涂盐碱地、洪积扇等地成功试种,且生物量显著高于当地优势草种[7]。 巨菌草还能吸附土壤重金属,具有修复镉污染土壤的能力,成为一种快速、有效的土壤修复方法[8]。 另有研究表明,巨菌草体内具有丰富的内生菌,可以起到促生作用[9]。 巨菌草作为C4植物,在能源方面也有应用,可以用于发酵,其产生的沼气用于燃烧释放能量,产生的沼渣还可以用于制备乙醇,具有广阔的发展潜力[10]。 随着不可再生资源的减少以及人们环保意识的提升, 清洁生物质资源利用研究和关注度持续增加,由于巨菌草产热高、灰分少、碳元素残留少,巨菌草也被认为是一种新能源作物[9]。巨菌草作为优质饲草、 生态治理材料和能源作物等获得广泛的栽培应用, 并创造了巨大的经济和生态效益。本文从巨菌草遗传分子标记开发,转录组研究及抗逆基因等方面对相关研究及其在生态治理修复方面的应用进行综述, 以期为牧草种质资源的开发与利用、 优异基因的挖掘和牧草分子育种及后续在生态治理中的应用提供参考。
1 遗传分子标记
遗传分子标记是能够明确反映遗传多态性的生物学特征的研究方法,包括形态学标记、生化标记、细胞学标记和分子标记。 形态学标记、细胞学标记、生化标记是基因表达后的标记,属于间接标记, 而分子标记是直接对基因进行标记的一种方法,广泛应用于生物分类和遗传多样性分析。近年来, 一些分子标记技术, 如简单重复序列(simple sequence repeat,SSR)检测技术、单核苷酸多态性(single nucleotide polymorphisms,SNP) 检测技术、相关序列扩增多态性 (sequence-related amplified polymorphism,SRAP)检测技术等被应用在苜蓿[11]、苏 丹 草[12]、狼 尾 草 属[13]等 禾 本 科 牧 草 的 分 类 和 鉴定中。 由于分子标记对个体来说扩增出的特异性DNA 片段具有高度特异性和稳定性的特点,因此可以来评估种质资源、鉴定种质及新品种。简单重复序列间标记(inter-simple sequence repeat,ISSR)技术由于具有简单、多态性丰富、稳定性、分辨率及重复性好等特点已经被广泛应用于植物种质资源遗传多样性评估和亲缘关系鉴定的研究[14]。 朱丹丹[15]对狼尾草属中常用的24 种菌草ISSR 遗传多样性进行研究及采用传统压片法对象草(Pemisetum purpureum)和巨菌草的细胞染色体核型进行分析。 随机扩增多态性DNA 标记(random amplified polymorphic DNA,RAPD) 分子标记也是植物遗传多样性研究的主要技术手段之一, 它是以PCR 为基础的DNA 检测技术,实验易于操作、程序简单快速、花费较少、不受物种限制、具有通用性、多态性丰富等优点,被广泛用于种质资源鉴定、遗传图谱构建、物种分类等研究领域。 冯喜[14]利用形态学、 细胞学、RAPD 和内源转录间隔区(internal transcribed spacer,ITS) 分子标记研究10种禾本科菌草的亲缘关系, 最终确定巨菌草的分类地位。 从分子水平上探讨了狼尾草属植物间的亲缘关系。 根据RPAD 标记改进后的分子标记方法特定序列扩增(sequence characterized amplified regions,SCAR)克服了RAPD 的不稳定性,其重复性好、对实验要求低,操作简单,成为较理想的分子标记工具之一, 尤其适合大量样品的快速检测和分析。 罗宗志[13]对狼尾草属包括巨菌草在内的牧草品种(系)进行核型分析,并成功将RAPD 标记转化为更加稳定可靠的SCAR 标记, 不仅能够鉴定生产实践中的狼尾草属品种(系),还能够从分子水平上对狼尾草属品种(系)种质资源的遗传多样性进行系统分析。 常用的分子标记如表1所示。
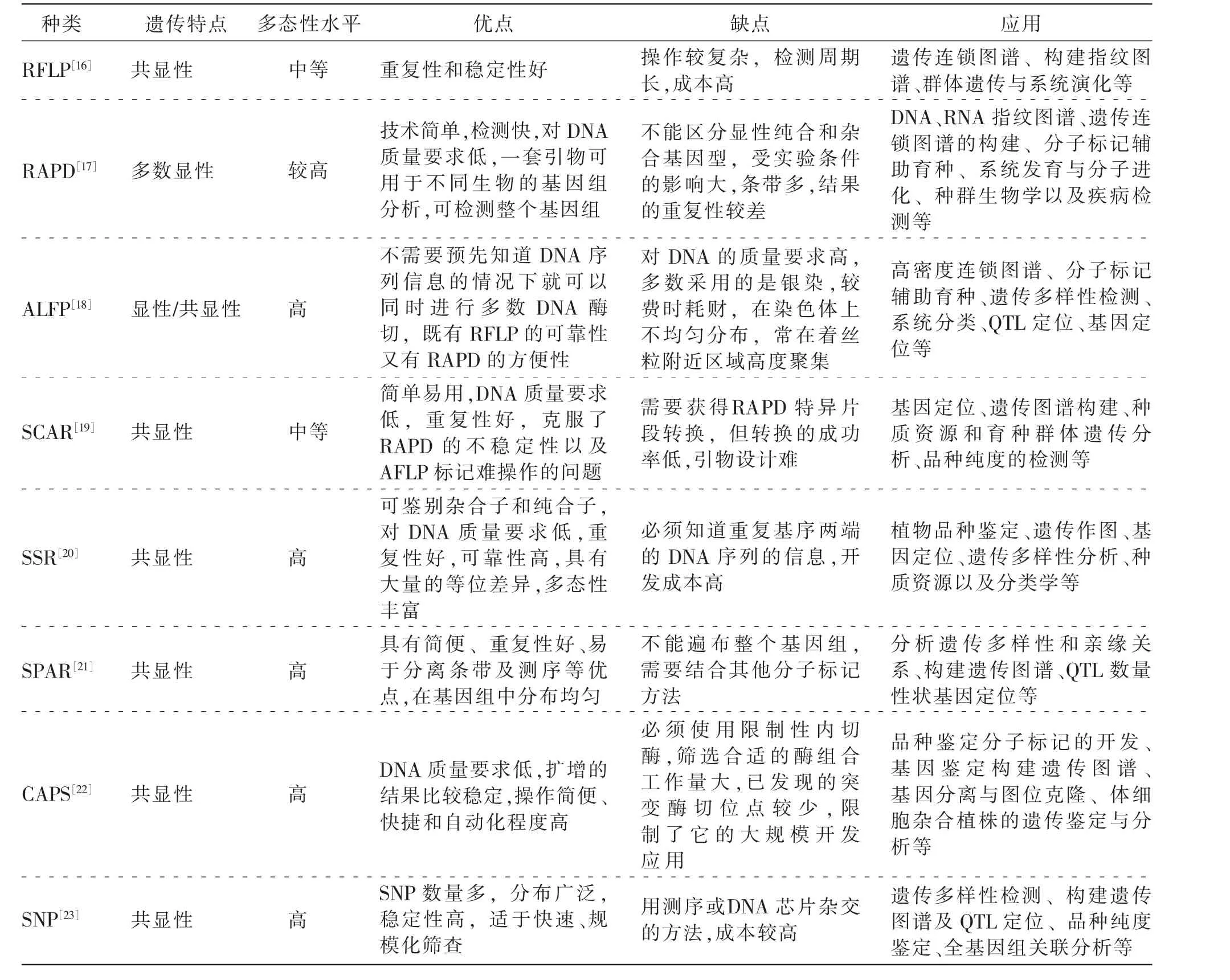
表1 常用分子标记特点
2 转录组学研究
转录组测序在牧草种质资源评价、 重要性状形成机制以及优异基因资源挖掘、 分子标记开发等方面取得了一系列重要进展。 转录组学可以提供全部基因的表达情况, 又可以反映基因蛋白互作的信息情况[24]。通过转录组测序得到的RNA 转录本信息完整,准确度又高,使得它在生物学、医学、 农学等多个领域都具有十分广泛的应用。 而RNA-Seq 技术可以直接对大多数生物体的转录本进行分析, 不需要了解所测物种的基因组信息背景,表现出很好的优势。 RNA-Seq 技术是目前深入研究转录组复杂性的强大工具, 该技术在作物抗逆、抗病基因发掘、种质资源评价中具有广泛的应用前景。 在高丹草 (Sorghum bicolor × sudanense)[25]、黄花苜蓿(Medicago falcata)[26]、紫花苜蓿 (Medicago sativa)[27]、 高羊茅 (Festuca arundinacea)[28]、羊 草(Leymus chinensis)[29]等 重 要 牧 草中,均有利用转录组测序技术进行植物生长发育、抗性等相关研究。 此前研究中缺乏巨菌草的转录组信息。 周晶等[30]采用高通量测序技术对巨菌草幼叶和根进行转录组测序, 对获得的单基因簇和差异表达基因开展功能注释、代谢通路、转录因子等方面的分析及研究。 鉴定到的上调差异基因主要富集在电子传递、碳水化合物代谢过程、离子结合等功能; 下调差异基因主要富集在代谢过程和催化活性等过程。 其中,巨菌草幼叶、根差异表达基因最主要的转录因子家族是bHLH 和WRKY。该研究极大地丰富了巨菌草不同组织中的转录组信息, 为今后其分子生物学研究提供了宝贵的数据基础。
实时荧光定量PCR 技术常被用于检测基因表达与转录分析等研究[31]。 内参基因是内源性参照基因, 由于时空的变化,RNA 的表达水平并不恒定, 不同阶段内以及不同类型的细胞或组织都可能存在差异。 使用合适的内参基因是获得准确试验数据的前提,因此需要根据实际实验情况,从多种内参基因中筛选出适合自己实验的稳定表达的内参基因对目的基因的相对表达量进行校正和标准化[32]。 李苏涛等[33]对巨菌草内参基因进行筛选,以巨菌草的叶片、茎、根为试验材料,通过分析8 个基因18S rDNA、Actin、GAPDH、ACTB、EF-1α、UBQ、CYP、TUB 在正常生长、 干旱以及盐碱胁迫下荧光定量PCR 中稳定性表达情况,最终筛选出ACTB 为巨菌草的稳定内参基因,为进一步开展巨菌草功能基因挖掘和功能基因表达分析奠定了基础。
3 巨菌草抗逆基因研究
植物在生长过程中常常会受到多种逆境胁迫,而对于巨菌草来说,不耐寒的特性和对干旱有限的耐受性很大程度地限制了其推广应用,因此,提高巨菌草对寒冷和干旱等非生物胁迫的抵抗能力对畜牧业和地球生态具有重要意义。 想要改良巨菌草的抗逆性, 最有效的办法就是培育抗逆品种[34],而解析巨菌草基因组信息及挖掘抗逆基因是培育抗逆品种亟待研究的问题。 2023 年,福建农林大学国家菌草工程技术研究中心林占熺团队构建近乎完整的异源四倍体基因组, 解析了巨菌草两套亚基因组的起源和演化过程, 并揭示了巨菌草高光合效率的机制。目前,关于巨菌草基因调控机理的研究也有少量的报道[35]。 Zhou 等[36]首次在转录组水平上研究了不同干旱和复水条件下巨菌草基因与干旱胁迫的关系, 为揭示巨菌草抗旱的分子机制提供新的思路, 并为抗旱研究提供了丰富的遗传资源。 李明珠等[37]采用转录组测序技术对盐胁迫下的巨菌草及盐胁迫下外源钙处理的巨菌草进行转录组测序, 发现盐胁迫下响应外源钙基因显著富集在碳水化合物代谢过程、 脂质代谢过程与光合作用过程, 外源钙处理能通过上调这三个过程中的基因表达, 以缓解盐胁迫对巨菌草造成的伤害。 该试验从分子水平上研究了盐胁迫下巨菌草响应钙离子的差异基因表达情况,解析了钙离子对盐胁迫下巨菌草的影响, 为提高巨菌草的耐盐性提供了科学参考, 对巨菌草今后在盐碱地的种植以及盐碱地的有效利用具有重大意义。 姜晓梦等[34]对CAMTA 转录因子家族进行了全基因组范围的系统性鉴定和分析, 在巨菌草全基因组中总共鉴定出15 个CAMTA 转录因子家族成员, 鉴定到7 个CfuCAMTA 转录因子的启动子上游含有响应低温的顺式作用元件,CfuCAMTA7 与模式植物拟南芥中参与正调控低温响应的AtCAMTA3 有相似的序列特征。 研究基于巨菌草基因组序列, 探讨了巨菌草CAMTA 家族成员在干旱胁迫下转录水平的变化, 以及CfuCAMTA 转录因子响应干旱胁迫的影响。
4 巨菌草在生态修复治理中的应用
4.1 调节土壤微生物
土壤微生物与地上植物的生长有密切关系,植物可以通过改变土壤微生物群落特征影响土壤环境[38],而土壤微生物在土壤有机质分解和生态系统养分循环的过程中又发挥着关键作用, 影响植被的发育与演替[39]。 不同生长时期的巨菌草均具有丰富的内生菌, 不仅能够提供具有多种价值的菌种资源, 还可通过与体内和根系微生物的互作,提高种植地土壤的养分含量和微生物多样性,改良荒漠化土地。 关于巨菌草在改良土壤理化特性方面已有报道。 王利蒙等[40]选择内蒙古达拉特旗、奈曼旗、土默特左旗进行巨菌草引种实验,发现种植巨菌草可有效改善土壤物理性质、 土壤透气状况与水分保持等性能,使土壤结构趋于稳定。林兴生等[1]研究发现不同生长年限的巨菌草土壤微生物对不同碳源的利用随培养时间延长而增大, 种植巨菌草于荒坡地可增加其土壤微生物群落功能多样性,在一定程度上提高土壤肥力,产生一定的生态正效应。 崔绾彤等[41]对包括巨菌草在内的6 个菌草品种种植前后的土壤养分、 土壤酶活性进行检测, 采用Illumina-MiSeq 高通量测序技术对微生物群落和多样性进行分析, 种植菌草与未种植菌草的土壤相比,速效氮、磷、全氮和有机碳等养分含量、 土壤脲酶活性以及土壤微生物多样性均显著升高。 刘凤山等[42]收集了黄河上中下游不同地区种植巨菌草和裸地的土壤, 系统地分析了巨菌草对土壤肥力、 微生物和酶活性的影响。结果表明,巨菌草能降低土壤pH 值和全钾、增加有机质、全氮、全磷、细菌、真菌、放线菌、过氧化氢酶、 多酚氧化酶和脲酶等作用, 消除裸地的影响,表明巨菌草具有改善土壤生态环境的作用。
4.2 修复土壤重金属污染
对生态环境具有危害的重金属污染物主要指镉、铅、铜、锌、汞、铬、镍、类重金属砷等。天然来源的各种重金属对生态环境危害较小, 而人为活动包括工业污染源、 生活污染源以及农业污染源产生的各种重金属是土壤重金属污染的主要来源[43]。 2014 年的《全国土壤污染状况调查公报》中显示我国土壤环境状况总体不容乐观, 耕地土壤环境质量堪忧。 一些典型地块重污染物点位超标率甚至高达36.9%[44]。 目前已有多种土壤修复技术, 而巨菌草改善土壤生态环境的同时对土壤重金属还具有一定的富集作用, 因此被用来与钝化剂联合进行综合调理。 李杰[45]以甘肃省某重金属复合污染农田土壤为研究对象, 评估掌握了研究区土壤重金属污染现状并确立了以羟基磷灰石配施腐殖酸联合巨菌草的处理方案进行实地土壤调理措施,可有效治理白银复合重金属污染农田。巨菌草促进了重金属从土壤向植物转移从而使土壤中活性重金属富集,实现重金属的部分去除,还能改善土壤微生物群落结构,优化土壤微环境。宋战旺[46]以膨润土、磷酸二氢钾、氢氧化钙为改性剂,土壤上种植巨菌草进行联合修复, 结果发现土壤中不同重金属的降低能力为铅>锌>铜>汞>镉。 随着时间的增加, 土壤中重金属下降幅度先增加后降低,最终确定60 d 的修复时间为最佳值,且巨菌草不同部位对重金属吸附能力表现为根>茎>叶。 王玺洋等[47]在野外小区试验施加不同用量稻秆炭与巨菌草联合对铜镉污染土壤进行修复,发现施加稻秆生物炭可以促进巨菌草在Cu-Cd 污染土壤中存活,提高其地上部生物量。稻秆炭在固化土壤Cu、Cd 的过程中, 一定程度上抑制了巨菌草对Cu 的吸收和富集, 但促进了巨菌草对Cd 的吸收和富集。 姬朋朋等[8]利用盆栽巨菌草实验,研究了不同土壤Cd 浓度条件下,接种两种丛枝菌根真菌和根内根孢囊霉后显著降低了土壤中可交换态和可还原态Cd 含量及其生物可利用性,并显著提高了巨菌草的重金属富集系数, 从而提高了巨菌草修复Cd 污染土壤的能力。
4.3 水土保持和荒漠化防治
在内蒙古干旱、半干旱以及部分湿润、半湿润地区均存在荒漠化严重现象,荒漠化不仅造成土地生产力降低,同时对环境以及人民的生活和生产都带来了极大的损害。 张博[48]对内蒙古乌兰布和沙漠、土默特左旗、奈曼旗三个地区种植巨菌草前后土壤进行采样,发现种植巨菌草后土壤物理指标得到改善, 且随着种植时间的增加,土壤中各物理指标改良效果逐渐增强。 在不同种植时间内,不同样地对土壤中各理化性质指标影响程度存在差异。 土默特左旗、奈曼旗种植巨菌草土壤改良效应最高,对乌兰布和沙漠改良则需要继续研究以及对改良技术进行优化,包括结合其他植物间作综合治理、改良种植行间距等。 梅兰等[49]的研究表明不同生长时期的巨菌草均能明显促进砒砂岩地区土壤细菌菌群多样性和丰富度,有效提高土壤有机质含量。 将巨菌草种植于生态脆弱区域,能减少水土流失、缓解土壤板结程度、吸附重金属离子,长年种植巨菌草还能降低土壤侵蚀程度。 四川省甘孜州的天然草场退化严重,土地沙化面积大,自然灾害及森林火灾频发, 生态环境形势严峻。 在甘孜州土壤退化治理中, 巨菌草对甘孜州土地贫瘠地区的土壤治理与修复具有良好效应,对甘孜州生态环境保护起到了积极作用[50]。 崩岗侵蚀是南方花岗岩低山丘陵区普遍分布的一种严重的水土流失类型。 植被恢复是防治崩岗侵蚀的重要方法。 何恺文等[5]研究了2 种草本植物根系对长汀县崩岗洪积扇土壤水分状况的影响,巨菌草和宽叶雀稗均对改善崩岗洪积扇土壤水分状况有明显的效果。 林占熺团队针对黄河流域不同类型生态脆弱区的生态治理难题建立了一整套巨菌草栽培技术体系,取得了多项国际领先的研究成果[9]。 在内蒙古鄂尔多斯市准格尔旗暖水乡圪秋沟开展砒砂岩区的生态修复试验。 针对不同砒砂岩类型,建立了集整地技术、淤地坝技术、优质菌草种苗选择及草灌乔配置方式为一体的菌草治理技术标准,形成了以小流域为单元、独具特色的菌草生态治理砒砂岩模式——基于雨养菌草建植技术的小流域空间梯层治理模式[51]。
5 小结
巨菌草在生态治理中不仅能修复生态脆弱地区、重金属污染土壤,还能防风固沙、提高土壤中微生物的数量和多样性,极大地改善土壤环境。除了在生态治理方面的应用, 巨菌草在其他多种应用方面已获得丰硕的成果, 但依旧存在着一些问题。巨菌草的研究体系尚未完善,由于狼尾草属形态上相似,因此导致菌草的命名混乱,而巨菌草由非洲引进我国,且是野生栽培品种,其种质资源遗传背景模糊, 因此在分子生物学理论研究方面相对薄弱,从而也限制了巨菌草品种的改良。随着测序技术和多种组学的快速发展, 对于巨菌草种质资源挖掘和优良基因克隆及抗逆机制等方面的研究提供了分子水平的大数据平台。
