马银花热激蛋白90基因家族的生物信息学及表达分析
2023-11-14李丰延瞿方茜赵芳梦王琪周泓张亮生夏宜平王秀云
李丰延,瞿方茜,赵芳梦,王琪,周泓,张亮生,夏宜平,王秀云
(浙江大学农业与生物技术学院,观赏植物基因组与基因工程实验室,浙江 杭州 310058)
热激蛋白(heat shock protein, Hsp)是广泛存在于原核生物和真核生物体内的一类能受环境刺激诱导的、具有分子伴侣功能等特征的蛋白质,参与植物生长发育调节、逆境胁迫响应等过程。植物中存在5 个公认的热激蛋白家族,分别是Hsp100、Hsp90、Hsp70、Hsp60、sHsp[1],同一家族的热激蛋白在进化上高度保守。Hsp90 是一类受腺苷三磷酸(adenosine triphosphate, ATP)调节的同源二聚体分子伴侣,由结合并水解ATP的N端结构域、用于识别底物蛋白和辅助分子伴侣的中间结构域、参与二聚体形成的C 端结构域组成[2]。一方面,Hsp90 在细胞内具有高度的组成型表达[3],参与植物信号转导、蛋白质转运与降解等过程,在胚[4]、气孔[5]、花[6]等组织器官的发育中发挥重要作用;另一方面,Hsp90 的表达也受多种环境因子诱导,在植物响应高温等逆境胁迫时发挥重要作用。研究表明,Hsp90 存在对遗传信息表达的缓冲效应,Hsp90 中一些罕见突变的表型效应被隐藏或沉默时,既能控制生物表型趋于稳定,又能维持一定的遗传多样性[7]。目前,Hsp90 基因家族在拟南芥[8]、烟草[9]、番茄[10]、白菜[11]、杨树[12]等模式植物中均已得到鉴定,但在观赏植物中研究较少。
杜鹃花是杜鹃花科(Ericaceae)杜鹃花属(Rhododendron)植物的统称。杜鹃花因花期长、花色艳丽、品种多等优点,得到世界园艺行业的青睐,成为国内外园林绿化领域重要的构成元素;然而,杜鹃花对阴凉湿润气候的喜好限制了它的栽培应用,夏季高温天气成为制约杜鹃花在中低纬度地区城市园林中应用的重要因素。 马银花(Rhododendron ovatum)是杜鹃花属马银花亚属中一种极具观赏性和应用价值的园艺植物,它拥有优雅的花形、令人愉悦的香气,同时它分布在海拔较低的地区,环境适应性较广。此前,本课题组已完成对马银花的高质量基因组测序[13],为在全基因组水平上对马银花Hsp90基因家族进行鉴定和研究提供了充分条件。因此,本研究利用生物信息学方法对马银花Hsp90 基因家族成员进行鉴定,并对其基因结构、表达模式等进行分析,为进一步研究马银花Hsp90基因的功能奠定基础,进而为杜鹃花的栽培应用和耐热育种等工作提供参考。
1 材料与方法
1.1 Hsp90 家族基因鉴定
马银花的全基因组蛋白序列信息由本课题组测序获得,已提交至RPGD 杜鹃花属基因组数据库(http://bioinfor.kib.ac.cn/RPGD/index.html)。杜鹃花属杜鹃(R.simsii)、河南杜鹃(R.henanense)、马缨杜鹃(R.delavayi)的基因组数据信息[14-16]也获取自RPGD 数据库。模式植物拟南芥(Arabidopsis thaliana)、番茄(Solanum lycopersicum)的基因组数据信息获取自Phytozome数据库(https://phytozomenext.jgi.doe.gov/)。茶(Camellia sinensis)的基因组数据信息获取自Tea Plant Information Archive 数据库[17](http://139.196.163.62)。
通过Pfam 数据库(http://pfam.xfam.org/)获取Hsp90保守区域的隐马尔可夫模型(PF00183),将其作为搜索条件,通过TBtools软件[18]从上述物种全基因组蛋白序列中筛选Hsp90序列并去除冗余。为避免错漏,利用NCBI数据库的BLAST功能对获取到的蛋白序列进行确认,并利用NCBI的CD-search在线程序(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)进行结构域预测,确保所有序列均为Hsp90序列。
1.2 马银花Hsp90 家族理化性质分析和亚细胞定位预测
利用在线工具Expasy-ProtParam[19](https://web.expasy.org/protparam/)分析马银花Hsp90家族成员的理化性质。利用在线软件WoLF PSORT(https://wolfpsort.hgc.jp/)预测马银花Hsp90 家族成员的亚细胞定位,以得分最高者为最终预测结果,若多项得分较高者得分相近则均予以保留。
1.3 Hsp90 家族进化分析
利用MAFFT 7.467软件对马银花,模式植物拟南芥、番茄,杜鹃花目植物茶,以及已完成基因组测序的杜鹃花属植物杜鹃、马缨杜鹃、河南杜鹃的Hsp90 家族成员进行多重序列比对,将比对结果导入FastTree软件,基于最大似然法构建系统进化树,再利用iTOL 在线网站(https://itol.embl.de/)对进化树进行图像美化,由此分析马银花Hsp90 与相关物种Hsp90之间的进化关系。
同时,利用TBtools 软件完成马银花全基因组所有蛋白质序列的自身比对以及马银花和相关植物拟南芥、茶、杜鹃基因组之间的两两比对。基于比对结果对马银花Hsp90家族进行物种内共线性分析以及对马银花、杜鹃、茶、拟南芥Hsp90 家族进行物种间共线性分析,利用TBtools 软件对分析结果进行可视化,并确定马银花Hsp90 家族成员的同源基因。同时,使用TBtools软件计算马银花Hsp90家族成员与其同源基因之间的非同义替换率(Ka)、同义替换率(Ks)和两者的比值(Ka/Ks)。
1.4 马银花Hsp90 家族基因结构分析
利用在线软件MEME Suite(https://memesuite.org/meme/tools/meme)预测马银花Hsp90蛋白序列的基序(motif),保存MAST.XML格式的输出文件。将基因组注释文件、MEME输出文件导入TBtools软件,利用TBtools 软件对基因的编码序列(coding sequence, CDS)结构和基序结构进行可视化。
1.5 马银花Hsp90 家族基因顺式作用元件分析
利用TBtools 软件提取马银花Hsp90 基因家族成员CDS上游2 000 bp的序列。利用New PLACE数据库[20](https://www.dna.affrc.go.jp/PLACE/?action=newplace)预测其中的顺式作用元件,并利用TBtools软件对预测结果进行可视化。
1.6 马银花Hsp90 家族染色体定位分析
根据基因组注释信息,利用TBtools 软件将马银花Hsp90 基因家族成员定位到马银花染色体上,并对结果进行可视化。
1.7 马银花Hsp90 家族基因表达分析
基于本课题组测序的马银花转录组数据进行马银花Hsp90 家族基因表达分析,原始数据文件已上传至NCBI 的SRA 数据库(SRA 登录号:PRJNA671625)。从中筛选了Hsp90 家族在不同组织器官中的表达信息,包括成熟叶片、幼嫩叶片、嫩茎、芽鳞、萼片、花瓣、雄蕊、心皮以及不同花基数的花。另外,将马银花幼苗分别放置在25、37、42 ℃的人工气候箱中处理,在高温响应基因最活跃(1 h)时剪取叶片并用液氮速冻,每个处理设置3个生物学重复。使用TBtools 软件绘制马银花Hsp90基因的表达热图,分析马银花Hsp90 基因家族在不同组织器官中的差异表达情况以及受高温胁迫的诱导情况。
2 结果与分析
2.1 马银花Hsp90 家族基因鉴定结果
对马银花进行隐马尔可夫模型搜索,并利用NCBI 的BLAST 功能对获取到的蛋白序列进行确认,共获得11个Hsp90家族成员。根据在系统进化树中与拟南芥Hsp90 成员的亲缘关系对马银花Hsp90 进行命名。对所有Hsp90 序列进行理化性质分析和亚细胞定位预测,结果(表1)显示,9 条马银花Hsp90 家族成员序列的分子量在80.11~94.17 kDa 范围内,而RoHsp90.7-3和RoHsp90.7-4这2条序列长度较短,分子量较小,可能是某些进化事件导致基因不完整。除RoHsp90.7-3外,其他马银花Hsp90家族成员的理论等电点(pI)的波动范围在4.84~5.55 之间。亚细胞定位预测结果显示,在11 个马银花Hsp90 家族成员中,可能定位于细胞质的有4个,可能定位于细胞核的有1个,可能定位于内质网的有2个,可能定位于线粒体的有3个,可能定位于质体的有3 个,可能同时定位于线粒体和质体的有2个。

表1 马银花Hsp90家族成员理化性质分析及亚细胞定位预测Table 1 Physicochemical property analysis of protein characterization and prediction of subcellular localization of Hsp90 family members in R.ovatum
2.2 Hsp90 家族进化分析结果
本研究选取模式植物拟南芥、番茄,杜鹃花目植物茶,杜鹃花属植物马缨杜鹃、杜鹃、河南杜鹃和马银花共同构建Hsp90 家族系统进化树,结果如图1 所示。根据系统进化树的拓扑结构,可划分为4 个不等大的主要分支(分支Ⅰ、Ⅱ、Ⅲ、Ⅳ),分别对应Hsp90C、TRAP、Hsp90B、和Hsp90A 4 个亚家族[21]。马银花Hsp90 家族成员不均匀地分布在4 个分支中。分支Ⅰ(cladeⅠ)仅包含RoHsp90.51 个成员,对应Hsp90C 亚家族,主要定位于质体中。分支Ⅱ(cladeⅡ)包含RoHsp90.6-1、RoHsp90.6-2 2个成员,对应TRAP 亚家族,主要定位于线粒体中。分支Ⅲ(clade Ⅲ)包含RoHsp90.7-1、RoHsp90.7-2、RoHsp90.7-3、RoHsp90.7-44个成员,对应Hsp90B亚家族,主要定位于内质网中,但是亚细胞定位预测结果显示,RoHsp90.7-3、RoHsp90.7-4 不具有高分值的内质网定位区段。分支Ⅳ(clade Ⅳ)包含RoHsp90.1-1、RoHsp90.1-2、RoHsp90.2、RoHsp90.3 4 个成员,对应Hsp90A 亚家族,主要定位于细胞质中,同时,其基因结构较为简单,内含子数量较少。
为进一步研究Hsp90基因家族的同源关系,选择和马银花亲缘关系较近的杜鹃Hsp90家族以及茶、拟南芥Hsp90家族进行多物种共线性分析,同时对马银花Hsp90家族进行物种内共线性分析。多物种共线性分析结果(图2)表明,杜鹃Hsp90家族中的8个成员和马银花Hsp90家族中的9个成员组成了11对直系同源关系,茶Hsp90 家族中的9 个成员和马银花Hsp90 家族中的8 个成员组成了10 对直系同源关系,拟南芥Hsp90家族中的4个成员和马银花Hsp90家族中的5个成员组成了5对直系同源关系。马银花Hsp90 家族物种内共线性分析结果(图3)表明,RoHsp90.7-1、RoHsp90.7-2、RoHsp90.7-4并系同源,RoHsp90.2和RoHsp90.3并系同源。以近缘物种杜鹃Hsp90 基因家族为参照,依据共线性分析得到的Hsp90 家族成员基因对间的同源关系,计算马银花Hsp90 家族成员与其同源基因对之间的Ka、Ks及Ka/Ks。结果(表2)表明,各同源基因对间的Ka/Ks<1,说明Hsp90家族在杜鹃花属分化过程中受到纯化选择作用。
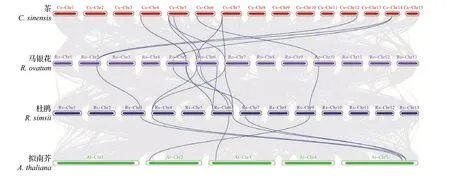
图2 拟南芥、杜鹃、茶、马银花Hsp90基因的物种间共线性分析Fig.2 Collinearity analysis of Hsp90 genes among A.thaliana, R.simsii, C.sinensis, and R.ovatum

图3 马银花Hsp90家族的物种内共线性分析Fig.3 Collinearity analysis of Hsp90 family in R.ovatum
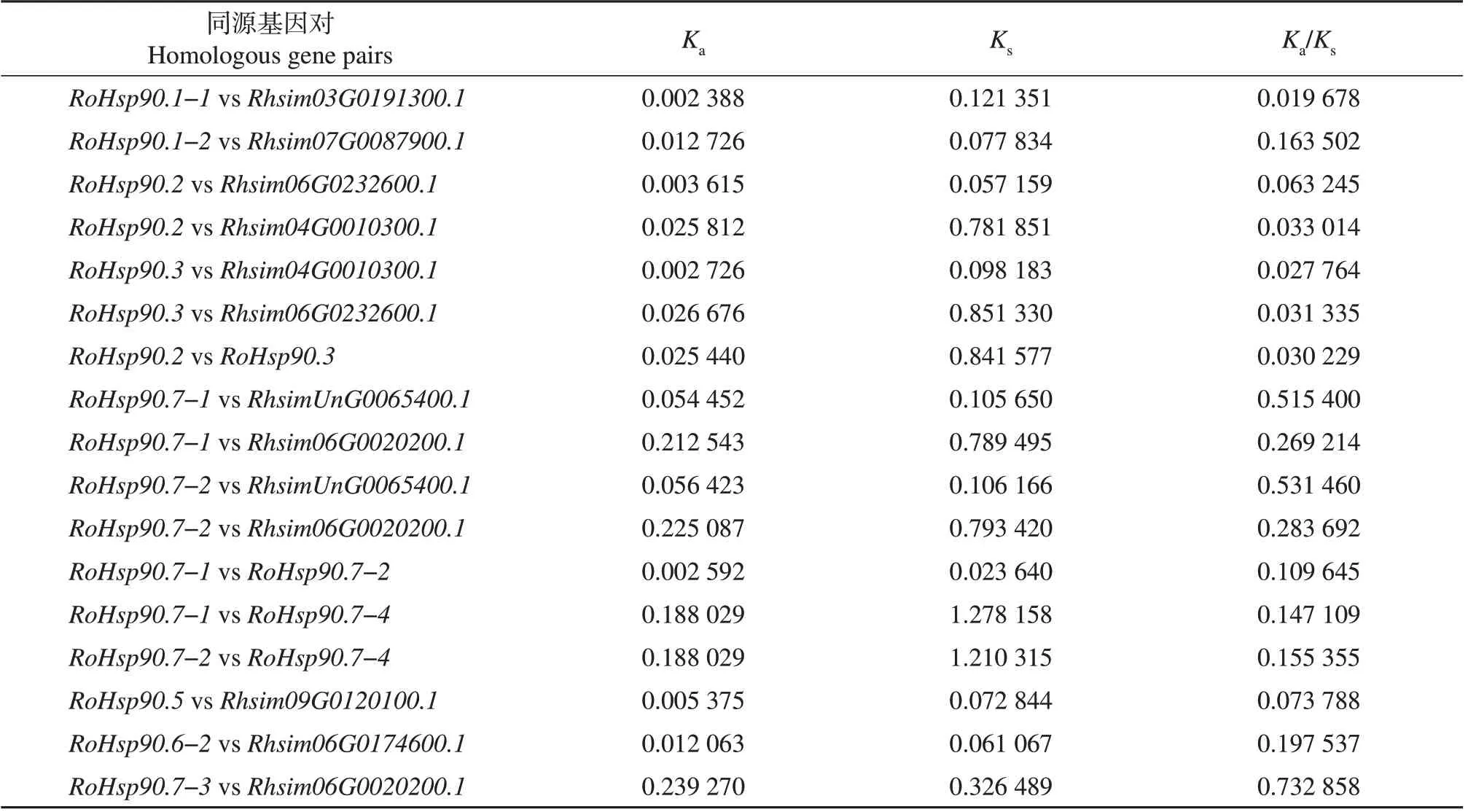
表2 马银花和杜鹃Hsp90同源基因对的Ka/Ks分析Table 2 Ka/Ks analysis between Hsp90 homologous gene pairs in R.ovatum and R.simsii
2.3 马银花Hsp90 家族基因结构分析结果
通过MEME Suite 在线软件从马银花Hsp90 家族中共识别获得10个保守基序,除RoHsp90.7-3和RoHsp90.7-4这2条较短的序列分别具有4个和2个基序外,马银花Hsp90家族其他成员均具有这10个基序(图4)。基因结构分析发现,如果忽略基因结构可能不完整的RoHsp90.7-3和RoHsp90.7-4,位于马银花Hsp90 家族系统进化树基部的RoHsp90.1-1、RoHsp90.1-2、RoHsp90.2、RoHsp90.3 均仅具有2~3个内含子,基因结构相对简单,而其他家族成员则往往具有10个以上的内含子,基因结构较为复杂。

图4 马银花Hsp90家族保守基序及基因结构Fig.4 Conserved motifs and gene structures of Hsp90 family in R.ovatum
2.4 马银花Hsp90 家族顺式作用元件分析结果
对马银花Hsp90基因家族进行顺式作用元件预测,主要关注植物激素代谢和逆境胁迫响应过程相关元件的分布。结果(图5)表明,马银花Hsp90 家族11 个成员上游的顺式作用元件均涉及植物激素代谢和逆境胁迫响应,涉及的植物激素包括生长素、水杨酸、赤霉素、脱落酸、乙烯、细胞分裂素、茉莉酸,逆境胁迫包括高温、低温、损伤、水分胁迫、未折叠蛋白。

图5 马银花Hsp90家族顺式作用元件分析Fig.5 Cis-acting elements analysis of Hsp90 family in R.ovatum
2.5 马银花Hsp90 家族的染色体定位结果
马银花共有13 对染色体,染色体定位结果(图6)显示,Hsp90基因家族在不同染色体上分布不均。6 号染色体上分布最多,有5 个Hsp90基因;其次是5号染色体,有3个Hsp90基因;2号、7号、10号染色体上各有1 个Hsp90基因;其他染色体上无Hsp90基因。位于5 号染色体上的RoHsp90.7-1、RoHsp90.7-2,以及位于6号染色体上的RoHsp90.7-3、RoHsp90.7-4排列较为紧密,可能为串联重复类型的基因复制。

图6 马银花Hsp90基因家族的染色体定位Fig.6 Chromosomal localization of Hsp90 gene family in R.ovatum
2.6 马银花Hsp90 家族基因在不同组织器官中的表达分析结果
根据马银花Hsp90 家族成员在不同组织器官中的表达数据绘制热图,结果如图7 所示。其中,RoHsp90.1-2、RoHsp90.7-3、RoHsp90.7-4在各组织器官中未见明显表达。马银花Hsp90家族各成员的表达量在幼嫩叶片和成熟叶片之间均未表现出差异。RoHsp90.1-1和RoHsp90.6-2在花器官及其各部分组织(生殖器官)中的表达量高于成熟叶片、幼嫩叶片、嫩茎、芽鳞(营养器官),其他家族成员的表达量在营养器官和生殖器官之间未表现出明显差异。部分家族成员的表达量在不同花基数的花中存在差异:RoHsp90.1-1的表达量在花基数为5的花中最高,其次是在花基数为4的花中,而在花基数为6的花中最低;RoHsp90.3、RoHsp90.7-1、RoHsp90.7-2、RoHsp90.6-2的表达量在花基数为4 的花中最高,其中RoHsp90.7-2和RoHsp90.6-2的表达量在花基数为5 和6 的花之间差异不明显,RoHsp90.3和RoHsp90.7-1在花基数为5 的花中的表达量高于花基数为6的花;RoHsp90.2在花基数为4 和5 的花中表达量相近,但均高于花基数为6 的花;其他成员的表达在3种花基数的花之间未见明显差异。

图7 马银花Hsp90基因在不同组织器官中的表达热图Fig.7 Heatmap of Hsp90 gene expression in various tissues and organs of R.ovatum
2.7 不同温度处理下马银花Hsp90 家族基因的表达分析
根据马银花Hsp90 家族成员在25、37、42 ℃3 种温度处理1 h 条件下叶片中的表达数据绘制热图。结果(图8)显示,除RoHsp90.1-2、RoHsp90.7-3、RoHsp90.7-4在3 种温度处理下均未见明显表达外,其他成员均不同程度地受到高温(37、42 ℃)处理的诱导。其中,RoHsp90.7-1、RoHsp90.7-2、RoHsp90.5、RoHsp90.6-1、RoHsp90.6-2在37 ℃处理下表现出明显的表达上调,但是在42 ℃处理下,这些成员的表达保持不变或略有下调;RoHsp90.1-1、RoHsp90.2、RoHsp90.3则在37、42 ℃处理下均表现出明显的表达上调。这说明绝大多数马银花Hsp90家族成员的表达会受到中度高温(37 ℃)胁迫诱导,从而在高温胁迫响应中发挥生理功能。同时,大多数Hsp90家族成员的表达不会受到重度高温(42 ℃)胁迫诱导,或者被诱导后也会被高温降解,反映出马银花可能对高温胁迫的响应存在一定的温度范围限制。因此,在37、42 ℃处理下均表现出明显表达上调的Hsp90基因可能在马银花高温胁迫响应中发挥重要作用。
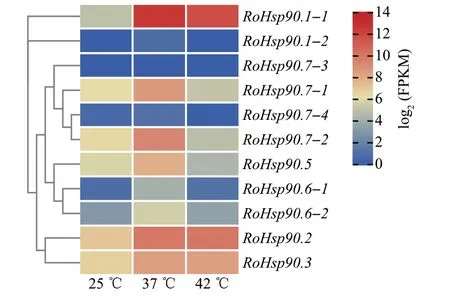
图8 不同温度处理下马银花Hsp90基因表达热图Fig.8 Heatmap of Hsp90 gene expression under different temperatures in R.ovatum
3 讨论
Hsp90 家族在植物生长发育调控、逆境胁迫响应等生命活动中发挥的重要功能已在多种园艺植物中得到研究,但在杜鹃花属植物中研究较少。本研究对杜鹃花属马银花亚属马银花的Hsp90 家族进行了全基因组鉴定,共鉴定出11个Hsp90家族成员,多于拟南芥(7 个)[8]、番茄(7 个)[10]等模式植物,但少于烟草(21 个)[9]、白菜(14 个)[11]等模式植物。马银花Hsp90 家族成员在氨基酸数量上差异很大,大多数成员编码的氨基酸数量在699(RoHsp90.3)至820(RoHsp90.7-1)范围内,但RoHsp90.7-4和RoHsp90.7-3编码的氨基酸数量则分别为117 个和517 个,原因可能是某些进化事件导致基因不完整。这一猜测被保守基序分析的结果证实,除RoHsp90.7-4 和RoHsp90.7-3 外的其他马银花Hsp90家族成员均具有全部10个保守基序,并按相同顺序排列,但RoHsp90.7-4 和RoHsp90.7-3 分别只有2个和4个保守基序,其排列顺序同其他成员。除RoHsp90.7-3外,马银花Hsp90 家族绝大多数成员理论等电点的波动范围在4.84~5.55 之间,和白菜[11]、番茄[10]等植物的研究成果类似。部分植物Hsp90 家族存在少数成员的理论等电点在9.00 附近[10,22],这一结果与RoHsp90.7-3相近。基因结构分析结果显示,位于马银花Hsp90 家族系统进化树基部的RoHsp90.1-1、RoHsp90.1-2、RoHsp90.2、RoHsp90.3 仅具有2~3个内含子,基因结构相对简单,而分子量相近的其他马银花Hsp90 家族成员则均具有10 个以上内含子,基因结构相对复杂,这说明马银花Hsp90家族成员在进化过程中发生过内含子丢失或获得事件。
本研究对马银花及相关植物的Hsp90家族构建系统进化树,并将其划分为4个主要分支,分别能够对应Hsp90家族内Hsp90A、Hsp90B、Hsp90C、TRAP 4个主要亚家族的划分,这一点被基因结构分析和亚细胞定位预测结果佐证。Hsp90亚细胞定位预测结果与亚家族的划分并非严格对应,而是可能依据具体的基因功能发生变化[21],因此,尚需亚细胞定位实验对预测结果进行进一步验证。将马银花Hsp90家族成员划入已被报道的亚家族,有助于对其家族成员的功能进行进一步研究。Ka/Ks值常被用于判断蛋白质在进化过程中受到的选择压力,马银花和杜鹃Hsp90 家族直系同源基因对之间以及马银花Hsp90家族并系同源基因对之间的Ka/Ks值均小于1,说明Hsp90家族成员在杜鹃花属分化过程中可能受到纯化选择作用,暗示了该家族在杜鹃花的生命活动中发挥着重要功能。
本研究分析马银花Hsp90家族在不同组织器官中和高温胁迫下的表达情况发现,RoHsp90.7-3、RoHsp90.7-4、RoHsp90.1-2在马银花的不同组织器官中及不同温度处理下的叶片中均无明显表达,说明它们在进化过程中可能因结构受损(如RoHsp90.7-3、RoHsp90.7-4)或积累有害突变而假基因化,从而不表达甚至丧失正常功能。
除上述3个成员外,马银花Hsp90家族其他成员均受高温胁迫诱导,这说明Hsp90 家族在马银花的耐热性调节中发挥重要作用。而且,Hsp90家族不同成员受高温诱导的模式不同,RoHsp90.1-1、RoHsp90.2、RoHsp90.3在37、42 ℃处理下均表现出明显的表达上调,其他成员则仅在37 ℃处理下表现出明显上调,而在42 ℃处理下这一上调可能会被逆转,说明马银花Hsp90 家族的不同成员可能在植物响应高温胁迫的不同途径中发挥功能。上述Hsp90家族成员在植物响应高温胁迫过程中的多种功能与张海[23]的研究相似。
马银花Hsp90 家族在不同组织器官中的表达结果显示,RoHsp90.1-1和RoHsp90.6-2在生殖器官中的表达高于营养器官,7 个Hsp90 家族成员在不同花基数的花中的表达存在差异。这说明马银花Hsp90 家族可能参与植物花器官的发育过程。MARGARITOPOULOU 等[6]研究发现,Hsp90 家族参与植物花器官发育过程的调控,Hsp90 的缓冲作用能够调控花器官形成相关基因的表达,从而预防花的畸形发育。马银花的花基数通常为5,偶尔会出现4或6的变异,本研究中马银花Hsp90家族部分成员在不同花基数的花中的表达量存在差异,表明它们可能也参与了花器官形成相关基因的调控,在花基数的变异发育中起到一定的作用,但具体机制仍需进一步研究。
4 结论
本研究在马银花全基因组水平上鉴定出11 个Hsp90基因,对其进行蛋白质理化性质、亚细胞定位预测、染色体定位、基因结构、系统进化分析发现,不同的Hsp 家族成员表现出定位、结构的多样性。启动子区顺式作用元件分析显示,马银花Hsp90 家族能够涉及多种植物激素代谢和逆境胁迫响应。不同组织器官中和高温处理下叶片中Hsp90基因的表达模式分析表明,Hsp90 基因家族成员参与花器官发育和响应高温胁迫。本研究对于马银花Hsp90家族不同成员的特性及潜在功能研究具有指导意义,也为杜鹃花响应高温胁迫及花器官的发育研究提供了候选基因。
