杀虫剂处理的梨小食心虫转录组测序分析及差异表达解毒酶基因筛选
2023-11-14刘晓庆庞钦玮冀佳悦高玲玲郭艳琼
刘晓庆 ,庞钦玮 ,冀佳悦 ,杨 春 ,高玲玲 ,郭艳琼
(1.山西农业大学 植物保护学院,山西 太谷 030801;2.山西农业大学 果树研究所,山西 太谷 030815;3.澳大利亚联邦科学与工业研究组织农业与食品部,西澳 文布利 6913)
梨小食心虫(Grapholita molesta)属鳞翅目卷蛾科,简称“梨小”,广泛分布于亚洲、欧洲和美洲,我国从南至北均有发生[1-2],主要危害苹果、梨、核桃和红枣等仁果类和核果类果树。目前,梨小食心虫的防治手段仍以化学防治为主。梨小食心虫幼虫具有钻蛀性,难以完全接触药剂,因此,果园喷施农药通常是用于灭杀成虫,阿维菌素、高效氯氟氰菊酯是防治梨小食心虫的有效药剂,吡虫啉作为果园防治害虫常用杀虫剂,对梨小食心虫也有一定的防治效果。
杜恩强等[3]试验表明,阿维菌素能杀灭梨小成虫、抑制成虫产卵;庾琴等[4]研究表明,阿维菌素、高效氯氟氰菊酯等杀虫剂对梨小初孵幼虫有一定的防治效果;陈静等[5]研究表明,高效氯氟氰菊酯、阿维菌素对梨小具有一定的防治效果;田间药效试验发现,4.5%高效氯氰菊酯EC 对梨小15 d 田间防效达98%[6]。然而,由于梨小的钻蛀特性,使部分龄期难以完全接触药剂,且化学农药的长期不合理使用,使梨小对不少农药耐受性增加。害虫对杀虫剂产生耐药性的过程,与解毒酶的活性密切相关。解毒酶代谢活性增加在害虫产生耐药性过程中起重要作用[7-9]。
细胞色素P450(Cytochrome P450,CYP)、羧酸酯酶(Carboxylesterases,CarEs)和谷胱甘肽硫转移酶(GlutathioneS-transferases,GSTs)是昆虫三大解毒酶系。细胞色素P450 可广泛地代谢外源和内源化合物[10],在解毒和激活方面具有双重作用。近年来,对双翅目、鳞翅目和鞘翅目等多种昆虫进行了全基因组测序,与昆虫解毒代谢相关的P450基因大部分集中在CYP4、CYP6 和CYP9 家族中,通过聚类分析发现,这些细胞色素基因分属于CYP2 簇、CYP3 簇、CYP4 簇和线粒体簇中[11]。CarEs 主要是通过将包含酯键的内源性和外源性化合物分解为醇和酸[12],达到解毒目的,其共有14 个进化支(A-N),分为3 个功能组,即与代谢解毒相关的酶类(A-C)、与激素和信息素降解相关的酶类(D-G)和与神经发育相关的酶类(H-N)。GSTs 通过催化谷胱甘肽与有毒的疏水亲电物质发生轭合反应,使毒性降低,可溶性增强,进而排出体外,达到解毒的目的[13],GSTs 在昆虫中可分为七大类:sigma、zeta、omega、theta、microsomal、delta和epsilon[14]。这3 种解毒酶广泛分布于动植物和微生物中[15],可将进入昆虫体内的有毒物质转变为弱毒或无毒的形式[16]。
随着转录组测序研究的深入,高通量双端RNA测序被广泛应用,它可以测得不同组织或不同条件下的转录组数据[17],例如,人、鼠、昆虫、微生物等的转录组[18-20],此外,RNA 测序还能进行差异表达基因的分析[21]。CRAVA 等[22]对樱桃果蝇Drosophila suzukii的转录组分析,筛选出了OR 基因;TIAN等[23]对桃蛀果蛾Carposina sasakii转录组进行分析,筛选出了化学感受蛋白基因;CHEN 等[24]对韭菜迟眼蕈蚊Bradysia odoriphaga转录组分析,筛选出4 个与吡虫啉代谢有关的P450 基因。
本研究对3 种杀虫剂不同浓度处理的梨小转录组文库进行高通量测序,筛选可能与杀虫剂代谢相关的解毒酶基因,旨在为合理科学使用杀虫剂及深入开展研究梨小解毒酶基因的功能提供理论依据。
1 材料和方法
1.1 试验材料
梨小食心虫成虫初始种群于2012 年采自山西省太谷县西山底村桃林(37°21′42″N,112°34′19″E),将其带回实验室放置于温度(26±1)℃、相对湿度70%±10%、光周期15L∶9D 的人工气候培养箱中饲养。
RNAiso Plus,购买自TaKaRa;cDNA 第1 链合成试剂盒PrimeScript RT reagent kit with gDNA Eraser,购买自北京宝日医生物技术有限公司;杀虫剂为阿维菌素(avermectin)、吡虫啉(imidacloprid)、高效氯氟氰菊酯(lambda-cyhalothrin),均购买自SIGMA-ALDRICH 公司。PRX-450C 人工气候箱(宁波海曙赛福仪器厂);HS-840 超净工作台(苏州安泰空气技术有限公司)。
1.2 试验方法
1.2.1 3 种杀虫剂室内毒力测定 用药膜法[25]测定吡虫啉、阿维菌素和高效氯氟氰菊酯对梨小食心虫的室内毒力。以丙酮为溶剂将原药配制成母液,并用丙酮稀释成一系列质量分数的供试药液。分别吸取1 mL 药液加入到指形管(直径16 mm,长度80 mm)中;立即滚动小瓶,使农药均匀地分布在内表面,在丙酮挥发后用吸水棉盖住。将7 只健康的梨小成虫接入带有杀虫剂薄膜的指形管中,每组设置3 个生物学重复,原饲养条件下饲养24 h 观察死亡情况并记录死亡率,轻触虫体不动为死亡。
1.2.2 样品制备 根据室内毒力测定结果,得到吡虫啉、阿维菌素和高效氯氟氰菊酯亚致死浓度(LC10、LC30)、致死中浓度(LC50),分别吸取3 种杀虫剂3 种浓度1 mL 药液于指形管中,立即滚动小瓶,使农药均匀地分布在内表面,并将7 只健康成虫放入带有杀虫剂薄膜的小瓶24 h,然后收集存活的成虫作为测序的一个样本,每组设置3 个生物学重复。所有的样品立即在液氮中冷冻,然后储存在-80 ℃用于提取RNA。
1.2.3 RNA 提取 按照说明用Trizol RNA 提取试剂和Rnase 提取1.2.2 样品中的总RNA,然后用Rnasy 离心柱纯化。利用超微量分光光度计检测RNA 的质量,用琼脂糖凝胶电泳检测RNA 的完整性。
1.2.4 转录组测序及数据组装 采用Illumine HiSeq 2000 测序仪进行测序,对获得的原始测序数据进行过滤,去除接头和含未知碱基N 比例超过5%的reads 以及低质量测序序列,得到待分析数据(Clean reads)。使用Trinity 软件对获得的高质量测序数据进行组装,然后使用Tgicl 软件进行聚类去冗余,获得Unigene。
1.2.5 功能注释 为获得全面的基因功能信息,将非重复序列基因通过BLAST 分别与NR、NT、SWISS-PROT、Pfam、GO、KOG、KEGG 数据库进行比对,获得相应的注释信息。
1.2.6 差异表达基因分析 使用DESeq 软件对差异表达基因进行分析,筛选差异表达基因的标准为误报率(falsediscovery rate,FDR)小于0.05 且差异倍数大于2。通过MEGA 7.0 软件使用P 距离模型1 000 次引导值的邻接法进行系统发育树分析[26]。
第三、加快推进社会体育改革,构建社区体育发展机制。经过十几年体育改革发展,我国体育社会化进度在加快,体育体制开始分化,社会普及程度较高的足球、篮球、排球等项目实现职业化发展。计划经济体制下的“单位体育”逐渐向社区体育转变,体育社团、体育协会、体育俱乐部等体育社会组织的发展,推动社区体育发展,社区体育成为全民健身运动发展基本载体。
2 结果与分析
2.1 3 种杀虫剂对梨小的毒力测定
采用药膜法进行吡虫啉、阿维菌素和高效氯氟氰菊酯对梨小的室内毒性测定,计算出3 种杀虫剂的毒力回归方程以及LC10、LC30、LC50浓度(表1)。

表1 3 种杀虫剂的室内毒力测定Tab.1 Indoor toxicity determination of three insecticidesμg/mL
2.2 梨小食心虫转录组测序与组装结果
梨小转录组测序数据中,Q20 碱基和Q30 碱基的比值分别>96%和>87%,说明RNA-Seq 数据质量较好,测序数据准确可靠(表2)。

表2 测序数据统计Tab.2 Sequencing data statistics
经过组装后产生了101 873 条Unigenes,总长度、平均长度以及覆盖50%所有核苷酸的最大序列重叠群长度(N50)分别为202 980 197、1 992、3 037 bp,GC 含量为40.64%(表3)。Unigenes 其不同长度的具体分布如图1 所示,可看出>3 kb 的Unigenes 最多,有22 913 条,说明组装结果较好。
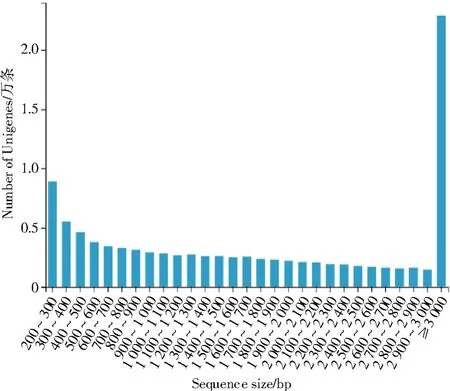
图1 梨小食心虫转录组拼接后的Unigenes 长度分布Fig.1 Length distribution of the assembled Unigenes in the G. molesta transcriptome
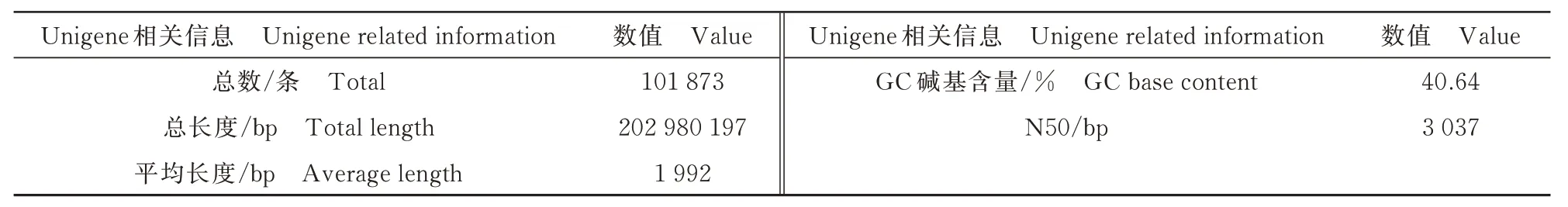
表3 组装结果统计Tab.3 Summary of assembly results
2.3 梨小食心虫转录组基因功能注释
将梨小食心虫转录组数据组装后的Unigenes序列与NR、NT、KEGG、KOG、SWISS-PROT、Pfam和GO数据库进行BLAST比对,对获得的101 873条Unigenes进行功能注释,其中通过NR、NT、SWISSPROT、Pfam、KEGG、KOG、GO 数据库比对获得注释的Unigenes 分别有57 209、39 186、43 749、46 168、46 678、42 247、27 486 条,共有66 658 条Unigenes 成功得到注释,占总Unigenes 的65.43%(表4)。根据NR 数据库比对,序列同源性最高的物种是大蜡螟Galleria mellonella(9.35%),其次是粉纹夜蛾Trichoplusia ni(9.32%)、棉铃虫Helicoverpa armigera(9.28%)、海波斯莫科马属蛾Hyposmocoma kahamanoa(9.16%)和脐橙螟蛾Amyelois transitella(8.13%)(图2)。
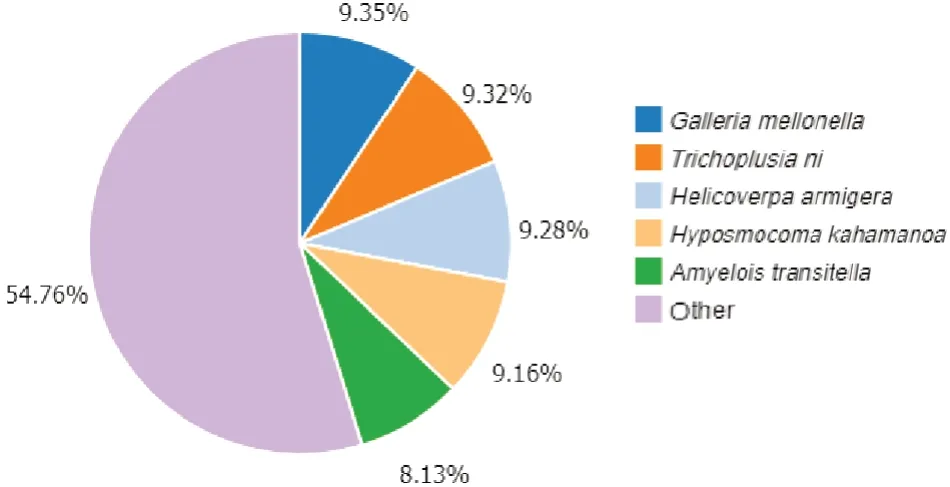
图2 梨小食心虫转录组Unigenes 在NR 数据库中的物种分布Fig.2 Species distribution of Unigenes in the G. molesta transcriptome in NR database
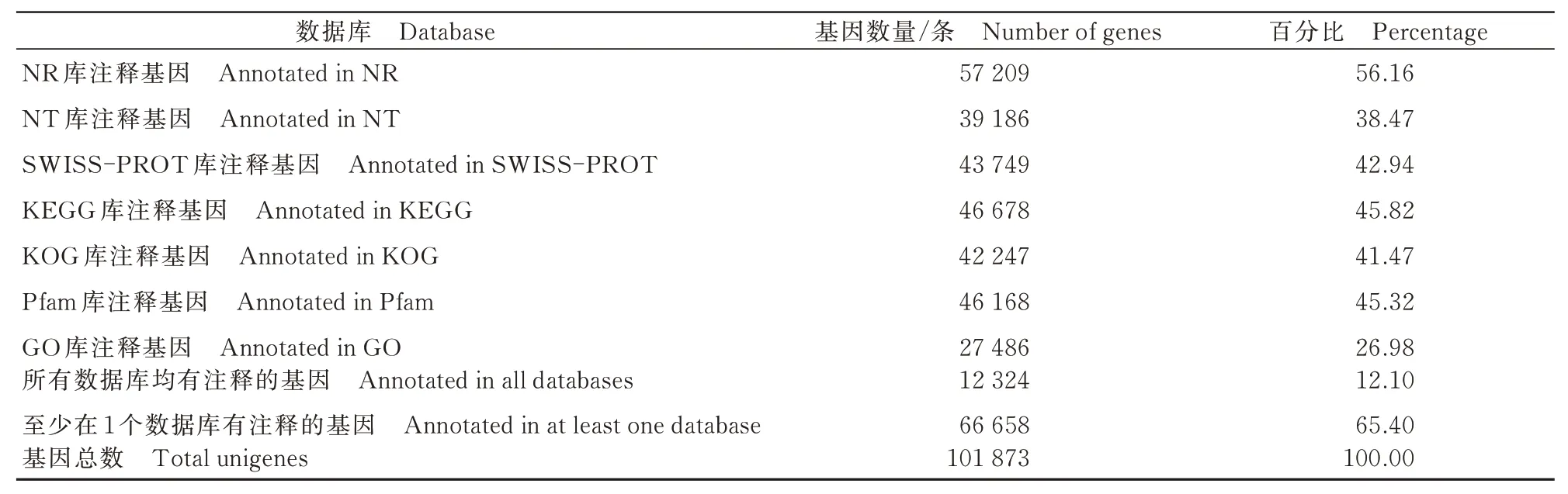
表4 梨小食心虫转录组Unigene 的功能注释信息Tab.4 Functional annotation information of G. molesta transcriptome
使用基因本体(GO)方法,将注释基因分为3 类(共24 个功能组):生物过程、细胞组成和分子功能。在生物过程中,细胞过程包含了最多的Unigenes;在细胞组分中,膜部分占主导地位;在分子功能分类中,注释基因数最多的是结合和催化活性(图3)。与KEGG 数据库比对获得注释的46 678 条Unigene主要涉及6 大类功能,包括细胞过程、环境信息处理、遗传信息处理、人类疾病、新陈代谢和生物系统。最具代表性的途径是信号转导以及运输和分解代谢(图4)。
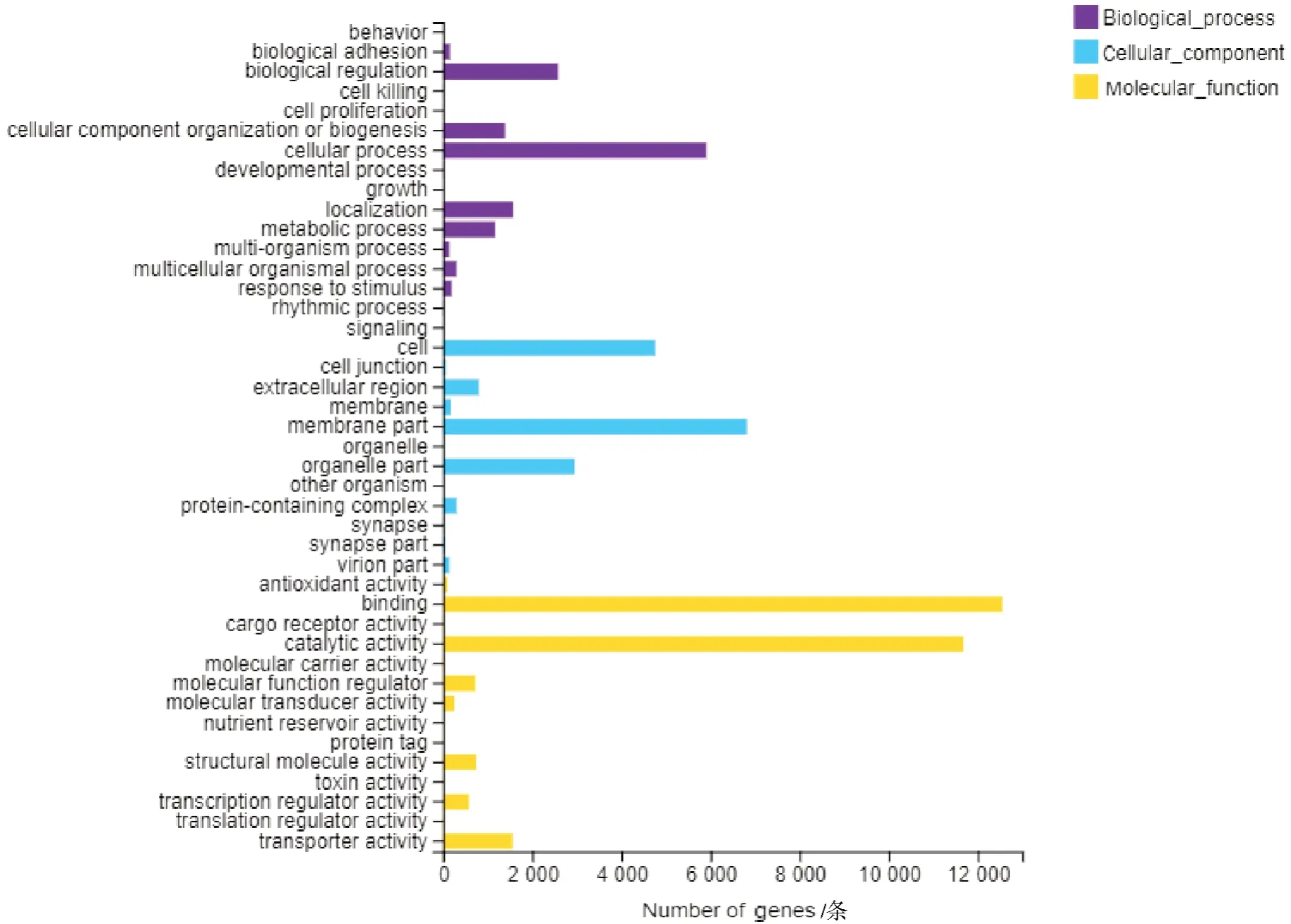
图3 梨小食心虫转录组Unigenes GO 功能分类Fig.3 GO functional classification of Unigenes in the G. molesta transcriptome
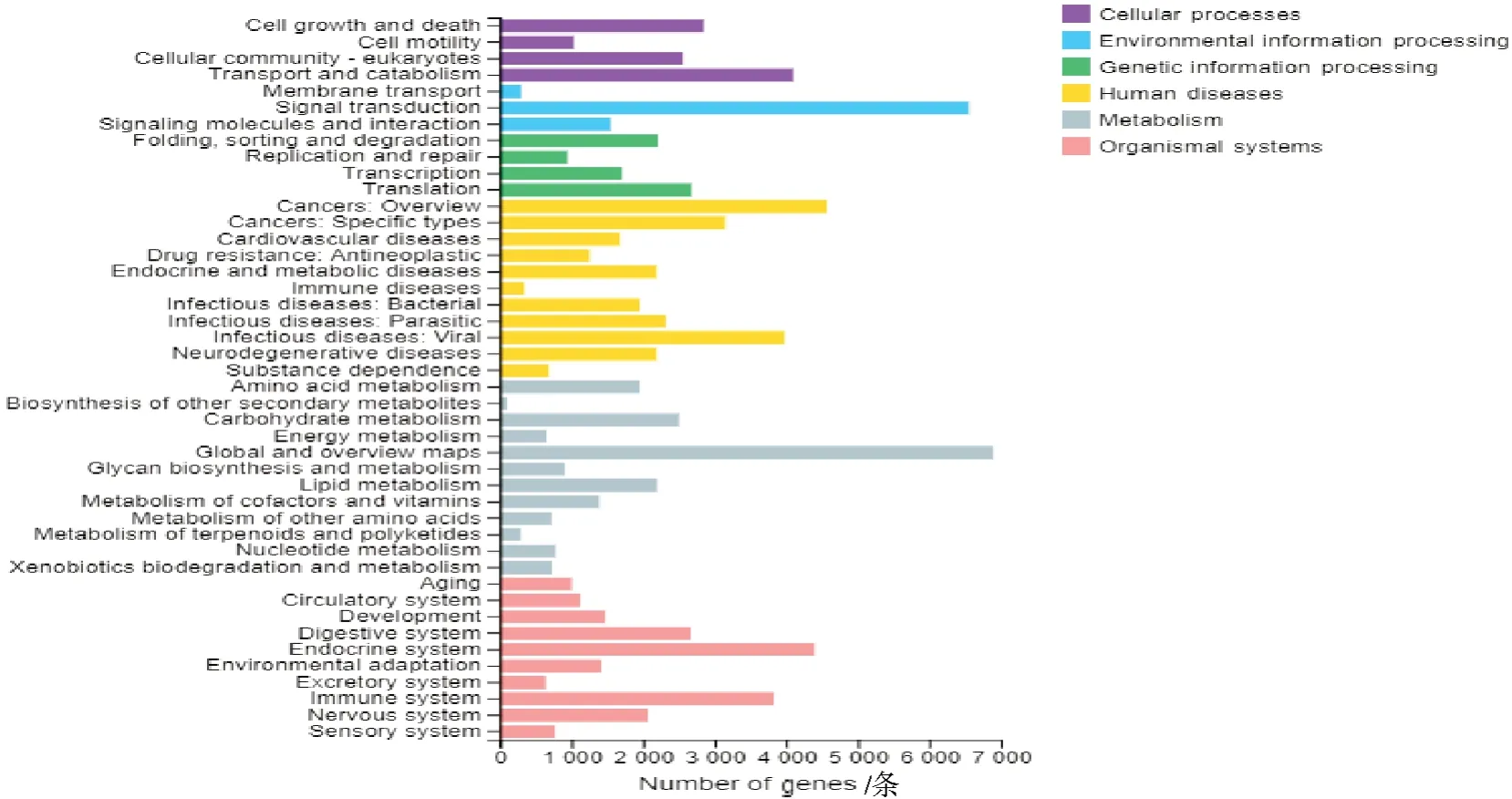
图4 梨小食心虫转录组Unigenes KEGG 功能分类Fig.4 KEGG functional classification of Unigenes in the G. molesta transcriptome
2.4 梨小食心虫差异表达基因及功能注释
采用生物信息学方法分别筛选3 种农药处理的梨小食心虫差异表达基因,阿维菌素处理梨小后筛选出825 个差异表达基因,其中639 个上调基因和186 个下调基因(图5-A),并对其功能进行注释,注释到结合(binding)的Unigenes 最多(图5-B)。
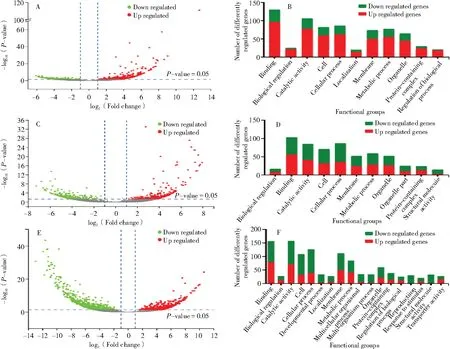
图5 差异表达量倍数变化及功能注释Fig.5 Differential expression fold change and functional annotation
吡虫啉处理后筛选出641 个差异表达基因,包括333 个上调基因和308 个下调基因(图5-C),功能注释到结合(binding)的Unigenes 最多(图5-D)。
高效氯氟氰菊酯处理后筛选出1 391 个差异表达基因,包括538 个上调基因和853 个下调基因(图5-E),并对其功能进行注释,注释到结合与催化活性(binding and catalytic activity)的Unigenes 最多(图5-F)。
2.5 梨小食心虫差异表达解毒酶基因筛选
经阿维菌素3 种浓度处理梨小后的转录组数据中,差异表达基因主要为P450s 和GSTs,其中有5 个P450,4 个上调,1 个下调;2 个GST 基因,1 个上调,1 个下调;其中P450 基因Unigene16556_All在LC10和LC50浓度下均显著上调;GST 基因CL996.Contig4_All 在LC10和LC50浓度下均显著下调(表5)。
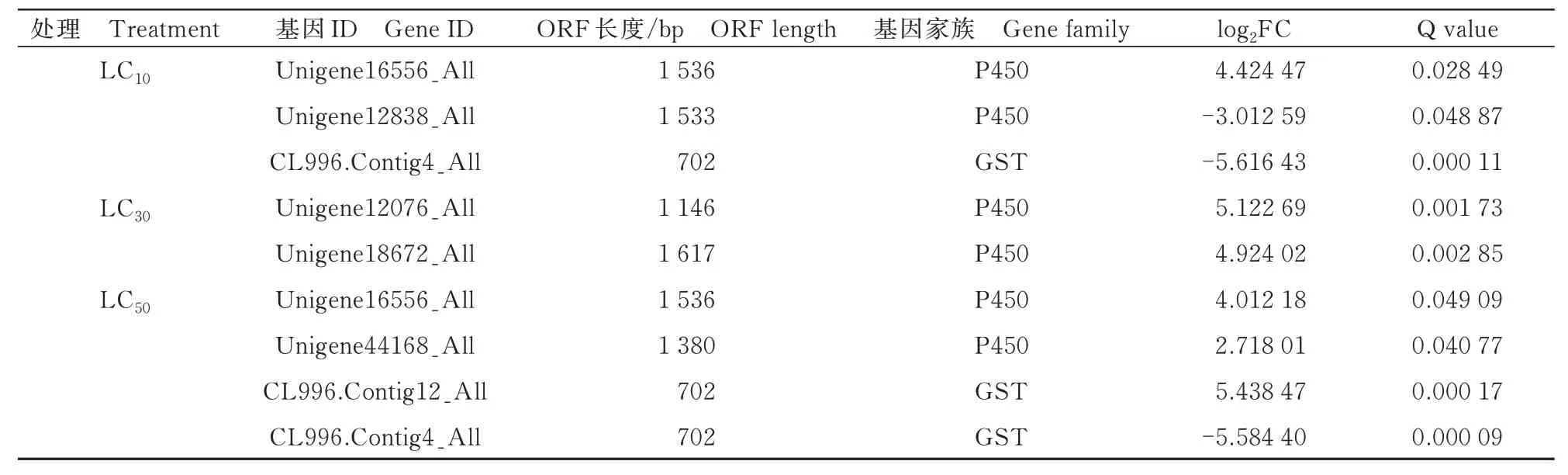
表5 阿维菌素处理下差异表达的解毒基因Tab.5 Detoxification genes differentially expressed under avermectin treatment

表6 吡虫啉处理下差异表达的解毒基因Tab.6 Detoxification genes differentially expressed under imidacloprid treatment
高效氯氟氰菊酯3 种浓度处理梨小后的转录组数据库中,3 种解毒酶基因均有差异表达,其中有6 个P450 基因,4 个上调,2 个下调;2 个GST 基因均上调;3 个CarE 基因均上调(表7)。
将3 种杀虫剂处理梨小后的差异表达基因进行汇总,共筛选到解毒代谢相关基因20 个,包括11 个P450 基因、5 个GST 基因、4 个CarE 基因,20 个差异表达基因的表达量热图聚类分析如图6所示。分析发现,2个P450基因(Unigene12076_All、Unigene18672_All)在阿维菌素、吡虫啉处理下均显著上调;P450 基因CL4554.Contig2_All 在吡虫啉、高效氯氟氰菊酯处理下显著下调;P450 基因Unigene15542_All、GST 基因CL2631.Contig4_All在吡虫啉、高效氯氟氰菊酯处理下显著上调;P450基因Unigene19792_All 在吡虫啉处理下显著下调,在高效氯氟氰菊酯处理下显著上调;P450 基因Unigene16556_All 在3 种农药处理下均显著上调。

图6 差异表达基因表达量聚类热图Fig.6 Clustering heat map of differentially expressed gene expression
将11 条P450 基因序列与已知鳞翅目昆虫的CYP 蛋白序列比对分析并构建进化树,其中有7 个基因属于CYP6 家族,2 个基因属于CYP4 家族,2 个基因属于CYP12 家族(图7)。将5 条GST 基因序列与已知鳞翅目昆虫的GST 蛋白序列比对分析并构建进化树,均属于sigma 家族(图8),将4 条CarE基因序列与已知鳞翅目昆虫的CarE 蛋白序列比对分析并构建进化树,其中3 个聚在A 簇,另一个聚在C 簇(图9)。
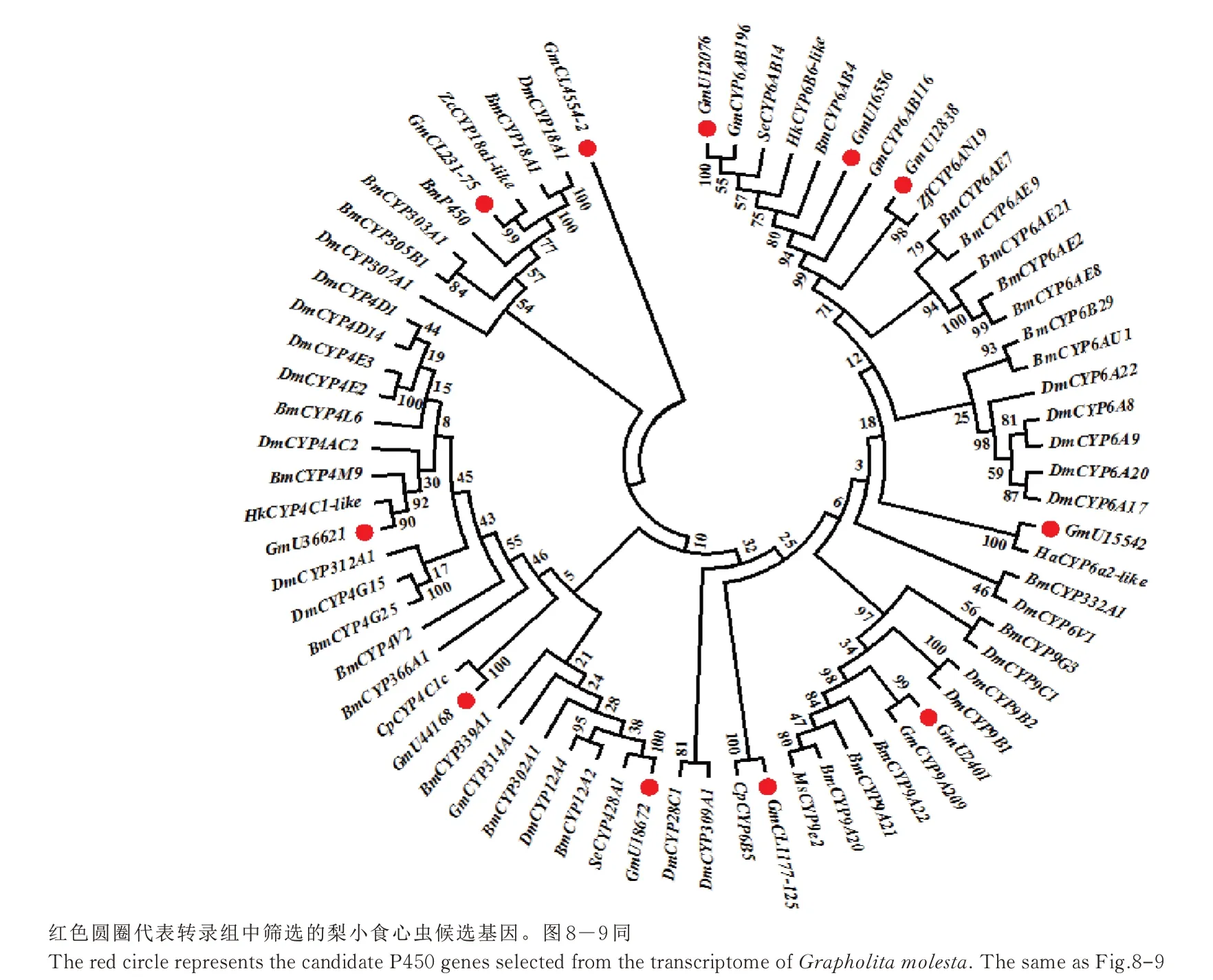
图7 梨小食心虫细胞色素P450 系统发育分析Fig.7 Phylogenetic analysis of cytochrome P450 in G. molesta
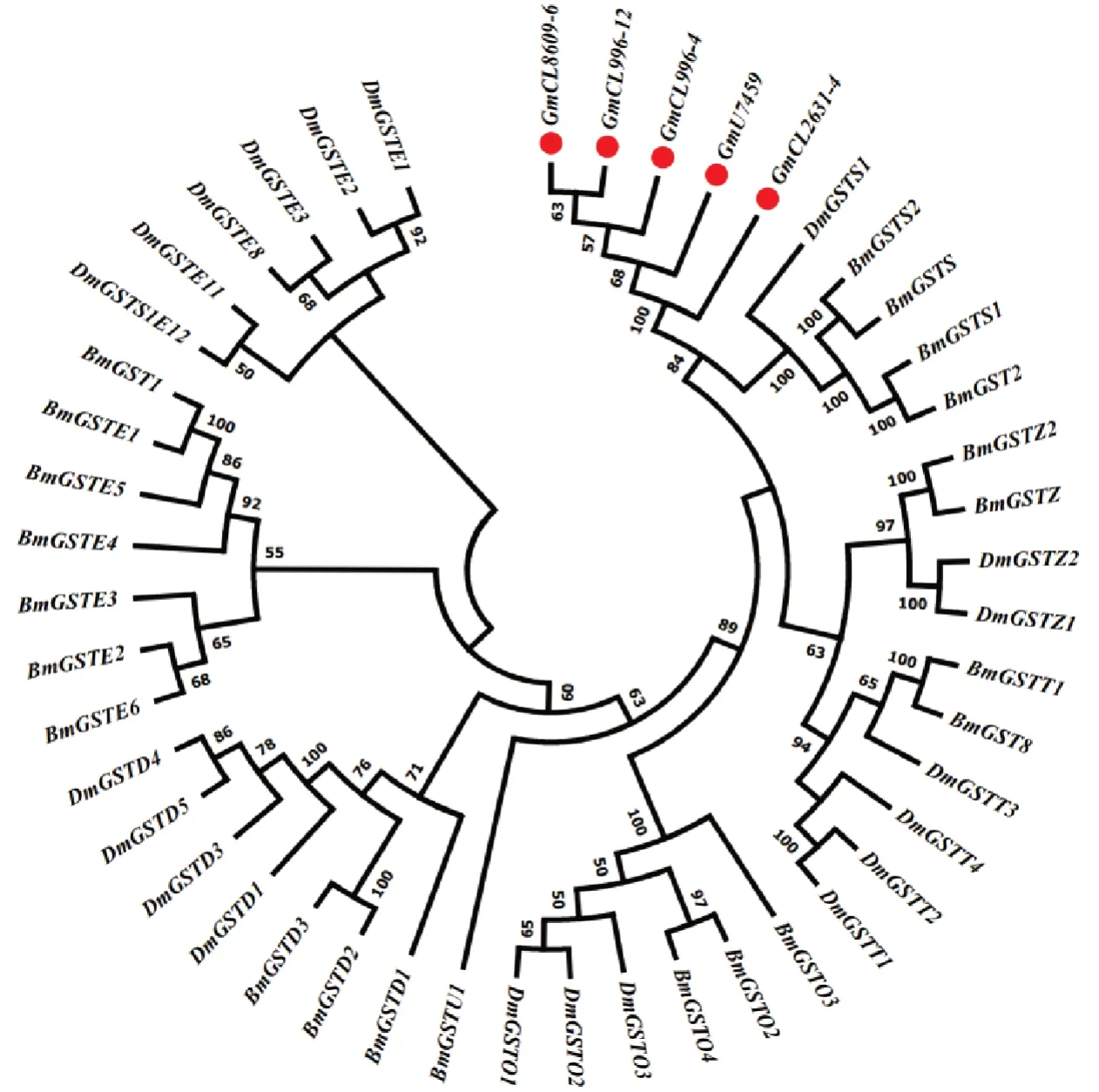
图8 梨小食心虫GST 系统发育分析Fig.8 Phylogenetic analysis of GST in G. molesta
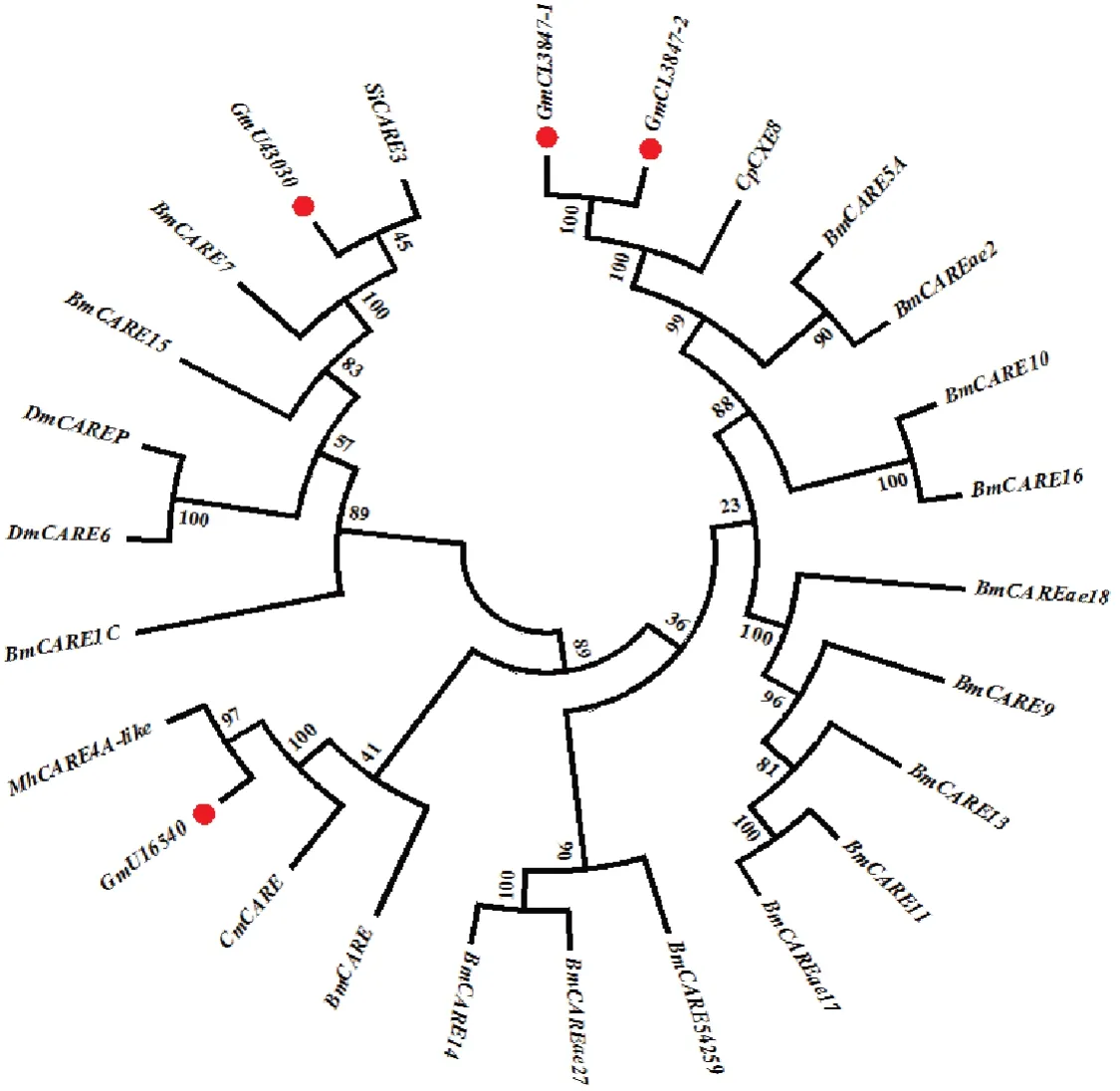
图9 梨小食心虫4 个CarEs 系统发育分析Fig.9 Phylogenetic analysis of four CarEs in G. molesta
3 结论与讨论
梨小食心虫是世界范围内重要的果树害虫之一,幼虫蛀食梨、苹果和桃等的果实以及嫩梢,给林果产业带来了威胁。近年来,许多学者对梨小食心虫进行了深入研究,但针对杀虫剂亚致死剂量下梨小体内差异表达解毒酶基因鲜有报道。本研究通过Illumina Hiseq 2000 测序仪对阿维菌素、高效氯氟氰菊酯、吡虫啉3 种杀虫剂处理后梨小食心虫成虫转录组进行测序和差异表达分析,得到解毒代谢相关基因,为下一步的分子研究奠定基础。本次研究共获得101 873 条Unigene,N50 为3 037 bp,Q20碱基和Q30 碱基的比值分别>96%和>87%。研究表明,Q30 在80%以上就认为测序质量可靠[27]。因此,本研究的测序数据组装质量和长度满足转录组分析的基本要求。
本研究共计有66 658 条Unigenes 被NR、NT、SWISS-PROT 瑞士蛋白质数据库(包括Pfam)、GO、KOG、KEGG 数据库注释,但仍有35 215 条序列没有成功比对,这可能与拼接的序列片段过短、缺乏注释信息有关[28],也可能本身是非编码序列或者是新的某种基因[29]。在NR 数据库中,梨小注释到大蜡螟的序列信息最多,可能与2 种昆虫均属鳞翅目,亲缘关系相对较近有关。梨小转录组Unigenes 在 GO 数据库得到27 486 条功能注释,其中参与催化活性与结合活性的基因功能注释较多,这与其他昆虫转录组结果一致[30]。在KEGG 数据库中得到46 678 条功能注释,其中注释到信号转导通路以及运输和分解代谢通路的基因较多,反映出梨小成虫有较强的代谢活力,为深入挖掘、鉴定解毒代谢基因提供基础数据。
山西农业大学植物保护学院郭艳琼教授课题组前期对未做任何处理的梨小成虫进行了转录组测序,得到77 条CYP 基因、46 条CarEs 基因、28 条GSTs 基因[31],为进一步研究梨小食心虫对杀虫剂的代谢作用提供了宝贵信息。基于此,进行3 种杀虫剂处理的梨小转录组测序分析及差异表达解毒酶基因筛选,得到11 条CYP 基因、4 条 CarEs 基因、5 条GSTs 基因。研究表明,外源性和内源性化合物可以在一些昆虫物种中诱导一些解毒酶基因的表达[32],如黑腹果蝇[33]、小菜蛾[34]、埃及伊蚊[35]等。与未经氯菊酯处理的家蝇相比,暴露于氯菊酯的家蝇多个P450 基因上调[36]。噻虫嗪诱导Q 型烟粉虱GST14的过表达[37]。家蝇CarEs 在氯菊酯处理后可进一步升高[38]。有研究认为,P450s 在昆虫体内的诱导参与了昆虫对环境的适应和对杀虫剂抗性的形成[39]。CYP6A1基因的过度表达使褐飞虱其对吡虫啉的抗性水平提高[40]。吡虫啉诱导后,麦二叉蚜中有6 个属于CYP3 家族的P450 基因表达量升高[41]。阿维菌素作用后,小菜蛾CYP340W1 基因表达量升高,RNA 技术证明此基因参与阿维菌素的代谢[42]。高效氯氟氰菊酯诱导后,斜纹夜蛾CYP301A1、CYP301B1、CYP367A18和CYP304F1可能参与了对高效氯氟氰菊酯的解毒代谢过程[43]。GST 的过表达也增强了杀虫剂的代谢,柑橘全爪螨Panonychus citri中的一个GST 基因(PcGSTm5)在阿维菌素处理下高表达,参与阿维菌素的解毒[44]。杀虫剂对CarE 基因的诱导使冈比亚按蚊[45]、埃及伊蚊[46]、朱砂叶螨[47]等昆虫代谢解毒能力增强。本研究经阿维菌素处理梨小后,有4 个上调的P450 基因、1 个上调的GST 基因;经吡虫啉处理梨小后,有5 个上调的P450 基因、1 个上调的GST 基因;经高效氯氟氰菊酯处理梨小后,有4 个上调的P450 基因、2 个上调的GST 基因、3 个上调的CarEs基因。由此推测,筛选得到的解毒酶基因可能使昆虫解毒能力增强,从而参与对3 种杀虫剂的代谢。
本研究通过对阿维菌素、吡虫啉、高效氯氟氰菊酯3 种杀虫剂不同亚致死剂量处理的梨小食心虫转录组的测序分析以及差异表达基因的筛选,获得与3 种杀虫剂解毒代谢相关基因的信息,为相关基因的克隆、表达、功能研究及进一步研究梨小食心虫解毒代谢机制奠定基础。
