Impacts of Extreme Droughts on Ecosystem Water Use Efficiency Diverge between Forest and Grassland
2023-11-10MengtianHUANGandPanmaoZHAI
Mengtian HUANG and Panmao ZHAI
State Key Laboratory of Severe Weather and Institute of Global Change and Polar Meteorology, Chinese Academy of Meteorological Sciences,China Meteorological Administration, Beijing 100081
ABSTRACT Ecosystem-scale water use efficiency (WUE), as a quantification of the coupling between carbon and water cycle,determines whether vegetation could survive under severe drought stress.Nevertheless, how extreme droughts affect ecosystem-scale WUE and its difference among regions and biomes are still poorly understood.In this study, using data-oriented gridded products of gross primary productivity (GPP) and evapotranspiration (ET), we assessed the ecosystem WUE change during extreme drought years over China, and further compared drought impacts on WUE between forest and grassland ecosystems.We found a spatial heterogeneity of WUE change in response to extreme droughts across regions.Particularly, grassland WUE was substantially reduced concurrent with suppressed photosynthesis, while most of forest exhibited slightly decreased or even increased WUE under extreme droughts.In addition, we demonstrated that drought characteristics and environmental conditions modulated drought impacts on WUE.Temperature anomalies during droughts and leaf area were found to be the predominant factors driving WUE change for both forest and grassland.With increasing occurrence of compound dry and hot extremes, our results,therefore, would be an insightful supplement to the current understanding of the influence of extreme events on terrestrial ecosystems.
Key words: extreme droughts, water use efficiency, terrestrial ecosystems, forest, grassland, China
1.Introduction
In the context of global warming, climate and weather extremes have been identified by the World Climate Research Programme as one of the grand challenges(Zhang, 2014; Sillmann et al., 2017).Droughts are well recognized to be one of the most significant extreme climate events with a variety of impacts on natural and social ecosystem, water storage, and human life (Huang et al., 2020; Pokhrel et al., 2021; Vogt et al., 2021).In China, previous studies have reported that droughts have caused profound changes in ecosystems structure and function as well as land processes (Deng et al., 2021).Particularly, extreme droughts have produced some of the most severe impacts on terrestrial vegetation, leading to widespread reduction in vegetation productivity (Zhao and Running, 2010; Schwalm et al., 2012; Li et al., 2019;Deng et al., 2021), growth retardation (Huang et al.,2018), and even plant mortality (Eamus et al., 2013).It thus calls for an accurate assessment on the influence of extreme droughts on terrestrial ecosystems and a deeper understanding of the underlying processes.
Facing with serious environmental stresses especially extreme severe water deficit, whether vegetation could survive is largely determined by its capacity to utilize the available water to maximize photosynthetic production and tissue maintenance (Huang et al., 2015, 2021).In this case, the concept of water use efficiency (WUE) is proposed as an indicator to reveal coupling relationship between plant photosynthesis and transpiration, reflecting the water use strategy of plant individual in response to change in ambient conditions (Yu and Wang, 2010;Medlyn et al., 2017).In arid and semiarid regions, especially, relatively higher WUE plays an important role for plant survival and individual growth under severe water deficit (Linares and Camarero, 2012).At ecosystem scale, WUE provides a quantification of the coupling between carbon and water cycle, or in other words, ecosystem productivity and water loss (Keenan et al., 2013;Ponce-Campos et al., 2013; Tang et al., 2014; Huang et al., 2015, 2016).Therefore, it is becoming vital to explore the drought impacts on ecosystem WUE and the underlying mechanisms, which is of great significance to revealing the response of terrestrial ecosystems to climate change and its extremes.
Considering the rare occurrence of extreme drought events, the current understanding regarding the impacts of extreme droughts on ecosystem carbon-water coupling is still limited.In most of the existing studies, the impacts of extreme droughts on ecosystem WUE have so far been analyzed mainly for specific drought years as local- or regional-scale case studies in China due to the constraints of available data in terms of either spatial coverage or time span (Liu et al., 2015; Xu et al., 2019;Wang et al., 2021).For example, Liu et al.(2015) selected the years of 2001, 2006, 2009, and 2011 from the period of 2000-2011 as the driest four years in China and examined change in WUE under drought based on a process-based ecosystem model.Although they concluded that extreme droughts reduced while severe drought slightly increased annual WUE, it should be noted that the “extreme/severe” droughts were defined empirically rather than based on the magnitude of drought indices.Xu et al.(2019) focused on northern China and investigated the response of WUE to drought using satellite-derived data.By selecting the top three regional drought events in 2001, 2005, and 2014 as cases based on the standardized precipitation evapotranspiration index(SPEI), they detected that severe and extreme drought generally resulted in decreased ecosystem WUE regardless of different hydroclimatic conditions.In a more recent study, Wang et al.(2021) examined the impacts of the 2009/2010 extreme seasonal droughts on ecosystem WUE in Southwest China and found that divergent responses of ecosystem WUE to extreme seasonal droughts could be attributed to different sensitivities of ecosystem carbon and water cycle to drought.Combining flux site observations and global gridded datasets, Yang et al.(2016) provided a synthesis of drought effect on ecosystem WUE at the global scale based on linear correlation analysis.They evidenced the influence of hydroclimatic conditions on the response of WUE to drought, but unfortunately, provided hardly any information in terms of extreme droughts.In such cases, how extreme droughts affect ecosystem WUE across a broader range remains for further investigation.
More importantly, given different ambient conditions(e.g., background climate and nutrient availability), community composition, and physiological process between ecosystems (Chapin et al., 2011), drought impacts on ecosystem carbon and water dynamics may diverge among different biomes or hydroclimatic conditions.In this regard, previous works have shown evidence using in-situ flux tower records, satellite-derived vegetation indices, flux sites upscaling estimations, and ecosystem model simulations (Zhang et al., 2016; Xu et al., 2018;Deng et al., 2021).For instance, Deng et al.(2021) observed that vegetation greenness and productivity of forest was less sensitive to droughts compared with grassland across China.In addition, forest has been found to be more resistant to meteorological (Xu et al.,2019) and soil moisture (Ji et al., 2021) droughts than shrubland, grassland, and cropland.Nevertheless, so far,little information is known about whether and how the impacts of extreme droughts might differ among biomes,as well as the possible drivers.
With this end in view, this study aims to provide the spatial pattern of changes in ecosystem WUE during extreme droughts across China.We hypothesized that the impacts of extreme droughts on WUE will be divergent between forest and grassland ecosystems.The overall objective of this study is to address the following questions:(1) How extreme droughts affect ecosystem WUE at a national scale? (2) Whether and how the impacts of extreme droughts differ between forest and grassland? (3)What are the major drivers determining the change in WUE in response to extreme droughts? Achieving these objectives will help to improve our understanding and predictions of the response of ecosystem carbon-water coupling to global climate change.
2.Data and methods
2.1 Drought metric and extreme drought year detection
To accurately and objectively quantify drought characteristics including intensity, magnitude, duration, and spatial extent, plenty of attempts have devoted to developing approaches to defining drought indices for drought monitoring and analysis (Vicente-Serrano et al., 2010a).To date, the most widely used drought indices include the Palmer drought severity index (PDSI), the standardized precipitation index (SPI), and SPEI.
In this study, SPEI was used considering its advantages that: (1) it is calculated based on the difference between precipitation and potential evapotranspiration so as to account for the potential influence of temperature variability and temperature extremes on drought assessment; (2) its multiscalar nature enables large-scale drought detection and monitoring through time and space(Vicente-Serrano et al., 2010a, b).The global 0.5° gridded SPEI dataset was obtained from the Global SPEI database (SPEIbase v2.7, https://spei.csic.es/spei_database_2_7), spanning the period from January 1901 to December 2020.To represent the long-term drought condition or annual drought, SPEI with a time-scale of 12 months (SPEI-12) was utilized by following previous studies (Hamal et al., 2021).Across the study area and for each year, the average SPEI-12 from May to September was calculated to derive the time series of annual growing season (GS) mean SPEI-12 as this study aims to detect extreme drought years.On this basis, extreme drought years were defined as years with GS-mean SPEI-12 value less than -1.5.
2.2 Ecosystem WUE definition
At ecosystem scale, WUE is generally estimated as the ratio of ecosystem measures of gross primary productivity (GPP) to evapotranspiration (ET) (Huang et al., 2015,2016; Xu et al., 2019), representing the trade-off between carbon assimilation and water loss of terrestrial ecosystem (Huang et al., 2015, 2016).In this study, global gridded datasets of GPP and latent heat (LE) were downloaded from the Biogeochemical Integration of Max Planck Institute (MPI) (www.bgc-jena.mpg.de/geodb/projects/Data.php).GPP/LE measurements from FLUXNET eddy covariance towers were merged with remote sensing and meteorological data by using machine learning approach to generate the global estimates (Jung et al., 2011).Both datasets have a 0.5° × 0.5° spatial resolution and a monthly temporal resolution from 1982 to 2011, and are suggested to be the data-oriented GPP/LE products with the longest time span so far and global continuously spatial coverage.Following Tang et al.(2014), LE was converted to ET units based on the formula ET = LE/λ (λ =2454 J g-1).Monthly GPP/ET values were aggregated over the period of May to September each year to obtain annual GS GPP/ET values for annual GS WUE calculation.
Previous studies have pointed out a nonlinear effect of atmospheric vapor pressure deficit (VPD) on WUE at both leaf and ecosystem scale (Baldocchi et al., 1987;Tang et al., 2006; Yu and Wang, 2010; Linderson et al.,2012).As ecosystem ET is comprised of canopy transpiration, soil evaporation, and canopy interception evaporation (Huang et al., 2015), the balance between vegetation carbon assimilation and water loss can be confounded by VPD-induced increase in atmospheric evaporative demand (Zhou et al., 2014; Huang et al., 2015; Zhang et al., 2019).Therefore, inherent water use efficiency(IWUE) was introduced to exclude the potential VPD impacts via the abiotic increase in evaporative potential(Beer et al., 2007, 2009) with a focus on the intrinsic vegetation response.Based on this, IWUE was also calculated in this study as the product of VPD and WUE, that is, GPP × VPD/ET.Corresponding to the GS mean WUE calculation, monthly VPD values were first averaged during May-September each year to obtain GS mean VPD and then used in GS mean IWUE calculation.
2.3 Meteorological data
Monthly temperature, precipitation, potential evapotranspiration, and air vapor pressure data were derived from the University of East Anglia’s Climatic Research Unit (CRU) via https://crudata.uea.ac.uk/cru/data/hrg/cru_ts_4.06/.CRU TS v4.06 dataset (Harris et al., 2020)covers the period of 1901-2021 at a spatial resolution of 0.5° × 0.5°.The average temperature/air vapor pressure from May to September each year was calculated as the GS mean temperature/air vapor pressure, and the sum precipitation during May-September each year was calculated as the GS total precipitation.The gridded temperature and solar download radiation data during 1901-2013 were obtained from the CRU/NCEP version 5 6-h dataset with a spatial resolution of 0.5° × 0.5°.Daily average/maximum temperature was then defined as the mean/maximum value for the daily 6-h temperature values.
In this study, VPD was calculated as the difference between air saturated vapor pressure (SVP) and air vapor pressure (AVP) following the approach by Zhao and Running (2010).Because photosynthesis and transpiration mainly occur in daytime with sunlight for most plants, thus daytime VPD was calculated and used in the analysis.To this end, SVP (in Pascal) was defined as a function of air temperature (in Celsius) [Eq.(1)]:
whereTis the average air temperature during daytime(Tday).Here,Tdaywas estimated based on the logic of Mountain Microclimate Simulation Model (MT-CLIM)as Eq.(2) (Running et al., 1987; Zhao and Running,2010):
whereTmaxis the daily maximum temperature andTavgis the daily average temperature, which were both calculated by using CRU/NCEP dataset.For AVP, assuming(1) there is little diurnal changes in air pressure and (2)the amount of water molecular is relatively stable during a day, the daytime AVP therefore could be directly estimated by using the average AVP (Zhao and Running,2010), which was derived from CRU dataset.
2.4 Vegetation data
The normalized difference vegetation index (NDVI)data was obtained from the Global Inventory Modelling and Mapping Studies (GIMMS) third generation product(NDVI3g) at a spatial resolution of 8 km × 8 km from 1982 to 2016 (Pinzon and Tucker, 2014).In this study,only pixels with a mean annual NDVI larger than 0.1 were defined as the vegetated region and included as the study region.The leaf area index (LAI) from GIMMS was generated by an artificial neural network model and GIMMS NDVI3gat the 8 km × 8 km spatial resolution from 1982 to 2016 (Zhu et al., 2013).In this study, both NDVI and LAI were resampled to a 0.5° × 0.5° spatial resolution to match other datasets in the analysis.
The vegetation distribution map was derived from the 1 : 1,000,000 digitalized vegetation map of China (Editorial Committee of China Vegetation Map, Chinese Academy of Sciences, 2007).Within the study region defined by NDVI threshold, the current vegetation types were combined and reclassified into six major categories of forest, shrubland, desert, grassland, wetland, and cropland (Fig.S1).This study has a special focus on forest and grassland, accounting for 19% and 30% of the study area, respectively.Although cropland also covers about 34% of the study area, it is excluded in this analysis considering that the strong human disturbance and land management activities over these regions may confound the influence of climatic variabilities.
2.5 Analysis
During 1982-2011 (the overlap period of all data used in this study), the standardized anomaly (SA) of WUE was calculated for each grid cell and each extreme drought to quantify WUE change in response to extreme drought:
wherexi,tis the GS WUE value in theith year,xi,tand σ(x i,t)are the average and standard deviation of GS WUE over the study period, respectively.Correspondingly, SA of GS GPP, GS mean temperature, and GS total precipitation were also calculated to assess changes in vegetation productivity and climate conditions during extreme drought years.The long-term trend was removed from the time series of each variable by using the linear regression method before calculating SA.To further explore the relationship between WUE responses to drought and climate variables, the values of SA of WUE were averaged in the climate space with 1°C intervals of mean annual temperature and 100-mm intervals of mean annual precipitation.When comparing WUE changes in drought for forest and grassland, the analysis of variance(ANOVA) was performed to determine whether there is a statistically significant (p< 0.05) difference between these two vegetation types.In addition, within each biome,we further compared GPP in extreme years and GPP in normal years for each pixel.Considering the limited sample size of pixels experiencing extreme drought years during the study period, Mann-WhitneyUtest was performed to determine whether GPP values are statistically significantly different between drought and normal years.
To investigate the possible drivers determining the spatial variations of WUE response to drought, boosted regression tree (BRT) approach, a machine-learning technique extended from traditional classification and regression trees, was employed using the “dismo” and “gbm”packages (Elith et al., 2008) in the R statistical software(version 4.2.2).Eight variables were used to identify their relative influence to WUE change (indicated by SA of WUE), including three variables indicating drought characteristics (drought intensity, SA of GS mean temperature in extreme drought year, and SA of GS total precipitation in extreme drought year), and five variables indicating background environmental conditions (mean annual temperature, mean annual precipitation, mean annual solar radiation, mean annual GS LAI, and mean annual aridity).Here, drought intensity was indicated by the magnitude of SPEI value for each extreme drought year;aridity was indicated by the Aridity Index (AI) calculated as the ratio of average annual precipitation amount to potential evapotranspiration amount based on CRU dataset (Abatzoglou et al., 2018).The model was fit to allow interactions by using a tree complexity of 10 and a learning rate of 0.01.
3.Results
3.1 SPEI-based extreme droughts in China
Across the vegetated area in China, regions experiencing extreme drought years during 1982-2011 cover about more than 80% of the study area.About 25% of the study area witnessed more than three extreme drought years, mainly in North and Northeast China, as well as parts of South China (Fig.1a).Particularly, five or more extreme drought years are detected in the eastern Inner Mongolia (Fig.1a).In terms of drought intensity, more severe droughts (i.e., with the lowest SPEI value) are found in Xinjiang, Southwest China especially Yunnan, Southeast China, and parts of eastern and northern Inner Mongolia (Fig.1b).Note that in the following analysis, we only focused on regions where extreme drought years have been detected during the study period.
During these extreme drought years, a vast area of decrease in GS precipitation is observed, among which shows about 34% dry anomalies of GS precipitation exceeding 1.5 standard deviations (SD) (Fig.1c).The most remarkable precipitation reductions during droughts are mainly located in the Tibetan Plateau, Northeast and Central China, and parts of South China (Fig.1c).Meanwhile, nearly 30% of the region with detected droughts exhibits higher than normal temperature (> 0.5 SD) during extreme drought years (Fig.1d).Notable hot anomalies (> 1.5 SD) can be seen in central Northeast China,Southwest China, central Inner Mongolia, and parts of Central and North China (Fig.1d).
3.2 Drought impacts on ecosystem WUE
In Fig.2, we calculated WUE changes in response to extreme droughts indicated by the mean SA of WUE in extreme drought years.The results show a significant decrease in WUE (exceeding -1 SD) under extreme drought conditions in about 30% of the study area, in particular in most parts of North and Northeast China,Xinjiang, the Tibetan Plateau, and Yunnan Province (Fig.2a).Increase in WUE above normal ranges (exceeding 1 SD) under extreme drought conditions are only found in parts of South China, accounting for less than 5% of the study area (Fig.2a).To remove the effects of VPD on evaporative potential, we also examined IWUE change during extreme drought years.As shown in Fig.2b, the distributions of WUE and IWUE change in extreme droughts have very similar spatial patterns, but with smaller negative IWUE anomalies in Northeast China and parts of Yunnan Province.Consequently, the spatial pattern of change in ecosystem productivity (indicated by GPP) during extreme drought years generally depends onthat of WUE change.Increased ecosystem GPP mainly occurs in South China, while decreased ecosystem GPP below normal ranges (exceeding -1 SD) are observed in most of other parts of the study area (Fig.2c).
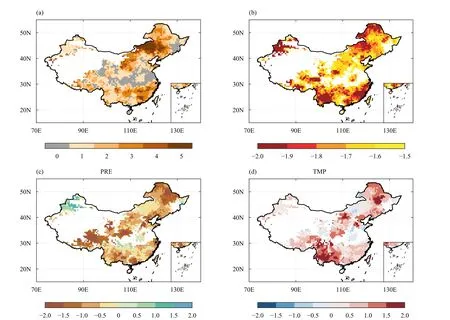
Fig.1.Characteristics of extreme drought years identified by SPEI.Shown are (a) drought frequency during 1982-2011, (b) drought intensity indicated by the magnitude of growing season mean SPEI-12 value, (c) the average standardized anomalies of growing-season precipitation in extreme drought years, and (d) the average standardized anomalies of growing season temperature in extreme drought years.Note that only the vegetated area where extreme drought year was detected during the study period are shown in panels (b-d).
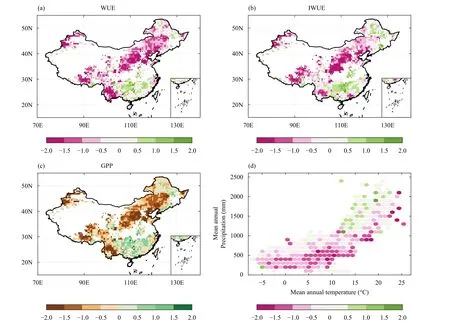
Fig.2.Drought impacts on ecosystem WUE and productivity.Shown are (a) the spatial pattern of the standardized anomalies (SA) of WUE, (b)IWUE, and (c) gross primary productivity (GPP) during extreme drought years.Only the vegetated areas where extreme drought years were detected during the study period are shown in panels (a-c).The SA of WUE in the climate space is shown in panel (d).Each climate bin is defined by 1°C interval of mean annual temperature (MAT) and 100-mm interval of mean annual precipitation (MAP) averaged between 1982 and 2011.
Then we investigated the relationship between WUE change under extreme droughts and background climate(Fig.2d).Generally, WUE response to extreme droughts has no clear relation to spatial gradient of temperature,but is more closely related to precipitation conditions.Negative WUE anomalies in extreme droughts are mainly located in regions with mean annual precipitation lower than 1000 mm, except for a tiny minority in the warmest regions of the study area.Positive WUE anomalies in extreme droughts are mostly found in regions where mean annual precipitation is higher than 1000 mm.
3.3 Divergent WUE responses between forest and grassland
To investigate whether and how the impacts of extreme drought on ecosystem carbon-water coupling differ across biomes, we further compared change in WUE and IWUE during extreme drought years between different biomes in China, taking forest and grassland as an instance.We show that extreme drought has a significantly (p< 0.05) stronger negative effects on grassland WUE than on forest WUE (Fig.3).On average, extreme drought years have resulted in mean negative WUE anomalies of -0.751 and IWUE anomalies of -0.639 for grassland ecosystems.By comparison, forest ecosystems show negative WUE anomalies of -0.128 in extreme drought years, and even a slightly increase in IWUE under drought stress (0.121).
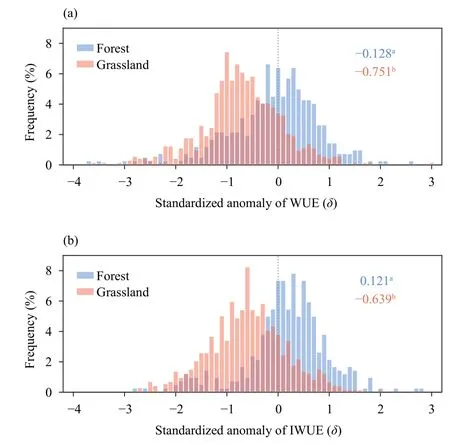
Fig.3.Change in ecosystem WUE in extreme drought years for forest and grassland.Numbers on the right side in each panel refer to the mean standardized anomalies (SA) of (a) WUE and (b) ET anomalies for forest and grassland, respectively.Different letters indicate statistically significant differences among drought timing at p < 0.05 based on analysis of variance (ANOVA).
Combining changes in GS GPP and GS ET during extreme drought years, we observed divergent underlying processes of ecosystem WUE to extreme droughts between forest and grassland (Fig.4).Figure 3 suggests that forest ecosystems on average experienced slight WUE and IWUE variations in extreme drought years.(14.9%) compared to that of other factor considered in this study (Fig.5a).SA of GS temperature during extreme drought years (SATMP) is the second major driver recognized here, with a relative influence of 14.6% and followed by mean annual GS LAI (13.7%).By contrast,mean annual precipitation (MAP), as a background climate condition, contributes the least (8.9%) to droughtinduced WUE change for forest ecosystem.For grassland, SATMPhas a strongest influence (15.2%) on WUE change under extreme droughts, followed by MAP(13.2%) and mean annual GS LAI (13.0%; Fig.5b).Although AI is found to be the predominant driving factors of WUE change for forest, it contributes the least(10.3%) to WUE change for grassland compared with other factors.
4.Discussion and conclusions
Meanwhile, there seems no statistically significant difference when comparing forest GPP between extreme drought and normal years (Fig.4a).We further show in Fig.4c that regions with no statistically significant difference cover about 84% of forest ecosystems.For grassland, however, drought-induced WUE and IWUE decreases are accompanied with significant reductions in GS GPP compared to normal years (Fig.4b).For nearly 80% of grassland ecosystems in the study area, the average GS GPP is lower in extreme drought years than in normal years, among which 34% is statistically significant (p< 0.1) (Fig.4d).
3.4 Possible drivers of WUE change under extreme droughts
To further explore the possible reasons for the divergent drought-induced WUE change for forest and grassland, we further compared the contribution of driving factors regarding drought severity and background environmental variables to WUE change under extreme drought for these two biomes, respectively.Interestingly,the results did reveal a remarkable difference in terms of the major driving factors of WUE response towards extreme droughts between forest and grassland.
To be specific, for forest, mean annual aridity (AI) is identified as the primary driver of WUE change under extreme droughts, having the highest relative influence
4.1 Different drought impacts on WUE across regions and biomes
In this study, we examined the impacts of extreme droughts on ecosystem carbon-water coupling over China.To address this question, two definitions of ecosystem WUE, i.e., WUE and IWUE, were used in our analyses.Generally, we found that IWUE, which excludes the effects of change in atmospheric evaporative demand, shows a similar manner in response to extreme droughts with WUE.This suggests that when drought intensity exceeds a certain hydroclimatic threshold (Ponce-Campos et al., 2013), the adjustments of ecosystem physiology in response to the changing environment would overwhelm those of land surface physical processes such as VPD-induced soil water loss (Beer et al.,2009), eventually dominating the change in ecosystem WUE under extreme droughts.
Further, we found a spatial heterogeneity of WUE change during extreme drought years across the study area (Fig.2).Additionally, WUE of grassland generally reduces in extreme drought years, while most of forest exhibits slightly decreased or even increased WUE under extreme droughts (Fig.3).These results suggest that the ecosystem WUE of forest is relatively more resistant to extreme droughts than that of grassland, which is in line with previous studies despite most of them focusing on general droughts instead of extreme drought events(Xu et al., 2019; Ji et al., 2021).
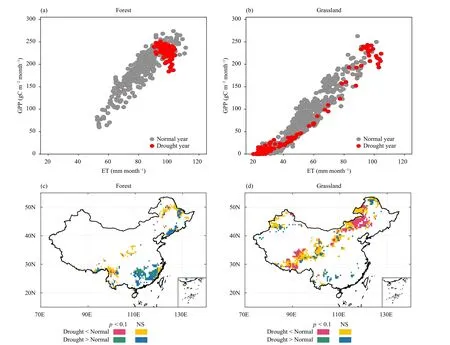
Fig.4.Changes in GPP and evatranspiration (ET) in extreme drought years for forest and grassland.In panels (a) and (b), each dot refers to a pixel in the region of a certain biome, with those in gray for normal years and those in red for extreme drought years detected during the study period.Shown is the average monthly GPP/ET during the growing season (GS; May-September).Panels (c) and (d) show the comparison between the average GS GPP in extreme drought years and that in normal years for forest and grassland, respectively.Statistical significance was determined using Mann-Whitney U test with “NS” indicating statistically insignificance.
Particularly, we showed that GPP of forest shows no obvious difference between during extreme drought years and during normal years, so does forest WUE(Figs.4a, c).This phenomenon can be explained by the fact that the carbon assimilation process of forest ecosystems is less likely to be negatively affected by extreme droughts, that is, forest could maintain its intrinsic photosynthesis capacity under drought stresses.In supporting of our finding, Deng et al.(2021) also observed that forest biomes are the least sensitive to extreme droughts compared with other biomes in China in terms of either vegetation greenness or vegetation photosynthesis.Such drought resistance of forest ecosystem is probably because forest, with extensive root system, can absorb available water to the full extent from deeper soil layer to relieve drought stress (Chapin et al., 2011; Xu et al.,2018).In this case, vegetation photosynthetic capacity of forest tends to remain relatively stable, unless extreme droughts lead to widespread tree mortality (Anderegg et al., 2013).On the basis of IWUE, which excludes the effects of bare soil and canopy ET, we did detect enhanced IWUE for forest under extreme droughts.This is consistent with Zhang et al.(2019) showing increased ecosystem intrinsic WUE for forests under extreme hydrological stress induced by increasing VPD.
On the contrary, GPP of grassland is widely suppressed in extreme drought years compared with normal years (Figs.4b, d), concurrent with substantial reduction in both WUE and IWUE.Such inhibition by extreme droughts on grassland WUE has also been suggested by Zhang et al.(2019).On the one hand, extreme droughts can lead to a considerable reduction in grassland productivity due to its shallow rooting system (Deng et al.,2021).On the other hand, with relatively lower vegetation coverage, evaporation from bare soil tends to be largely enhanced due to increase in atmospheric evaporative demand during droughts.Therefore, significant decrease in ecosystem WUE is observed for grassland in response to extreme droughts.
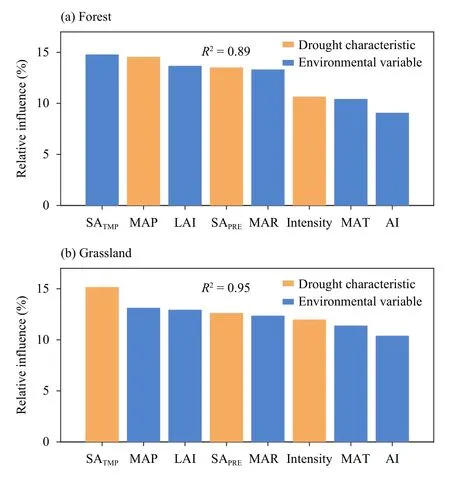
Fig.5.Average relative influence (%) of drought-related and environmental variables to WUE change in extreme drought years for forest and grassland.Variables are ranked in decreasing order based on average relative influence based on boosting regression tree analysis.Orange bars represent variables regarding drought characteristics, including drought intensity (Intensity), SA of GS temperature (SATMP), and SA of GS precipitation (SAPRE).Blue bars represent variables indicating background environmental conditions, including mean annual temperature (MAT), mean annual precipitation (MAP), mean annual solar radiation (MAR), mean annual GS LAI (LAI), and mean annual aridity (AI).In each panel, R square value in top right refers to the percentage of which SA of WUE in extreme drought years could be explained by these variables considered.
4.2 Contribution of drought characteristic and background environment to WUE change under extreme droughts
Based on comparison of the relative influence of drought-related and environmental variables on WUE change under droughts, we found that for both forest and grassland, the SA of GS temperature (SATMP) during extreme drought years acts as top-drawer drought-related factors driving WUE response to extreme droughts.Further analysis shows a negative relationship between SATMPand SA of WUE for both forest and grassland ecosystems (Fig.S2).Obversion-based studies have suggested that drought directly decreases vegetation photosynthesis by decreasing CO2supply with stomatal closure (Konings and Gentine, 2017) and/or by decreasing the activity of photosynthetic enzymes (Chaves et al., 2009; Misson et al., 2010), especially when heat stress compounds drought impacts.Besides, even though plant transpiration may also decrease as a result of stomatal closure,this is largely offset by increase in bare soil and canopy evaporation with temperature increases.In this case, relatively low ecosystem WUE is observed as GPP decreases and ET increases.
Interestingly, we also observed that LAI, as an indicator of vegetation state and canopy structure, has an important influence on WUE change that cannot be ignored.LAI, which characterizes the mass (e.g., water and CO2)and energy exchanges among soil, vegetation, and atmosphere, has widely been suggested to influence canopy physiological and biochemical processes including photosynthesis and transpiration (Zhu et al., 2013).We further found that both forest and grassland show more serious WUE reduction, i.e., larger negative SA of WUE, in regions with lower mean annual GS LAI (Fig.S2).Based on CMIP5 climate models, Lian et al.(2018) revealed that LAI is the primary driver of spatial variations of the ratio of plant transpiration to total terrestrial evapotranspiration (T/ET), which presents the role of vegetation in surface-atmosphere interactions (Lian et al., 2018).In this way, low LAI suggests spare vegetation coverage over a certain region, especially for grassland.Lower WUE, therefore, is more likely to be observed because of the larger contribution of dramatic increase in soil evaporation to the total amount of ecosystem water loss when drought occurs.
4.3 Uncertainties and limitations
In our analyses, gridded datasets of SPEI, GPP, ET,and meteorological data at a spatial resolution of 0.5° ×0.5° were used to investigate drought impacts on ecosystem WUE.However, there remained some uncertainties to be carefully considered.On the one hand, such a spatial resolution could be too coarse to accurately measure the drought impacts on WUE in regions with complex terrain, such as in high mountains and the Tibetan Plateau.On the other hand, relatively coarse resolution has the mixed pixel problem (Xu et al., 2019), but it is difficult to validate against observation-based results.Accordingly, further investigations are required to compare results based on gridded datasets with studies into the response of WUE to extreme droughts among ecosystem types based on in-situ observations.Therefore, these data uncertainties should be taken into account when interpreting our results.Apart from the uncertainties in the data, other factors should also be considered.For example, land use and land cover change were not considered in our study, which might lead to uncertainties and should be further examined.
Meanwhile, the drivers of WUE change under extreme droughts can also be spatially heterogenous.Nevertheless, due to the limited sample size, the BRT analysis in this study was only conducted by taking forest and grassland as study area, respectively.For a better understanding of the predominant factors driving the WUE change across regions and other biomes in addition to forest and grassland, more in-depth studies associated with in-situ observations, manipulation experiments, and model simulations are much needed.
By combining SPEI and gridded datasets of ecosystem GPP and ET, we examined the spatial pattern of change in ecosystem WUE in response to extreme droughts across China.Our results highlight divergent impacts of extreme droughts on ecosystem WUE between forest and grassland, which implies that land use and land cover change should be fully considered in analyses regarding the influences of climate change and extreme events on WUE to reduce the uncertainties.Further, a latest investigation has indicated different drought resistance between genus of forest or gymnosperms and angiosperms due to their variations in eco-physiological traits (Li et al., 2020).Considering the variety of forest and grassland species across China, more detailed studies are still needed for a better understanding of the spatial variation in WUE change under extreme droughts.In addition, the expected magnitude of drought impacts is not merely determined by drought intensity (Liu et al.,2015), but also by drought timing (Huang et al., 2018;Deng et al., 2023), as well as its interactions with drought magnitude, duration, and frequency.This requires more observation-based investigation on the underlying processes of how droughts affect ecosystem carbon and water cycles.Last but not least, although the influences of temperature variability and temperature extremes have been taken into consideration in SPEI calculation (Vicente-Serrano et al., 2010a, b), we found that the magnitude of drought impacts on ecosystem WUE is associated with temperature variability during extreme drought years.That is to say, warm spells concurrent with severe water deficit amplifies the negative impacts of extreme droughts on terrestrial vegetation.Given this, compound dry and hot events, which have occurred frequently in the past decade (Feng et al., 2020; IPCC, 2021), may have non-linearly increased impacts on natural and human systems compared to single extreme events, which is, unfortunately, poorly understood so far.Our study, therefore,suggests further experiments and observational designs at different scales to verify such amplified impact and quantitatively compare its magnitude with that of single drought or heat events.
杂志排行

Journal of Meteorological Research的其它文章
- Changes in Atmospheric Circulation during the Winter Regional Extreme Cold Events over China since 1960
- Changes in Persistent Precipitation in Northwest China and Related Large-Scale Circulation Features
- Variation of Dust in Northern China and Its Reproduction in BCC-ESM1 since 1980
- Predictability and Risk of Extreme Winter PM2.5 Concentration in Beijing
- Operational Plan, Effect Verification, and Key Technical Settings for a Stadium-Scale Artificial Rain Reduction Experiment
- Random Forest-Based Snow Cover Mapping in China Using Fengyun-3B VIRR Data