西方蜜蜂Sirtuin蛋白家族基因鉴定及表达分析
2023-10-31宋文菲苗春辉余玉生胡宗文王艳辉李亚辉
宋文菲,刘 娜,苗春辉,余玉生,胡宗文,王艳辉,李亚辉
(1. 云南农业大学 动物科学技术学院,昆明 650201; 2. 云南省农业科学院 蚕桑蜜蜂研究所,云南蒙自 661101;3. 红河学院 生物科学与农学学院,云南蒙自 661199)
沉默信息调节因子2(Silent information regulator2,Sir2)是依赖于烟酰胺腺嘌呤二核苷酸(NAD+)的Ⅲ类组蛋白/蛋白去乙酰化酶[1-2]。已知Sir2样蛋白均含有一个去乙酰化酶(Sirtuin)核心结构域,因而将Sir2同源的蛋白统称为Sirtuin蛋白[3-4]。该蛋白家族广泛存在于原核生物和真核生物中,在基因表达、细胞凋亡、新陈代谢和衰老等细胞过程中发挥了关键作用[5-7]。研究发现生物的Sirtuin蛋白基因分为5类,Ⅰ类包含3个基因(Sirt1~Sirt3),Ⅱ类和Ⅲ类各包含1个基因,分别为Sirt4和Sirt5基因,Ⅳ类包含2个基因(Sirt6和Sirt7),U类包括古细菌和细菌Sirtuin基因[6-7]。不同生物间Sirtuin蛋白基因数和分布存在差异,例如原核生物的枯草芽孢杆菌Bacillussubtilis、粪肠球菌Enterococcusfaecalis属于U类,仅有1种[6-7];真核生物的酿酒酵母Saccharomycescerevisiae有4种,均属于Ⅰ类;黑腹果蝇Drosophilamelanogaster有5种,属于Ⅰ、Ⅱ和Ⅳ类;西方蜜蜂Apismellifera有6种,哺乳动物人类有7种,均属于Ⅰ~Ⅳ类[6-8]。
目前Sirtuin蛋白功能已在许多真核生物中相继得到验证,例如在酿酒酵母中Sirtuin蛋白参与端粒、rDNA和沉默交配型位点的沉默,维持基因组完整性和寿命调控[9-10],Sir2基因缺失会导致酿酒酵母寿命缩短,而增加Sir2p基因拷贝可延长酵母寿命[10-11]。Sir-2.1和dSir2基因表达增加可分别延长秀丽隐杆线虫Caenorhabditiselegans和黑腹果蝇的寿命[11-12],黑腹果蝇和酿酒酵母的Sirtuin蛋白可通过热量限制延长生物体寿命[12-13]。在哺乳动物中,Sirtuin蛋白家族成员有7个,Sirt1、Sirt6和Sirt7定位于细胞核,Sirt2定位于胞质,Sirt3、Sirt4和Sirt5定位于线粒体[14]。 Sirt1对许多转录调控的关键组蛋白(H3-K9、H4-K16和H1-K26),以及非组蛋白靶点(p53、FOXO1/3和PGC-1α)有去乙酰化作用,在DNA修复与凋亡、肌肉与脂肪分化、线粒体生物发生、葡萄糖与胰岛素稳态和激素分泌中发挥了重要作用[14]。哺乳动物组织中的Sirt1蛋白水平会被禁食和热量限制所诱导,Sirt1基因在大脑过量表达可延长小鼠寿命[15-16];Sirt2通过调控H4K20甲基化的有丝分裂沉积,进而控制细胞周期进程和基因组稳定性,并间接参与神经退行性疾病的自噬、氧化应激和炎症等细胞过程[17];Sirt3可调节线粒体蛋白的整体乙酰化,控制线粒体氧化途径的流动,减少氧化应激,在造血干细胞中高度富集,在衰老造血干细胞中上调可提高其再生能力[18-19];Sirt4是DNA损伤反应通路的重要组成部分,协调谷氨酰胺代谢中的代谢阻滞、细胞周期阻滞和肿瘤抑制[20]; Sirt5是一种依赖于NAD+的赖氨酸脱酰基酶,可去除琥珀酰和丙二酰,以糖酵解为主要靶标调节细胞溶质和线粒体的蛋白丙二酰化[21]; Sirt6通过调节DNA修复、端粒维持以及葡萄糖和脂质代谢来影响细胞稳态,其在转基因小鼠的过量表达可延长雄性小鼠寿命[22-23];Sirt7在染色质调控、细胞转化程序和肿瘤形成中起到关键作用,通过调控泛素-蛋白酶体途径控制肝脏的脂质代谢[24-25]。
目前,国内外学者已对哺乳动物Sirtuin蛋白功能开展了系统研究,并发现Sirtuin蛋白在哺乳动物代谢相关疾病和延缓衰老的调控作用[14-16],针对昆虫Sirtuin蛋白功能研究仅集中在黑腹果蝇中,发现了Sirt1、Sirt2、Sirt4和Sirt6基因对黑腹果蝇的代谢平衡、寿命具有调控作用[26-30],而其他昆虫Sirtuin蛋白的功能研究还相对匮乏。蜜蜂是自然界的一类重要授粉昆虫,对维持自然生态系统的稳定具有重要意义,研究发现西方蜜蜂有6种Sirtuin蛋白家族基因[7],Sir2基因在西方蜜蜂蜂王和工蜂表达存在差异,食物中氨基酸与碳水化合物比例可影响Sir2基因表达,显示Sir2可影响西方蜜蜂寿命[31-32]。鉴于国内外有关西方蜜蜂Sirtuin蛋白家族基因的系统研究还鲜有报道,本研究采用生物信息学方法对西方蜜蜂Sirtuin蛋白家族基因进行鉴定,分析该家族基因的结构、染色体位置、结构域和系统发育关系,以及在西方蜜蜂不同发育时期的表达模式,以期为后续探讨西方蜜蜂Sirtuin蛋白家族基因的功能提供基础数据。
1 材料与方法
1.1 材料
1.1.1 数据 西方蜜蜂全基因组来自2006年Weinstock等[33]公布的数据,并选取7种昆虫(东方蜜蜂A.cerana、大蜜蜂A.dorsata、小蜜蜂A.florea、地熊蜂Bombusterrestris、苜蓿切叶蜂Megachilerotundata、家蚕Bombyxmori、黑脉金斑蝶Danausplexippus)与西方蜜蜂进行Sirtuin蛋白家族基因的系统发育分析,7种昆虫的基因组数据来自NCBI数据库(https://www.ncbi.nlm.nih.gov/genome/)。
1.1.2 供试昆虫 西方蜜蜂采自云南省农业科学院蚕桑蜜蜂研究所国家现代农业产业技术体系(蜜蜂)红河综合试验站西方蜜蜂试验蜂场。选择3群群势相当的健康蜂群,放入一张空巢脾,待蜂王产卵后,分别取2、3、5日龄工蜂幼虫、2日龄工蜂蛹(复眼为粉色)和羽化出房工蜂。移1日龄幼虫到装有王浆的育王框内,放入原蜂群的继箱里育王,分别取2、3、5日龄蜂王幼虫、2日龄蜂王蛹(复眼为棕色)和羽化出房处女蜂王。相同虫态的蜂王和工蜂样本所含个体数一致,即2日龄幼虫以10头为一个样本;3日龄幼虫以6头为一个样本;5日龄幼虫和2日龄蛹均以3头为一个样本;羽化成蜂以3只成蜂头部为一个样本,每个样本3个生物学重复。取样后采用液氮冻毙,-80 ℃保存。
1.1.3 试剂 Trizol试剂购自Invitrogen公司;RNA反转录试剂盒、TB Green Premix ExTaqⅡ购自宝日医生物技术(北京)有限公司;DEPC水购自北京索莱宝科技有限公司。引物合成由生工生物合成(上海)股份有限公司完成。
1.2 西方蜜蜂Sirtuin蛋白家族基因的鉴定及分析
1.2.1 Sirtuin蛋白家族基因鉴定 为鉴定西方蜜蜂Sirtuin蛋白家族基因,在NCBI数据库中下载相关序列信息,将序列提交至Pfam数据库(https://pfam.xfam.org/),获得保守结构域隐马尔可夫模型序列文件(PF02146)。采用HMMER 3.0软件将模型序列文件转化为hmm格式,使用hmmsearch程序在氨基酸序列数据中进行查找,设置阈值为e<10-5,找到相应候选Sirtuin家族基因,使用序列抽提软件Seqtk抽提相应的氨基酸序列FASTA格式文件。使用在线数据库SMART(https://smart.embl-heidelberg.de/)对氨基酸序列进行预测,评估其是否包含Sirtuin结构域。
1.2.2 西方蜜蜂Sirtuin蛋白家族基因分子特征及定位 采用在线软件ProtParam tool(https://web.expasy.org/protparam/)对西方蜜蜂所鉴定的Sirtuin蛋白家族基因进行氨基酸数目计算,预测各家族基因的蛋白质分子量、等电点。使用在线软件WoLF PSORT(https://www.genscript.com/wolf-psort.html)对各家族基因的亚细胞位置进行预测。使用本地BLAST比对程序将预测的西方蜜蜂Sirtuin蛋白家族基因氨基酸序列与CDS序列数据进行比对,确定相应基因CDS序列。将CDS序列与参考基因组数据进行比对,明确Sirtuin蛋白家族基因在参考基因组的位置。采用BioEdit对参考基因组上各个染色体长度进行统计,使用R语言包RIdeogram绘制Sirtuin蛋白家族基因在参考基因组上的物理位置。基于BLASTN比对结果,获得各家族基因的外显子数量及长度信息,使用R语言包gggenes绘制家族基因结构图。
1.2.3 西方蜜蜂Sirtuin蛋白家族基因系统发育及保守结构域分析 将西方蜜蜂Sirtuin蛋白家族基因,以及7种昆虫的Sirtuin家族基因(基因鉴定方法同“1.2.1”)氨基酸序列合并,使用MEGA 7.0软件对Sirtuin蛋白氨基酸序列进行多重序列比对。基于最大似然法(maximum likelihood,ML)构建系统发育树,设置bootstrap值为 1 000。使用Clustal W对西方蜜蜂Sirtuin蛋白家族基因进行多重序列比对,将比对结果导入至GeneDoc中,预测其保守结构域。在Pfam数据库中查找人类、动物、植物等6 942个物种19 213条序列所构建的Sirtuin保守结构域信息(HMM logo),与西方蜜蜂预测的保守结构域进行比对分析。
1.3 西方蜜蜂Sirtuin蛋白家族基因qRT-PCR分析
采用Trizol法提取西方蜜蜂总RNA,取 1 μL RNA测定其浓度及OD值(A260/A280),要求OD值为1.8~2.1,利用反转录试剂盒将RNA反转录成cDNA,获得的cDNA在-20 ℃保存,备用。以Zhang等[34]报道的Gapdh为内参基因,采用实时荧光定量仪(StepOnePlusTM型,Applied Biosystems公司)检测西方蜜蜂6个Sirtuin蛋白家族基因(AmSirt1、AmSirt2、AmSirt4~AmSirt7)相对表达量,相关基因引物序列信息见表1。反应体系按说明书进行操作,反应程序:95 ℃,10 min;95 ℃,15 s,57 ℃,1 min(40个循环);添加熔解曲线;设置3个水平重复孔;以2-△△Ct法计算基因的相对表达量。采用SPSS 17.0进行单因素方差分析和多重比较。
有一天,老大爷阿扁在厅堂下做车轮,齐桓公正好在厅堂上看书。老大爷阿扁就想找齐桓公聊聊,于是放下干活的工具,走到厅堂上。阿扁说:“冒昧地请教一下,您读的是什么?”齐桓公答:“圣贤说的话啊。怎么了?”阿扁又问:“那圣人还活着吗?”齐桓公答:“已经死了。”阿扁摇摇头,说:“唉,您读的书不过是古人的糟粕而已!”齐桓公差点被这句话给气炸了,心想:“本人堂堂一国之君,亲切地接见一个小小的老工匠,他不但不识抬举,还不给面子。”于是,愤怒地说:“你一个做车轮的工匠,有什么资格评议寡人读什么书,你懂什么?你说不出个所以然来,寡人就判你死刑,把你给砍了!”

表1 实时荧光定量PCR扩增引物序列信息Table 1 Primer information of real-time quantitative PCR
2 结果与分析
2.1 西方蜜蜂Sirtuin蛋白家族成员鉴定与分布
在西方蜜蜂基因组中共鉴定出10个Sirtuin蛋白基因,属于Sirtuin蛋白的6个家族成员,按直系同源物分别命名为AmSirt1(AmSirt1-1~AmSirt1-3)、AmSirt2(AmSirt2-1~AmSirt2-3)、AmSirt4~AmSirt7(表2)。6个家族基因分别定位于不同的染色体,其中AmSirt1和AmSirt2呈现典型串联重复分布,其余基因则单独存在。AmSirt1-1~AmSirt1-3分布于4号染色体;AmSirt2-1~AmSirt2-3分布于10号染色体;AmSirt4分布于7号染色体;AmSirt5分布于14号染色体,AmSirt6分布于3号染色体,AmSirt7分布于11号染色体(图1)。对西方蜜蜂Sirtuin蛋白家族基因分子特征及亚细胞位置预测分析显示,其编码氨基酸数为265~899个,分子质量为29.45~102.91 ku,等电点为4.58~ 9.09。AmSirt1-1、AmSirt6和AmSirt7定位于胞质/细胞核,AmSirt1-2、AmSirt1-3、AmSirt2-2、AmSirt2-3、AmSirt5定位于胞质,AmSirt2-1和AmSirt4定位于线粒体(表2)。
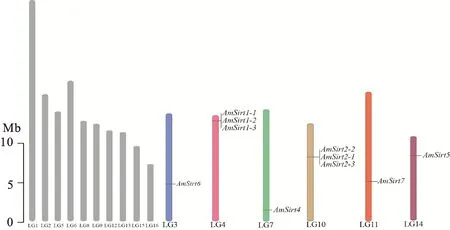
灰色柱子:不含Sirtuin蛋白家族基因的染色体;有色柱子:含有Sirtuin蛋白家族基因的染色体
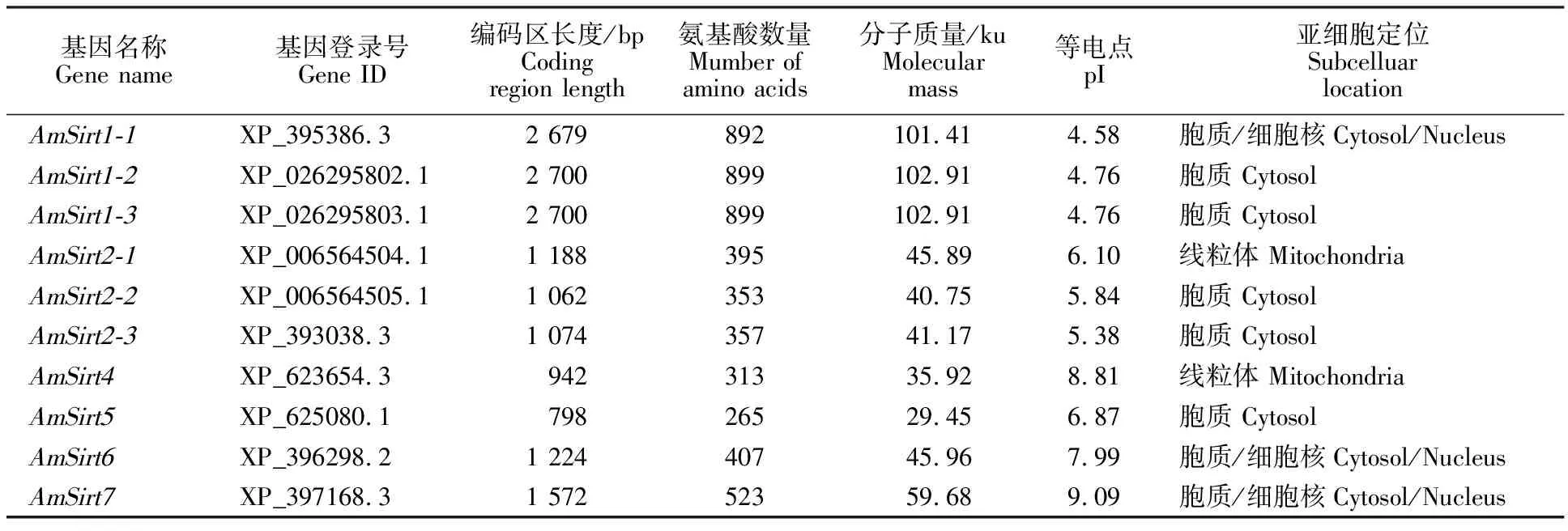
表2 西方蜜蜂Sirtuin蛋白家族基因鉴定及序列特征Table 2 Identification and sequence characterization of Sirtuin protein family genes in Apis mellifera
2.2 西方蜜蜂Sirtuin蛋白家族基因结构分析
西方蜜蜂Sirtuin蛋白家族基因平均外显子数为6.5个,其中AmSirt1-1~AmSirt1-3外显子数最多(9个),AmSirt4和AmSirt7外显子数最少(4个)。4个基因(AmSirt7、AmSirt1-1~AmSirt1-3)具有较长的外显子(844~1 159 bp), 2个基因(AmSirt2-2和AmSirt2-3)第1外显子较短(17 bp和28 bp)。3个基因(AmSirt1-1~AmSirt1-3)具有最长的内含子(均为2 719 bp), 2个基因(AmSirt5、AmSirt7)内含子较短(56~62 bp)。在AmSirt1-1~AmSirt1-3中,除AmSirt1-1第1内含子与其他2个基因有差异外,其余外显子和内含子长度基本一致。在AmSirt2-1~AmSirt2-3中,除第1外显子和内含子有差异外,其余外显子和内含子长度基本一致(图2)。说明Sirtuin蛋白同一家族内不同基因的中、后端外显子和内含子具有一定的保守性。

图2 西方蜜蜂Sirtuin蛋白家族基因结构Fig.2 Structure of Sirtuin protein family genes in Apis mellifera
2.3 西方蜜蜂等8种昆虫的Sirtuin蛋白家族基因系统发育分析
对西方蜜蜂等8种昆虫Sirtuin蛋白家族基因分析(表3)显示,8种昆虫均未发现Sirt3家族基因。西方蜜蜂、东方蜜蜂、小蜜蜂、大蜜蜂和地熊蜂5种蜜蜂属昆虫均含有(Sirt1、Sirt2、Sirt4~Sirt7)6个家族基因,但种间基因数量存在差异,西方蜜蜂和大蜜蜂基因数最多(10个),分布基本一致,小蜜蜂基因数最少(6个)。苜蓿切叶蜂家族基因成员与蜜蜂属5个种类一致,基因数达到14个;家蚕和黑脉金斑蝶家族基因成员及基因数均为5个,缺少Sirt1家族基因。基于ML法构建西方蜜蜂等8种昆虫Sirtuin蛋白家族基因的系统发育树(图3),Sirtuin蛋白家族基因明显聚合为4类6个分支(Sirt1、Sirt2、Sirt4~Sirt7),其中Sirt1和Sirt2聚为第Ⅰ类(bootstrap值100%),Sirt4和Sirt5由于bootstrap值较低,分属第Ⅱ、Ⅲ类,Sirt6和Sirt7聚为第Ⅳ类(bootstrap值100%)。西方蜜蜂3个AmSirt2基因(AmSirt2-1~AmSirt2-3)聚为一个小支(bootstrap值99%),并与东方蜜蜂的2条基因聚为一支(bootstrap值90%)。3个AmSirt1基因(AmSirt1-1~AmSirt1-3)分化为两个小支,两个小支分别与大蜜蜂3条基因聚合(bootstrap值分别为84%和90%)。西方蜜蜂AmSirt4、AmSirt5分别与东方蜜蜂、大蜜蜂和小蜜蜂的Sirt4、Sirt5基因聚为一支(bootstrap均为值98%),AmSirt7与东方蜜蜂、小蜜蜂和大蜜蜂的3条基因聚为一支(bootstrap值74%),AmSirt6与蜜蜂属近缘物种未形成明显聚类分支。
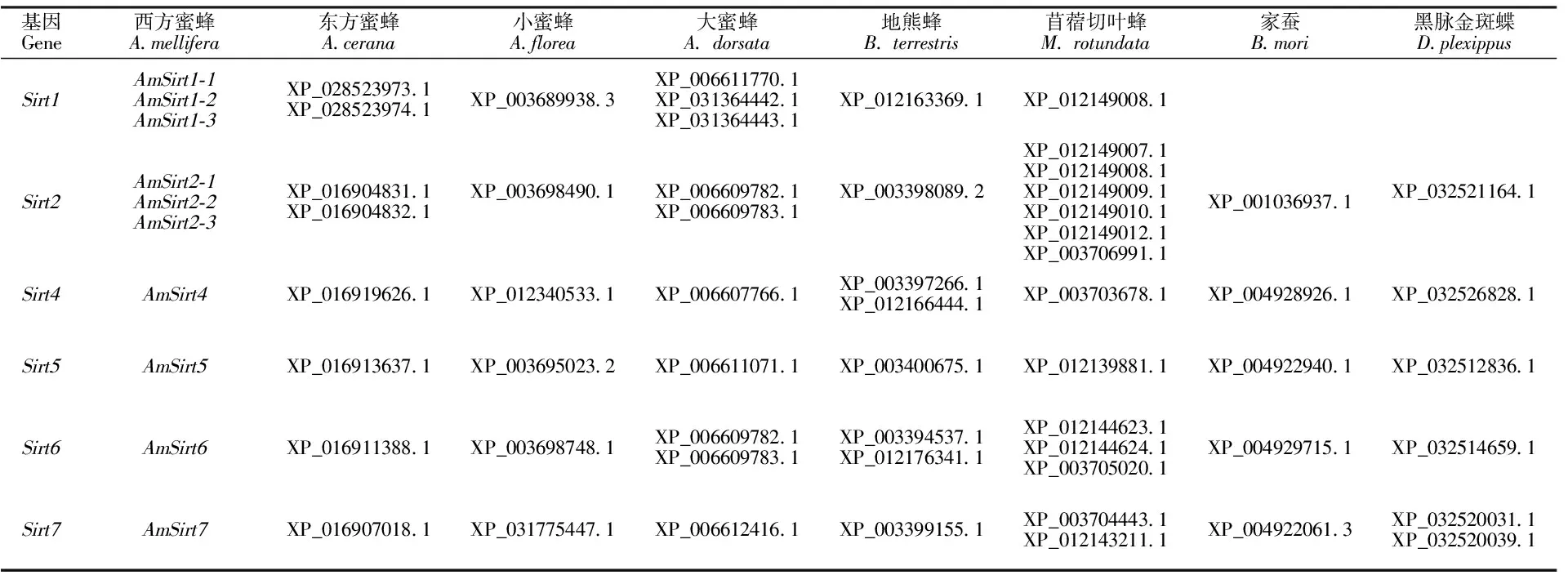
表3 西方蜜蜂等8种昆虫的Sirtuin蛋白家族基因分析Table 3 Genes analysis of Sirtuin protein family in eight kinds of insects contanining Apis mellifera
2.4 西方蜜蜂Sirtuin蛋白家族基因保守结构域分析
对西方蜜蜂6个Sirtuin蛋白家族基因氨基酸序列的保守结构域分析结果(图4)显示,与Pfam数据库中多个物种的Sirtuin蛋白保守结构域相比,西方蜜蜂家族基因具有Sirtuin蛋白的典型特征,存在12个保守结构域,其中在6个家族基因氨基酸序列前段均存在一段高度保守的富含甘氨酸的结构域(结构域1~3),在中段均存在QN-D-L-G的保守结构域(结构域6~7)。此外,发现3个西方蜜蜂所特有的保守氨基酸位点,即在结构域4~5之间存在1个高度保守的脯氨酸位点,在结构域7~8之间存在1个高度保守的谷氨酸位点,在基因后段存在1个高度保守的天冬酰胺位点。
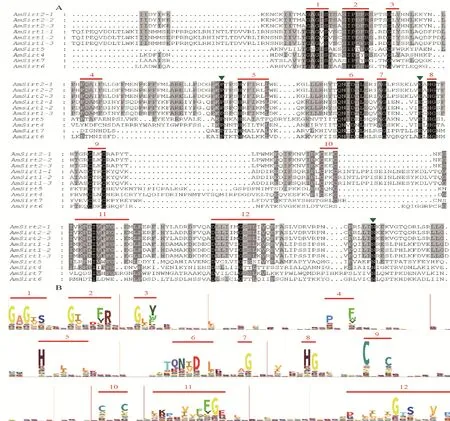
A.西方蜜蜂Sirtuin蛋白氨基酸序列比对结果;B.Pfam数据库相关Sirtuin蛋白氨基酸序列比对结果;数字1~12代表保守结构域,绿色倒三角代表西方蜜蜂所特有的保守氨基酸位点
2.5 西方蜜蜂Sirtuin蛋白家族基因表达模式分析
Sirtuin蛋白家族基因在西方蜜蜂蜂王和工蜂不同发育时期均有表达。在蜂王中,6个基因在幼虫2日龄到5日龄的表达量总体呈逐渐下降,至蛹期逐渐上升趋势。以3日龄和5日龄幼虫的基因表达量相对较低,5日龄幼虫基因表达量显著低于2日龄蛹(P<0.05);成虫期除AmSirt1与AmSirt6表达量较低外,其余4个基因表达量与2日龄蛹相当(图5)。在工蜂中,6个基因在2日龄、3日龄和5日龄幼虫的表达量相对较低,均在2日龄蛹期达到峰值;2日龄蛹期与3个日龄幼虫期均达到显著差异(P<0.05),2日龄蛹期到成虫期呈下降趋势(图6)。

L2.2日龄幼虫;L3.3日龄幼虫;L5.5日龄幼虫;P2.2日龄蛹;EB.羽化蜂;不同字母表示不同发育时期存在差异显著(P<0.05,Duncan’s test),下同
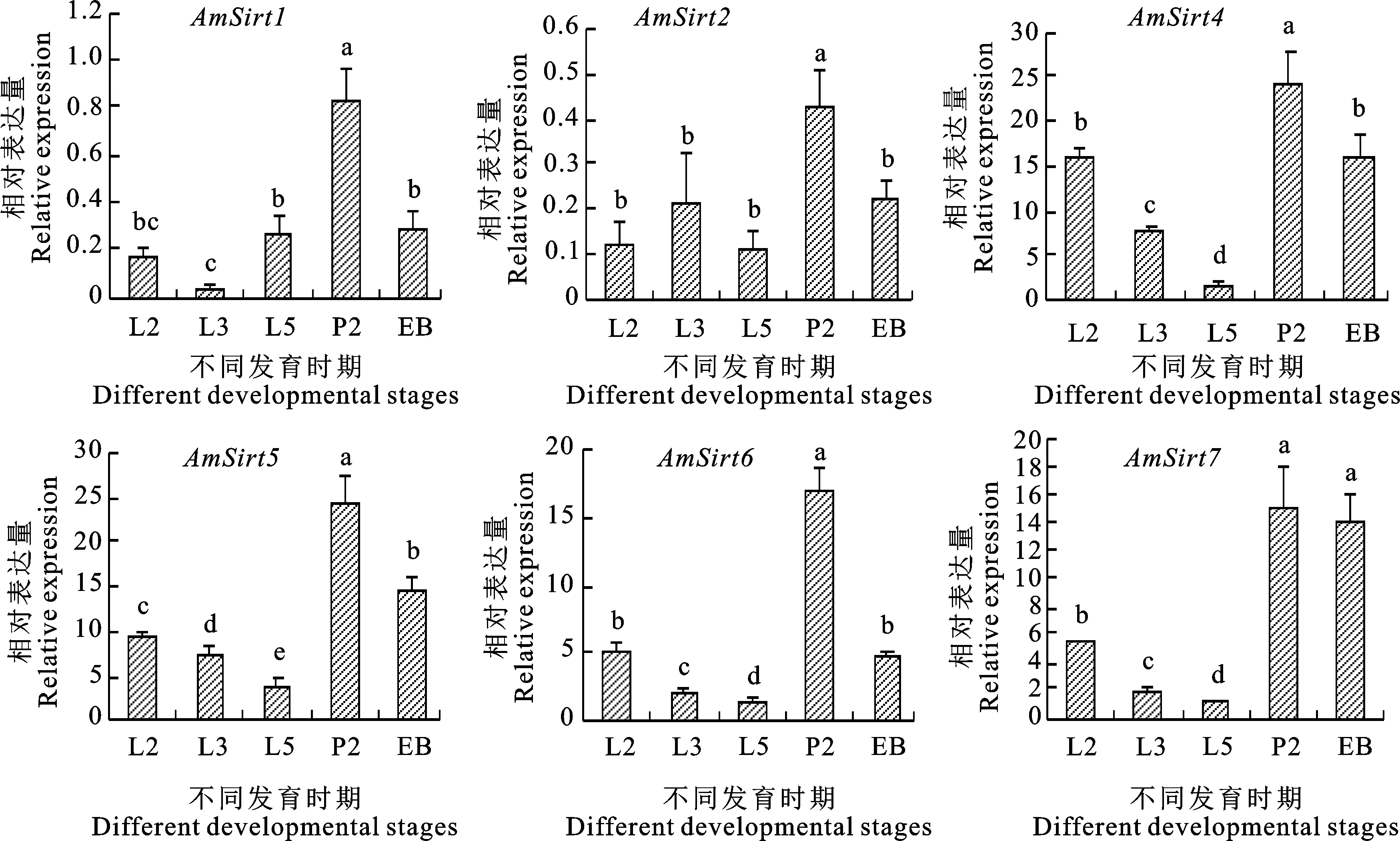
图6 Sirtuin蛋白家族基因在西方蜜蜂工蜂不同发育时期的相对表达量Fig.6 Relative expression of Sirtuin protein family genes at different developmental stages of Apis mellifera worker
3 讨 论
Sirtuin蛋白是一类进化保守的NAD+依赖性脱酰基酶家族,从原核生物到哺乳动物的几乎所有物种中都存在1种或多种Sirtuin蛋白[6-7]。本研究发现,西方蜜蜂等8种昆虫同时缺少Sirt3家族成员,膜翅目6种昆虫均有6个家族成员,而鳞翅目2种昆虫仅有5个。这与Frye[6]和Greiss等[7]在不同生物的Sirtuin基因系统进化分析结果一致。值得注意的是,在真核生物中类似Sirtuin蛋白家族成员基因的现象较为普遍。Greiss等[7]研究表明,在蜕皮动物总门(昆虫纲、线虫纲)的物种中均缺失Sirt3基因,推断Sirt3在蜕皮动物进化早期丢失。Frye[6]依据真核生物起源于细菌与古生菌融合假说,提出第一个真核生物从原核亲本获得3类Sirtuin家族基因(Ⅱ、Ⅲ和U),早期真核生物在进化中出现4类家族基因(Ⅰ~Ⅳ),在真核生物的进化中,部分真核生物失去1类(缺失Ⅲ类的西方蜜蜂和赤拟谷盗[7])、失去2类(缺失Ⅰ、Ⅲ类的家蚕和黑脉金斑蝶[4]),或失去3类(缺失Ⅱ、Ⅲ和Ⅳ类的酿酒酵母[6])。关于昆虫丢失Sirt3基因的现象,一方面可能与昆虫物种进化有关,如膜翅目和鞘翅目昆虫家族成员基因数量偏多,鳞翅目和双翅目昆虫数量偏少[4,7];
另一方面暗示昆虫体内保留的Sirtuin蛋白基因可能赋予冗余功能以补偿基因的缺失,如Sirt4是黑腹果蝇中唯一定位于线粒体的家族基因[29],黑腹果蝇Sirt2与哺乳动物Sirt3功能相似,可调节ATP的合成[28]。本研究也发现西方蜜蜂Sirt4基因定位于线粒体,预示西方蜜蜂Sirt2和Sirt4基因可能赋予冗余功能,以弥补Sirt3基因的 缺失。
为探讨Sirtuin蛋白家族基因对西方蜜蜂生长发育的调控作用,本研究对西方蜜蜂雌性蜂不同发育时期的6个家族基因进行qRT-PCR分析。结果显示,6个家族基因在蜂王和工蜂不同发育时期均有表达,蜂王各家族基因表达量整体高于工蜂,蜂王和工蜂各家族基因表达量均在2日龄蛹期达到峰值。近年以黑腹果蝇为代表的昆虫Sirtuin蛋白家族基因的功能得到系统研究[26-30],黑腹果蝇Sirt1在脂肪体中起到维持胰岛素信号传导和代谢稳态的作用[26],并通过调节Notch通路影响黑腹果蝇感觉器官前体和翅的发育过程[27]。Sirt2是线粒体能量代谢的重要调节器,通过ATP合酶β调节ATP合成[28],Sirt4可介导黑腹果蝇对禁食反应、代谢稳态作用,禁食可诱导脂肪体中Sirt4基因上调[29]。Sirt6基因过表达可延长黑腹果蝇寿命,引起抗氧化应激能力增强,蛋白质合成率降低[30]。蛹期是全变态昆虫所特有的时期,尽管在蛹期处于不食不动的禁食状态,但体内进行着组织器官(触角、胸足、翅等)的分化[35]。因此推测蛹期的禁食行为、热量限制和强烈的器官分化是引起西方蜜蜂雌性蜂6个Sirtuin家族基因表达量明显上升的主要原因。
此外,6个家族基因中以Sirt4和Sirt5表达量较高,Sirt1和Sirt2的表达量较低,推测与基因功能相关。如Sirt4、Sirt5主要涉及线粒体能量代谢过程,生物体禁食和热量限制均可诱导Sirt4和Sirt5基因上调[21,29],黑腹果蝇禁食可引起葡萄糖-1-磷酸和葡萄糖-6-磷酸水平显著降低[29],意大利蜜蜂体内葡萄糖-6-磷酸异构酶在蛹期的表达量较低[36],说明禁食行为对昆虫糖酵解中间产物有显著影响。而Sirt1涉及生物体细胞凋亡、组织分化、能量代谢和寿命调节等多种功能[15-16],Sirt2控制细胞周期进程和基因组稳定性[17],Sirt1和Sirt2对生物调节功能的多样性也预示着自身表达水平可能受多种因素的调控。本研究也发现,西方蜜蜂AmSirt1和AmSirt2出现典型的串联重复现象,暗示西方蜜蜂AmSirt1和AmSirt2功能的复杂性。目前有关昆虫Sirtuin蛋白家族基因的研究还相对匮乏,后续还需要进一步探讨和验证西方蜜蜂Sirtuin蛋白家族成员的功能,以期为揭示Sirtuin蛋白对昆虫生长发育、新陈代谢与寿命的调控机制提供参考依据。
4 结 论
本研究从西方蜜蜂全基因组数据中鉴定出10个Sirtuin蛋白基因,分属6个家族成员基因(AmSirt1、AmSirt2、AmSirt4~AmSirt7),AmSirt1和AmSirt2家族成员各包含3个基因,缺少Sirt3基因。利用生物信息学分析西方蜜蜂Sirtuin蛋白家族基因的结构、染色体位置和保守结构域,通过qRT-PCR进一步明确Sirtuin蛋白家族基因在西方蜜蜂雌性蜂不同发育时期的表达情况,研究结果为后续系统研究西方蜜蜂Sirtuin蛋白家族基因的结构和功能奠定基础。
