FaesCAL基因在同型长花柱甜荞中的表达分析
2023-10-31马志远王子翔刘志雄
马志远,王子翔,刘志雄
(长江大学 园艺园林学院,湖北荆州 434025)
甜荞(FagopyrumesculentumMoench.)是蓼科(Polygonaceae)荞麦属(Fagopyrum)兼食、药、观赏和土壤修复等多种用途的经济作物,荞麦籽粒不含谷蛋白,但富含类黄酮、多酚、多糖、赖氨酸和抗氧化活性物等,近年作为一种具疾病预防和保健功效的功能食品备受关注[1-3]。然而,甜荞因二型花柱导致的异型自交不亲和,自然群体中短花柱长雄蕊(thrum)和长花柱短雄蕊(pin)植株1∶1分离,仅异型花间互相授粉才能正常结实,同型花自交不亲和,产量低,杂交育种因难,成为制约这一重要经济作物推广应用的瓶颈[4]。寻找或创制亲和性好的甜荞种质资源,对于开展甜荞杂交育种工作、提高甜荞产量均具有重要的意义。笔者课题组前期在甜荞品种‘北早生’群体中发现长花柱长雄蕊lpls自然突变体单株基础上,通过隔离传粉获得了遗传性状稳定、亲和性好、自花、与同型、异型花间均能正常授粉结实的同型长花柱甜荞株系(图1),成为甜荞高产育种不可或缺的种质资源。深入研究同型长花柱甜荞花和果实的发育规律与分子调控机制,是合理利用该材料的前提和基础。
在拟南芥(ArabidopsisthalianaL.)中,CAULIFLOWER(CAL) 基因编码一个MADS-box转录因子,其在花发育过程中参与成花转变、花分生组织特性决定和花原基的形成[5-6]。为探究CAL同源基因在甜荞花和果实发育调控中的功能是否保守,本研究在分离甜荞CAL同源基因FaesCAL基础上,结合qRT-PCR和石蜡切片检测该基因在同型长花柱甜荞中表达的组织特异性和花芽分化关键时期表达量的动态变化,从而解析该基因在与参与同型长花柱甜荞花和果实发育中的作用和功能,以期为甜荞分子改良和杂交育种积累资料和资源。
1 材料与方法
1.1 试验材料
挑选同型长花柱甜荞籽粒饱满的种子,种植在盛满营养基质(主要成分为泥炭土和椰糠)的规格为21 cm×14 cm×20 cm的花盆中,将花盆露天放置在种质资源圃内,常规水肥管理,在盛花期及坐果时搬至实验室内,用经过消毒灭菌的镊子和剪刀分别从不同植株(≥3)上采集根、茎、叶片、雄蕊、雌蕊、花被片以及发育5 d的果实,置于 2 mL离心管中,经液氮速冻后存放于-80 ℃冰箱中备用。在甜荞花序发育和花芽分化时期,分别将不同发育时期的花芽用小剪刀剪下,每个时期的花芽分成2份,一份经液氮速冻后存放于 -80 ℃冰箱中备用,另一份固定在FAA[V(甲醛)∶V(冰乙酸)∶V(70%乙醇)=1∶1 ∶18]中,经石蜡切片确定花芽的发育时期。
1.2 试验方法
1.2.1 同型长花柱甜荞CAL同源基因的克隆 根据笔者课题组前期甜荞花芽的第3代转录组测序结果(BioProject ID:PRJNA517031)拼接获得甜荞CAL同源基因序列,根据3′- Full Race Core Set Ver.2.0 试剂盒(TaKaRa)说明书上的引物设计原则,采用Oligo 7.0软件在5′非翻译区设计基因特异性引物GSPFaesCAL,以同型长花柱甜荞的花芽总RNA 为模板,采用 3′- Full Race Core Set Ver.2.0 试剂盒(TaKaRa)反转录试剂盒合成甜荞花芽3′RACE第1链cDNA,PCR扩增获得同型长花柱甜荞FaesCAL同源序列的全长序列。PCR扩增程序为94 ℃预变性 5 min, 94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸 1 min; 共30个循环,最后72 ℃ 延伸 10 min。扩增产物经检测后送至上海生工生物公司测序,克隆获得甜荞FaesCAL同源基因的全长序列。试验所用引物见表1。
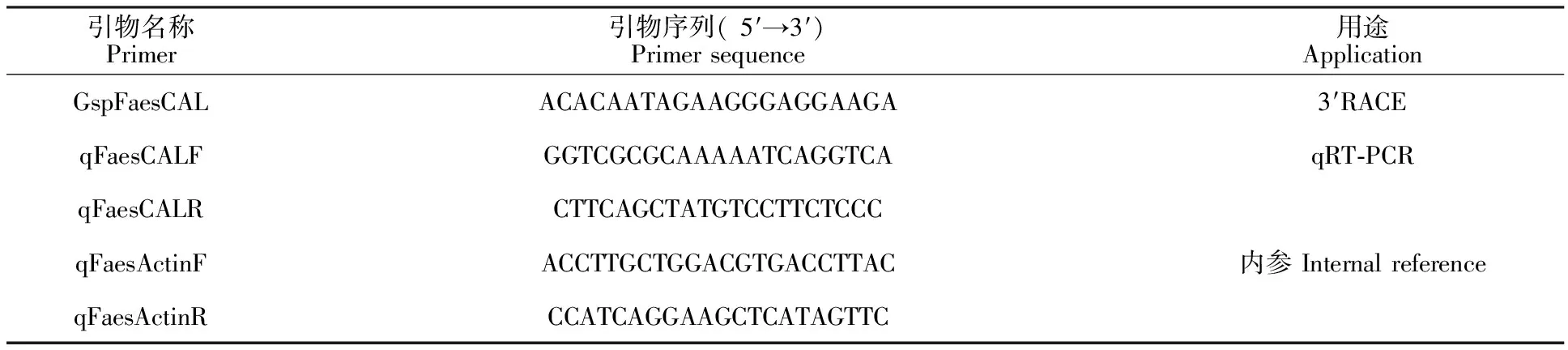
表1 引物序列及用途Table 1 Primer sequences and application
1.2.2 蛋白同源序列比对及分子系统发育分析 将同型长花柱甜荞FaesCAL同源基因的开放阅读框(open reading frame,ORF)编码的氨基酸序列在 NCBI 数 据 库(https://blast.Ncbi.Nlm.Nih.gov/Blast.Cgi)中进行 BlastP 同源比对搜索,选取麻风树(JatrophacurcasL.)、蓖麻(RicinuscommunisL.)和棉花(GossypiumhirsutumL.)等植物的CAL同源蛋白,采用 MEGA 7.0 软件的邻接法(Neighbor-Joining,NJ) 构建蛋白序列的分子系统发育树。同时选取5个物种的同源蛋白,采用Bioedit7.0软件中的ClustalW程序,对FaesCAL转录因子的结构域进行分析。

按照王旋等[7]的方法制作同型长花柱甜荞不同时期花芽的石蜡切片:将在FAA中固定好的花芽经脱水、透明、浸蜡、组织包埋之后,利用Leica RM2235石蜡切片机将包埋入石蜡的材料切成厚度为8 μm的连续切片。切片使用1%番 红-0.1%固绿染色后封片保存。制作的甜荞花芽切片在Caikon RCK-40C显微镜下观察细胞的形态结构并拍照,通过细胞形态特征确定花芽发育的时期。并选取5个关键发育时期的花芽,分别提取其总RNA,通过qRT-PCR检测FaesCAL基因在甜荞花芽分化5个关键时期表达量的动态变化。
2 结果与分析
2.1 甜荞FaesCAL基因全长cDNA克隆
通过3′RACE技术从同型长花柱甜荞中分离到CAL基因的cDNA全长,CAL的同源基因cDNA全长为984 bp,包含1个28 bp的3′UTR、726 bp的完整ORF、230 bp的5′UTR,编码241个氨基酸残基和1个终止密码子。结构分析表明该基因为MADS-box基因家族的CAL进化系,命名为FaesCAL(Genbank登录号:OM056 742)。
2.2 蛋白质分析比对与分子系统发育分析
蛋白质同源比对分析结果显示(图2):FaesCAL转录因子的N端含有一个高度保守、由57个氨基酸(1-57)残基组成的MADS-box结构域,该结构域通过与DNA结合, 调控下游基因的表达[8];该转录因子还含有一个由68个氨基酸残基(94-161)组成的次级保守的K结构域,其C末端转录激活域在序列长度和氨基酸组成与其他同源蛋白差异较大。分子系统发育与进化树重建显示(图3):甜荞FaesCAL转录因子与其他双子叶植物CAL-like转录因子聚于1个进化分支,其与藜科植物藜麦(Chenopodiumquinoa)的CqCAL亲缘关系最近,共同聚到一个小的进化分支,经典分类学中同源蛋白所属植物种属间的进化关系也在树上得以很好地呈现。
2.3 FaesCAL基因在同型长花柱甜荞中的表达模式分析
FaesCAL基因的表达组织特异性分析表明(图4-A):FaesCAL基因主要在同型长花柱甜荞根、花被片、雄蕊以及发育5 d的果实中表达,在雌蕊中仅能检测到微弱的转录信号,在茎和叶片中不表达。从其表达量的差异来看,其在雄蕊中的表达量最高,极显著高于其在其他组织的表达量(LSD,P<0.01);在花被片中的表达量其次,但极显著高于其在根、雌蕊以及发育5 d果实中的表达量(LSD,P<0.01)。
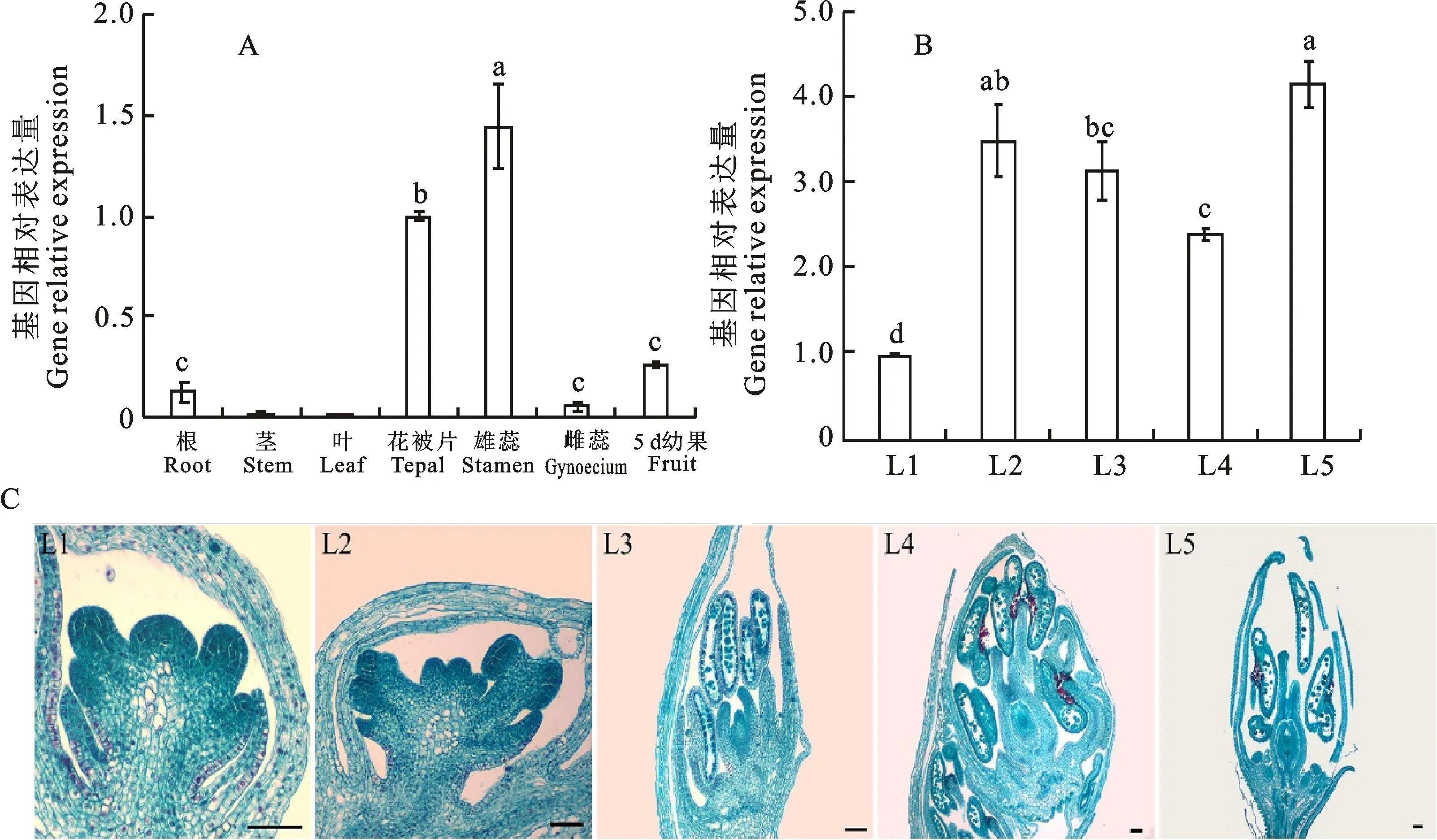
A. FaesCAL基因在同型长花柱甜荞不同组织中的表达量;图中小写字母表示不同组织之间的差异显著性;B. FaesCAL基因在同型长花柱甜荞花芽分化5个关键时期的表达量;L1.雄蕊原基出现;L2.雄蕊花丝快速伸长、花被片原基出现;L3.小孢子母细胞减数分裂、小孢子四分体的形成;L4.单核小孢子靠边期、花被片闭合;L5.开花前雌雄蕊均发育成熟的花芽;图柱上不同小写字母表示花芽不同发育时期之间的差异显著性;C. 同型长花柱甜荞花芽分化5个关键时期的石蜡切片图;比例尺:100 μm
进一步分析该基因在同型长花柱甜荞花芽分化5个关键时期表达量的动态变化发现(图4-B、4-C):随着同型长花柱甜荞雄蕊原基的出现(L1),FaesCAL基因在花芽中有明显的表达(图4B-L1、C-L1);随着雄蕊花丝的迅速伸长和花被片原基的出现(L2),FaesCAL基因的表达水平迅速升高,直到小孢子母细胞减数分裂和小孢子四分体的形成时(L3)都维持在较高水平;随着花被片的闭合和花药中出现单核靠边期小孢子(L4),FaesCAL基因的表达呈现缓慢的下降;在开花前,随着花芽雌、雄蕊发育成熟(L5),基因的表达量迅速升高并达到峰值。从基因表达量的差异显著性来看:花芽成熟时(L5)FaesCAL基因的相对表达量最高,与L2时期该基因的相对表达量无显著差异,但显著高于其在L1、L3和L4共3个时期的相对表达量(LSD,P<0.05);在花丝快速伸长和花被片原基出现时FaesCAL基因的表达量其次,其与L3时期该基因的相对表达量无显著差异,但显著高于其在L1和L4时期的相对表达量(LSD,P<0.05)。
3 讨 论
在拟南芥中,CAL开始在花分生组织中表达,随着花萼的发育,花瓣和雄蕊原基的出现,其转录活性主要集中在花萼和花瓣中,同时其在花序的维管束中也有明显表达,在花发育过程中主要参与花分生组织特征决定和花原基的形成,调控AP1基因参与花萼和花瓣的发育,并促进开花[9-11]。拟南芥近缘种甘蓝(Brassicaoleraceavar.capitataL.) 的CAL-like基因BoCAL在甘蓝花球的形成及发育中发挥重要作用,在拟南芥中的过表达能促使其提早开花[12-13]。花椰菜(Brassicaoleraceavar.botrytisL.)CAL同源基因BobCAL主要在花序分生组织和花原基中表达,之后随着花芽分化之后,表达量显著下降,主要参与花椰菜花分生组织的的形成,并具有促进拟南芥提早开花的功能[13-14]。在蓝莓(VacciniumcorymbosumL.)中,其CAL-like基因在花中表达量最高,主要参与促进开花[15]。棉花(GossypiumhirsutumL.)的CAL-like基因GhCAL主要在茎尖、茎、芽和叶片中表达,在根中不表达,在茎尖的表达量最高,其通过调控下游GhAP1-A04和GhAGL6-D09基因的表达,从而促进棉花从营养生长到生殖生长的转变[16]。麻风树(JatrophacurcasL.)的CAL-like基因JcCALA在雄花小孢子母细胞形成期的表达量显著升高,在随后的四分体时期又有较大幅度下降,说明其可能在雄花小孢子母细胞的形成和发育过程中发挥作用[17]。以上的研究结果表明,双子叶植物中CAL-like 基因的表达模式与其功能有明显的相关性。
本研究通过分析FaesCAL基因在甜荞中的组织表达特异性发现:FaesCAL基因在同型长花柱甜荞雄蕊中的表达量最高,在花被片中的表达量其次。分析甜荞花芽分化5个关键时期表达量的动态变化发现:FaesCAL基因在开花前花芽雌雄蕊发育成熟时的表达量最高,其次是在雄蕊花丝的迅速伸长和花被片原基的出现时期。推测FaesCAL基因参与了甜荞的成花转变,并在雄蕊以及花被片的发育中发挥作用,但该基因在甜荞花发育中的具体作用还需要进一步研究和考证。
