四倍体小麦GPAT基因家族比较分析
2023-10-30师毅君王康君郭明明张广旭谭一罗李晓峰何茂盛徐大勇樊继伟
师毅君,王康君,郭明明,张广旭,谭一罗,李晓峰,何茂盛,徐大勇,樊继伟
(连云港市农业科学院,江苏 连云港 222000)
甘油-3-磷酸酰基转移酶(glycerol-3-phosph-ateacyltransferase, GPAT)是三酰甘油(Triacylglycerol, TAG)生物合成的关键酶,催化并限制其第一步反应的速度,是细胞膜、表皮蜡质、种内油脂等重要组分,对植物生长发育及抗逆能力具有重要影响[1]。具体而言,GPATs分为两类:一类为sn-1型GPAT,负责将脂肪酰基从酰基-酰基载体蛋白(-acyl-ACP)或酰基辅酶A(-acyl-CoA)向sn-1转移;另一类属于sn-2型GPAT,具有磷酸酶活性[2-4]。目前,已在多种植物中鉴定到GPAT基因家族成员,如拟南芥[5]、棉花[6]、玉米[7]、青稞[8]、大麦[9-10]等,其中,拟南芥GPAT家族包含10个成员[5],ATS1和AtGPAT9 属于sn-1型GPAT,AtGPAT1~8 为陆生植物特有的sn-2型GPAT,这些酶参与磷脂酰甘油(phosphatidylglycerol, PG)[11-12]、植物膜脂、植物油脂、角质和软木脂[4,13]等多种脂质的生物合成[14]。研究发现,GPAT在植物响应冷、热等胁迫中起着关键作用[15],例如,在不同抗冷性水稻中,GPAT核苷酸和推导氨基酸序列差异与抗冷性差异正相关[16]。番茄LeGPAT基因表达受低温诱导,过表达可以提高甘油-3-磷酸酰基转移酶含量而增强植株的耐冷性,抑制表达则可以增强耐热性并影响番茄的育性[17]。百合LpGPAT基因也被证明与低温胁迫有关[18]。在水稻品种J9516的叶绿体类囊体膜磷脂分子中,不饱和PG含量较高的植株能抵御低温诱导的光抑制作用[16]。反之,GPAT酶通过提高质体中类囊体膜脂的饱和度来增加植物对高温的耐受性。颜坤等[19-21]研究发现,高温胁迫下,转入GPAT基因的烟草类囊体膜上脂质饱和度增加,其光合速率较野生型的变化慢。
四倍体小麦分为硬粒小麦和野生二粒小麦,是栽培小麦的近缘物种,近年来,随着其基因组测序的完成,已逐渐成为新的研究热点,许多其他关于GPAT基因及其功能的类似研究已经被报道[14,22]。然而,人们对该基因家族在四倍体小麦中的作用知之甚少,特别是它们在冷或热胁迫中的潜在作用。为了解四倍体小麦中GPAT基因家族进化及其功能,本研究通过比较基因组学系统分析硬粒小麦和野生二粒小麦GPAT基因家族的结构特性,进化关系等内容,为深入探索麦类作物GPAT基因家族的功能提供更多的信息。
1 材料与方法
1.1 四倍体小麦GPAT基因家族成员的鉴定及理化性质分析
从植物参考基因组数据库EnsemblPlants(https://plants.ensembl.org/index.html)下载硬粒小麦、野生二粒小麦以及拟南芥的基因组数据,包括基因组序列文件、注释文件、蛋白序列文件。利用从Uniportswissport(https://www.uniprot.org/)下载拟南芥GPAT蛋白序列,在Pfam网站(http://pfam-legacy.xfam.org/)下载GPAT蛋白保守结构域(PF01553)的隐马尔可夫模型,使用TBtools(https://github.com/CJ-Chen/TBtools)软件[23]在硬粒小麦和野生二粒小麦蛋白质库进行BLASTP和HMMER搜索,得到包含结构域模型的蛋白序列。将HMMER和BLASTP的结果去冗余后,将序列提交到NCBI-CDD(https://www.ncbi.nlm.nih.gov/cdd/)数据库对其结构域进行鉴定,保留包含有完整结构域的基因。通过网站ExPASY(https://www.expasy.org/)来分析硬粒小麦和野生二粒小麦GPAT基因家族成员分子量、等电点等理化性质,并用网站WoLFPSORT(https://wolfpsort.hgc.jp/)来预测其家族成员的亚细胞定位信息。
1.2 四倍体小麦GPAT基因家族系统发育分析
使用软件MEGA 11.0软件内置muscle对硬粒小麦、野生二粒小麦、拟南芥的所有GPAT基因进行多序列对比分析,使用邻接法Neighbor-joiningTree构建GPAT基因家族系统进化树,Bootstrap值取1000。通过网站ITOL(https://itol.embl.de/)进行美化。
1.3 四倍体小麦GPAT基因家族基因结构和保守结构域分析
硬粒小麦、野生二粒小麦和拟南芥GPAT基因结构分析使用TBtools软件中Visualize Gene Structure(Basic)工具,根据其基因组注释gff3文件提取内含子和外显子信息;通过网站MEME(http://memesuite.org/)预测蛋白的保守结构域(Motif设为10);通过网站NCBI-CDD(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)确认GPAT家族成员的保守结构域。使用TBtools软件的GeneStructureView工具将上述结果进行合成及可视化。
1.4 四倍体小麦GPAT基因家族共线性分析
根据硬粒小麦、野生二粒小麦和拟南芥全基因组及注释文件,使用TBtools对其进行可视化的共线性关系分析,以此发现三者GPAT基因家族间的同源基因对。
2 结果与分析
2.1 GPAT基因家族成员的鉴定结果及理化性质分析
利用拟南芥GPAT蛋白序列,在硬粒小麦和野生二粒小麦蛋白质库进行BLASTP和HMMER搜索鉴定,最终确定硬粒小麦和野生二粒小麦各有33个GPAT基因家族成员。由表1可知,GPATs蛋白质成员的氨基酸长度为149(TdGPAT19)~589(TtGPAT1),分子量为16.22 kDa(TdGPAT19)~62.76 kDa(TtGPAT1),等电点为5.92(TdGPAT5)~11.39(TdGPAT7),但绝大多数蛋白(56个)的pI大于8,说明大多数GPATs蛋白富含碱性氨基酸,属碱性蛋白。亚细胞定位结果显示:有40.9%成员定位在质膜,25.7%成员定位在叶绿体,12.1%成员定位在内质网,10.6%成员定位在细胞液,6%成员定位在线粒体,1.5%成员定位在细胞壁,1.5%成员定位在细胞核,1.5%成员定位在液泡,结果暗示GPATs蛋白成员参与不同过程。
2.2 GPAT基因家族系统发育分析
为解读GPAT基因家族成员进化关系,以拟南芥、硬粒小麦和野生二粒小麦构建系统发育树(图1),将76个GPAT基因家族成员大致划分为3类:其中GroupⅠ(5个)约占6.57%,GroupⅡ(16个)约占21.05%,GroupⅢ(55个)约占72.36%,Group Ⅲ中又分为3个小类(Ⅲ-a有21个、Ⅲ-b有8个、Ⅲ-c有26个),3个植物GPAT基因家族成员在3个大类中的分布见表2。
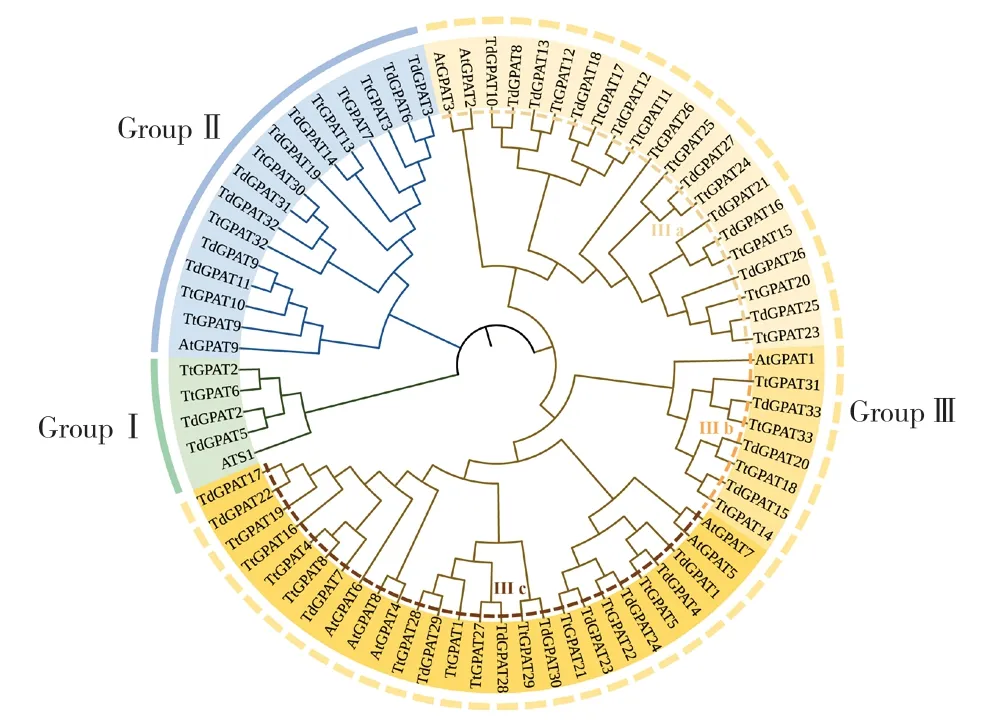
图1 拟南芥、硬粒小麦、野生二粒小麦GPAT基因家族进化分析

表2 GPAT基因家族成员在系统发育树的分布情况
2.3 基因结构和保守结构域分析
为了进一步研究GPAT蛋白质的结构域,以此推测相同结构域可能具备的功能,利用MEME在线软件对GPAT成员蛋白序列进行分析,如图2A所示,同一分组的大多数家族成员具有相似的Motif构成,GroupⅠ和GroupⅡ的Motif缺失较为严重,而GroupⅢ几乎涵盖所有Motif,所有家族成员均含有Motif-2,推测该Motif可能具有特别的生物学功能。其中,Group Ⅰ除硬粒小麦GPAT蛋白成员外其他成员仅含有Motif-2。GroupⅡ所有GPAT蛋白成员均含有Motif-1、Motif-2,部分GPAT蛋白成员有Motif-3、Motif-5、Motif-7、Motif-9。

图2 GPAT蛋白质保守基序(A)、保守蛋白结构功能域(B)和基因结构分析(C)
同时,从图2-B可知,GPAT家族成员中共检测到7个保守蛋白结构功能域,其分组与系统发育树的聚类分析相似,在保守结构域方面进一步支持了GPAT基因家族分类。其中,GroupⅠ家族成员蛋白结构域较为保守,在组内成员中均发现PLN02349结构域;GroupⅡ组内成员保守结构域多样性较高,鉴定到PLN02833、LPLAT_LPCAT1-like、LPLAT结构域;GroupⅢ中发现PLN02177、PLN02499、PLN02588结构域。
硬粒小麦与野生二粒小麦GPAT基因家族结构大致可以分为2类,这与大麦和青稞在基因结构方面表现相似[8,10]。如图2C所示,GroupⅠ和GroupⅡ内基因的内含子较多,大多在7~12个之间,硬粒小麦中TtGPAT2、TtGPAT6、TtGPAT30、TtGPAT32,野生二粒小麦中TdGPAT2、TdGPAT11、TdGPAT31、TdGPAT32的内含子最多,由12个内含子组成;GroupⅢ内的基因结构较为简单,插入的内含子较少,大部分在1~3个之间。
2.4 共线性分析
对拟南芥、硬粒小麦和野生二粒小麦进行共线性分析,结果表明:硬粒小麦和拟南芥基因组中仅有2个基因存在共线性关系,硬粒小麦和野生二粒小麦的共线性基因对数目为66个,说明四倍体小麦组间关系较为亲密(图3)。
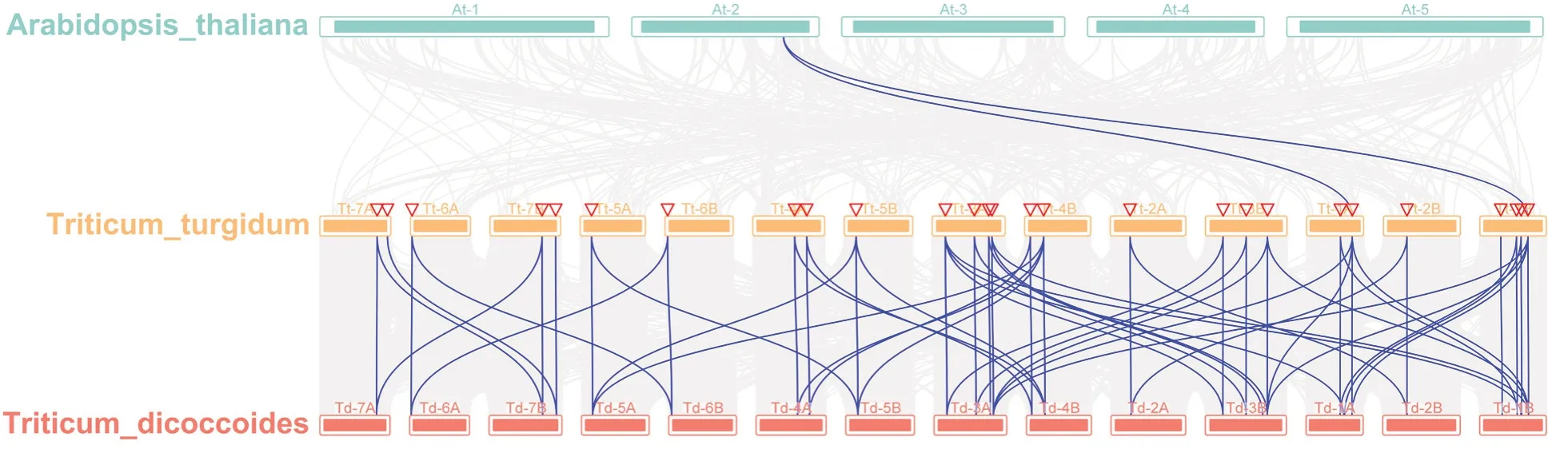
图3 GPAT基因在拟南芥、硬粒小麦和野生二粒小麦间的共线性分析
3 讨论
本研究从硬粒小麦和野生二粒小麦中各鉴定到33个小麦GPAT基因家族成员,在数量上超过了大麦、青稞等其他禾本科作物。理化性质结果表明:GPAT成员氨基酸长度为149(TdGPAT19)~589(TtGPAT1),分子量为16.22 kDa(TdGPAT19)~62.76 kDa(TtGPAT1),等电点为5.92(TdGPAT5)~11.39(TdGPAT7),但绝大多数蛋白(56个)的pI大于8,说明大多数GPATs蛋白富含碱性氨基酸,这与其他物种相似。
系统进化分析表明,四倍体小麦GPAT家族成员可分为3个分支,与拟南芥、玉米、大麦等其他物种研究结果相似[6-7,10],硬粒小麦和野生二粒小麦GPAT家族成员分布在3个分支的基因数量几乎一致,表明四倍体小麦组间相似性较高。结合基因结构分析,GroupⅠ和GroupⅡ内大部分成员含7~12个内含子,GroupⅢ内大部分成员仅含1~3个内含子,表明相同亚家族中GPAT基因有着相似的基因结构,且其进化进程和基因功能也具有一定的相似性。共线性分析显示,硬粒小麦基因与拟南芥的共线性较低,但是与野生二粒小麦的共线性关系较高,说明四倍体小麦之间亲缘关系更近。
