温度胁迫对烤烟生理机制的影响及相关调控研究进展
2023-10-29叶晓青曾嘉楠李俊业李淮源邓世媛陈建军王媛媛
叶晓青,曾嘉楠,邹 勇,周 诚,李俊业,李淮源,邓世媛,陈建军,王媛媛
(1.深圳烟草工业有限责任有限公司,广东 深圳 518109;2.华南农业大学农学院,广东 广州 510642)
温室效应导致极端天气事件频繁发生,严重制约作物生产。烟草(Nicotiana tabacum)是重要的经济作物和模式作物,为一年生或有限多年生温度敏感型草本植物[1],极易受到环境因素影响。烟草生育前期若遭遇低温胁迫,将导致烟株发育加速,叶片未积累足够物质,提前由营养生长转为生殖生长,即“早花现象”。生育后期若遭受高温胁迫,将缩短正常大田生育期以及诱发各类病害,导致烟叶减产,工业可用性降低,严重制约优质烤烟生产。
在烤烟田间生产中,温度胁迫作为环境胁迫的表现形式之一,给国内各烟区带来不同程度影响[2]。在我国广东韶关、梅州烤烟种植区[3],每年2—3 月北方冷空气南下,常造成种植区气温急剧下降,最低温度低于8 ℃,使烟株生长前期气温偏低,难以稳定在正常生长所需的13 ℃,进而导致早花现象。数据显示,1998 年广东烟区低温导致2 万hm2烤烟中50%出现早花。此外,处于亚热带季风气候的广东烟区,烤烟生长后期往往面临高温高湿天气,不仅使大田生育期被迫缩短至100~105 d,造成上部烟叶非正常成熟,还会诱发烟株染上青枯病等根茎类病害。我国广西靖西县烤烟种植区也发生因温度胁迫导致的病害爆发情况,该地区5 月中下旬常处于30 ℃以上的高温天气,极端高温气候引起高温高湿型病害爆发,如烟草黑胫病、青枯病,进而导致烟叶减产、工业可用性降低[4]。处于中亚热带东部温润气候区的湖南烟区[5],同样也受极端气温灾害影响:春季湘中烟区常处于低温寡照,导致烟株出现“早花”现象;夏季湘南烟区高温频发,引起烟田出现“高温逼熟”现象,造成烟叶优质与高产矛盾比较突出。鄂西南烟区春季常处于低温阴雨天气,使烟株生长环境温度过低,烟株前期出现早花现象;夏季湖北西部烟区由于海拔高度较低,使烤烟生长后期常遭遇高温热害,严重影响烟叶风格特色形成,易造成烟叶产质量大幅下降[6]。温度胁迫对西南产区的烟叶品质也产生显著影响,在平均海拔最高的云南烟区,中后期常多雨寡照、气温较低,导致烟株生理代谢减弱,造成致香型物质种类和含量不同,蛋白质和总氮含量下降,影响烟叶产质量[7-8];位于中海拔的重庆烟区,移栽期时段的平均温度为11~18 ℃,导致烟株生长缓慢,进而影响后期烟叶生长发育以及品质形成[9]。
综上所述,生育温度胁迫严重影响国内烟草生长、制约优质烟叶生产。如何增强烟草自身应对温度胁迫的抗逆性,改善烟草抗逆栽培措施已成为国内烟草行业稳定发展、国家烟草种植可持续发展的重要研究课题。本文重点回顾了近年来温度胁迫对烤烟生长的影响,针对高温热害及低温冷害对国内烤烟产质量的影响及其影响机制进行综述,为筛选抗(耐)温度胁迫的烤烟品种、改善栽培措施等提供理论参考。
1 温度胁迫对烟株形态及生理的影响
1.1 温度胁迫对烤烟生长发育的影响
烤烟的生长发育阶段主要分为苗期(播种期、萌发期、苗床期)、移栽期、还苗期、伸根期、团棵期、旺长期、现蕾期、打顶期、成熟期,且不同生育期的适宜温度及对温度胁迫的响应不同(表1)。

表1 各生育期烟株在温度胁迫下的响应Table 1 Effects of temperature stresses at different stages on Flue-cured Tobacco
1.1.1 低温胁迫 烤烟在苗期常遭遇冷害[10],冷害一方面抑制幼苗生长,另一方面诱导早花,导致优质烟叶减产。在种子萌发期,适宜的室温(25~28 ℃)是烤烟种子正常萌发的必要条件。研究发现,将种子进行短期低温前处理有助于打破种子休眠、促进种子萌发、提高种子萌发率,若萌发温度长期低于8 ℃则会抑制种子发芽,且种子发芽持续时间随温度降低而延长[11]。在烤烟苗期,最适生长温度为25~28 ℃,此时期遭遇1~2 ℃的低温易造成幼苗死亡,10~13 ℃低温会使烟草生长停滞,而16~17 ℃以下的低温会导致成熟烟叶的品质低劣[12-13]。在广东韶关、梅州、连州烟区[3],烤烟移栽时正值早春时节,此时土地还未回温,较低的土壤温度不利于烟苗发育,易造成烟株生长缓慢,老根颜色变褐、变黑且无新根长出,茎杆木质化,蹲苗期延长,烟株进入还苗期和伸根期时间延后,进而影响烟草的团棵期和旺长期。若此时再遭遇寡照、地表结霜等灾害,会造成烟苗晚发、生长近乎停滞等现象。在云南烤烟育苗期,遇到“倒春寒”天气,导致烟苗心叶叶片出现黄斑、叶片变薄、叶片边缘内卷或舌状伸展等现象,而遭遇严重冷害的烟苗则会出现畸形、矮化,甚至造成生长停止[14]。相关研究显示,云南、贵州、四川等高海拔烟区,在烤烟成熟期常遭遇低温胁迫,造成烤烟生理代谢强度减弱,抑制烟碱以及干物质形成,影响烟叶品质[7-9]。
低温诱导“早花现象”,使采收时期的烟株留叶数降低、产能下降:烟株在6~12 片叶期,若气温低于13~18 ℃且持续10~20 d,会引起早花株率显著增加,且留叶数减少。正常烟株留叶数为18~22 片,而“早花”烟株仅有13~15 片或更少[13]。
1.1.2 高温胁迫 温度高于30 ℃,烤烟种子萌发速度减缓;高于35 ℃,已萌动种子活力降低。苗床期时,温度高于30 ℃容易发生烧苗,叶片暗黄褶皱[13]。在团棵期至旺张期,多数烟区(湘南、广西靖西、湖北西部)[4-6]持续干旱高温,烟株长期遭受高温胁迫,导致上部叶叶尖萎蔫下垂、枯焦,主脉一侧叶肉组织萎蔫、黄化、扭曲、畸形或呈枯斑,严重时导致数片新生叶干枯、歪头,整株萎蔫甚至死亡。烤烟在生育后期也常遭受高温胁迫[15],即烟田温度≥35 ℃持续3 d 及以上时,烟株生长速度变缓,此时干物质消耗量大于积累量,干物质积累减少;而烟叶落黄速度加快,各部位烟叶落黄层次减弱,甚至出现白化,即“高温逼熟”现象[16]。此现象的发生使烟叶品质严重下降,降低工业可用性以及烤烟生产经济效益。“高温逼熟”现象在水稻、小麦作物中也常出现,导致叶片发白、千粒重降低、产量及籽粒品质降低等[17-18]。
1.2 温度胁迫对烟株光合呼吸的影响
1.2.1 低温胁迫 烟株在冬季育苗过程中常遭遇低温冷害,导致许多生理过程发生紊乱,其中对光合作用的抑制效果明显。低温直接影响叶绿素的合成和光合器官的活性[19-20],首先是叶绿体形态发生改变,膜系统被破坏,进一步影响光合作用的主要环节(水气交换通道、光合电子传递及碳同化等),严重限制光合产物的形成和能量的产生。随着低温胁迫时间的延长,叶片内叶绿素含量呈下降趋势,且叶绿素a/叶绿素b 增大[21]。低温胁迫下叶片气孔导度和蒸腾速率显著降低,说明低温中烟叶内正常气体交换受到限制,阻碍CO2供应,造成胞间CO2浓度降低,进而影响叶片的光合速率;叶片净光合速率随温度降低而下降,且在8、3 ℃条件下发生低温光抑制现象[19]。
烟株对低温环境的耐受性在很大程度上取决于植株对于活性氧(Reactive Oxygen Species,ROS)的清除或防护能力。交替呼吸途径在降低植物细胞ROS 中发挥重要作用,但低温环境会抑制植物线粒体内交替呼吸途径,从而加速ROS 的积累,损伤光合机构,降低烟株光合利用能力,光反应系统电子传递受阻,降低碳同化能力和同化力(ATP 和NADPH)的积累[22-23]。
1.2.2 高温胁迫 在植物响应高温胁迫的研究中,普遍认为光合作用是植物对高温响应最敏感的生理过程[24-26]。烤烟叶片内光合作用在高温热害下遭受明显限制,在同等光照强度下,高温处理的云烟87 上部叶蒸腾速率、气孔导度和光合强度大幅下降,参与叶黄素循环的两个关键酶(紫黄质脱环氧化酶、玉米黄质环氧化酶)活性均呈下降趋势;可见,高温对植株光合作用的破坏程度,取决于高温对叶片光合相关酶活性的钝化程度。在中度高温胁迫后植物光合碳同化过程首先被抑制,主要原因是高温导致核酮糖二磷酸缩化酶(Rubisco)的活性降低,但光系统Ⅱ(PhotosystemⅡ Complex,PS Ⅱ)的功能受到影响较小。当温度升高到一定程度,光系统Ⅱ会遭受不可逆伤害,类囊体膜结构破损,电子传递发生紊乱。研究发现,30~45℃高温处理15 min 后叶片叶绿素荧光参数显著降低,光系统Ⅱ反应中心活性受到抑制[27]。而长期处于高热环境将造成细胞、叶片乃至植物的死亡[28]。
高温胁迫直接影响呼吸酶活力,从而降低呼吸强度。研究发现,在高温处理条件下,烤烟品种云烟87 的胞间CO2浓度随处理时间延长呈先下降后上升再下降的趋势,说明长时间的高温胁迫导致烟叶呼吸作用变弱[29]。在其他作物上的研究证实,不同品种对高温胁迫的耐热型与叶片呼吸强度有关,耐热性越弱,叶片呼吸速率的降低幅度越大。另外,呼吸作用可以清除一些由于高温产生的有害物质,减轻高温逆境危害。例如,耐热性较强的黄瓜品种在高温胁迫下降低呼吸作用,减少对有机物的消耗,维持代谢平衡[30-31]。
1.3 温度胁迫对烟株碳氮代谢的影响
烟株内的碳、氮代谢过程互为基础、相互促进,前期旺盛的氮代谢和营养生长是保证后期旺盛的碳代谢和营养物质积累的基础,而适宜的生长温度是决定碳氮代谢平衡的关键因子之一。
当烟株生长过程遭遇低温胁迫时,会加大气孔对CO2的扩散阻力,减慢光合产物的输送速度,进而抑制叶片的光合作用,打乱碳代谢的平衡[32];烟株体内细胞环境发生变化,细胞趋于氧化态势,提高蔗糖转运蛋白装卸能力,加快叶片顶端分生组织输送蔗糖,启动成花诱导[33],造成“早花现象”。在氮代谢方面,低温胁迫降低叶片中大部分参与氮代谢的关键酶活性,影响根系对氮素的吸收,进而减少叶片中的氮代谢物质的积累,影响氮代谢的系列反应[34]。当然,植株本身对逆境适应的生理机制也会对低温胁迫作出反应。研究表明,低温促使植物可溶性蛋白积累加快,而可溶性蛋白含量的增加可以束缚更多水分,降低原生质因结冰而损伤致死的可能。在低温胁迫下,植株内游离氨基酸迅速积累,提高了细胞液浓度,对细胞起保护作用[35]。此外,由于具有强亲和性,游离氨基酸一定程度上提高了原生质的保水能力和胶体稳定性[36]。
高温胁迫主要通过钝化烟叶内的碳氮代谢相关酶活性,从而影响烟株正常生理活动。高温常伴随强光的发生,叶片硝酸还原酶(Nitrate Reductase,NR)活性降低,烤烟对硝态氮的利用率降低,说明高温强光一定程度上导致烤烟氮代谢出现不协调现象[37]。但当烟株处于高温逆境时,烟株通过产生更多的可溶性糖、脯氨酸、甜菜碱等渗透调节物质,用以维持烤烟的正常生长[38]。
综上所述,烤烟在遭遇外界极端温度时,烟株体内参与碳氮代谢的淀粉酶、蔗糖转化酶、硝酸还原酶等关键酶活性受到抑制,蛋白质合成受阻,影响正常的碳氮代谢过程,造成烟叶产质量发生不同程度下降;在一定范围内的温度胁迫下,植株本身会对胁迫作出响应,以适应环境。
1.4 温度胁迫对烤烟抗氧化系统的影响
无论是高温胁迫还是低温胁迫,均会影响植株内的抗氧化酶活性,细胞内ROS 产生和清除的动态平衡被打破,造成ROS 的过量积累[39-41],引起膜脂过氧化,致使膜脂和膜蛋白受损,膜透性增大,膜结构完整性破坏,最终导致蛋白质和核酸变性,使植株发生代谢紊乱[42]。即使短暂的热刺激也能促使ROS 含量升高。其中O2-、H2O2是ROS 的重要组成部分,在高温强光胁迫下烟叶H2O2含量显著升高,叶片内膜脂过氧化产物丙二醛(MDA)含量增多,且超氧化物歧化酶(SOD)、过氧化物酶(POD)活性升高,过高的POD 活性促使POD-H2O2分解系统参与叶绿素降解,从而降低叶绿素含量[37,43]。但不同烤烟品种的抗氧化系统对温度胁迫的响应不同,其耐寒性和耐热性也有所差异。研究表明,耐寒烤烟品种K326 的存活率显著高于敏感型烤烟品种云烟203,SOD、POD、CAT 活性均高于云烟203[44];抗寒性较差的品种其保护性酶活性呈先升高后降低的趋势。
综上所述,烟株在应对不同温度胁迫对生理形态、碳氮代谢、光合作用以及抗氧化系统的生理机制响应方面存在异同性(图1)。

图1 温度胁迫对烤烟生长的影响及其影响机制Fig. 1 Effects and mechanisms of temperature stresses on growth of Flue-cured Tobacco
1.5 温度胁迫对烟株植物蛋白和基因表达的影响
温度胁迫会扰乱植物细胞内的蛋白质稳态。当烟株受到热胁迫时,细胞壁的结构和性质会发生改变,此时改变了膜的流动性和通透性,促进外胞体Ca2+的释放。ROS 和NO 作为第二信使,迅速激活下游调控网络。高温胁迫将导致细胞质内Ca2+、ROS 和NO 增加以及错误折叠或未折叠蛋白质的过度积累,对植物细胞产生毒性[45]。
面对失衡的细胞体系,植物已经进化出应对温度胁迫的各种反应,以尽量减少细胞损伤,包括快速合成热休克蛋白(Heat Shock Proteins,HSPs),即通过稳定和帮助重新折叠热失活蛋白来保护细胞免受热损伤的分子伴侣。热应激诱导细胞质热应激反应(Heat Shock Response,HSR),其中热休克转录因子(Heat Shock Transcription Factor,HSF)激活一系列编码热休克蛋白的基因。研究表明,在高温胁迫下超表达热休克蛋白基因家族的ZmHSP16.9[46]、RcHSP17.8[47]、BcHSP70[48]、LeHSP21 等[49]通过增加烟草种子发芽率、叶绿素含量和抗氧化特性,维持光合作用水平,降低MDA 含量和电导率,均表现出显著的热胁迫抗性。此外,MsDJLP基因在抗寒性和耐热性中均发挥正向作用[50]。
伴侣蛋白编码基因占热激响应基因的比例较低,更多的HSR 基因编码参与信号传导、转录调节、各级代谢过程[51]。超表达脱氢蛋白基因cardhn[52]、非特异性脂质转移蛋白基因NtLTPI.38[53]、调控因子ClpB 蛋白基因[54]和扩张蛋白基因PpEXP1[55]均通过减轻烟草的光合损伤、改善渗透调节和增强抗氧化能力调节烟草的光合能力和抗氧化能力,正向调节烟草的耐热性。此外,超表达转录激活因子转CAP2 基因[56]、植物发芽延迟家族基因NtDOG1L-T[57]和热休克转录因子(HSF)基因BcHsfAl[56]的烟草幼苗的启动子活性在热胁迫下被显著激活,热休克蛋白和热休克因子基因的表达水平也相应提高。N-乙酰谷氨酸激酶ZmNAGK 通过激活抗氧化相关防御信号,如抗坏血酸过氧化物酶2(Ascorbate Peroxidase 2,APX2)和超氧化物歧化酶C(Superoxide Dismutase C,SODC)以及热休克网络基因来增强植株耐热性[58]。
2 温度胁迫下烤烟抗逆调控措施
2.1 水肥调控
水肥管理调控对烟叶产量结构有重要影响。高温条件下进行合理灌溉,在降低地温的同时能促进烟叶对氮磷钾养分的吸收[59],提高烟叶碳氮代谢关键酶活性,从而缓解高温对烟株的损伤[60-64]。
硅、钾元素是一种调节性元素,能够调节植物的生长发育,提高抗逆胁迫能力[65-67]。研究发现,增施钾肥可保持原生质膜的稳定性,降低膜渗透率,叶片萎焉率大幅下降[68],有效增强烟株的抗性。此外,在低温胁迫下增施锌肥也能显著提高作物根系活力,减轻低温对细胞膜结构的破坏,同时促进氮与干物质积累,有效缓解营养生长期低温对作物的伤害[69]。因此,在温度胁迫时适当增施部分功能抗逆性肥,有助于提高烟株抗逆性、维持烟株正常生命活动、降低经济损失。与增施单一的功能性肥料相比,复配硅钾肥的施用展现出更好的抗寒性效果[70],也可以综合考虑作物所处的非生物胁迫因素作出施肥调控措施,以达到稳产优质生产目的[71]。
2.2 外源物质调控
与培育耐胁迫品种、基因编辑技术等分子调控手段相比,通过喷施外源物质来缓解温度胁迫效应具有操作成本低、见效快的特点,应用前景更广。目前,多种外源物质处理被证实能通过维持抗氧化系统和光合作用水平,恢复细胞组织结构,从而有效缓解或恢复温度胁迫对烟株的损伤。外源物质因对温度胁迫下烟株的作用机理存在差异,其产生调控效果有所差异(表2)[72-95]。
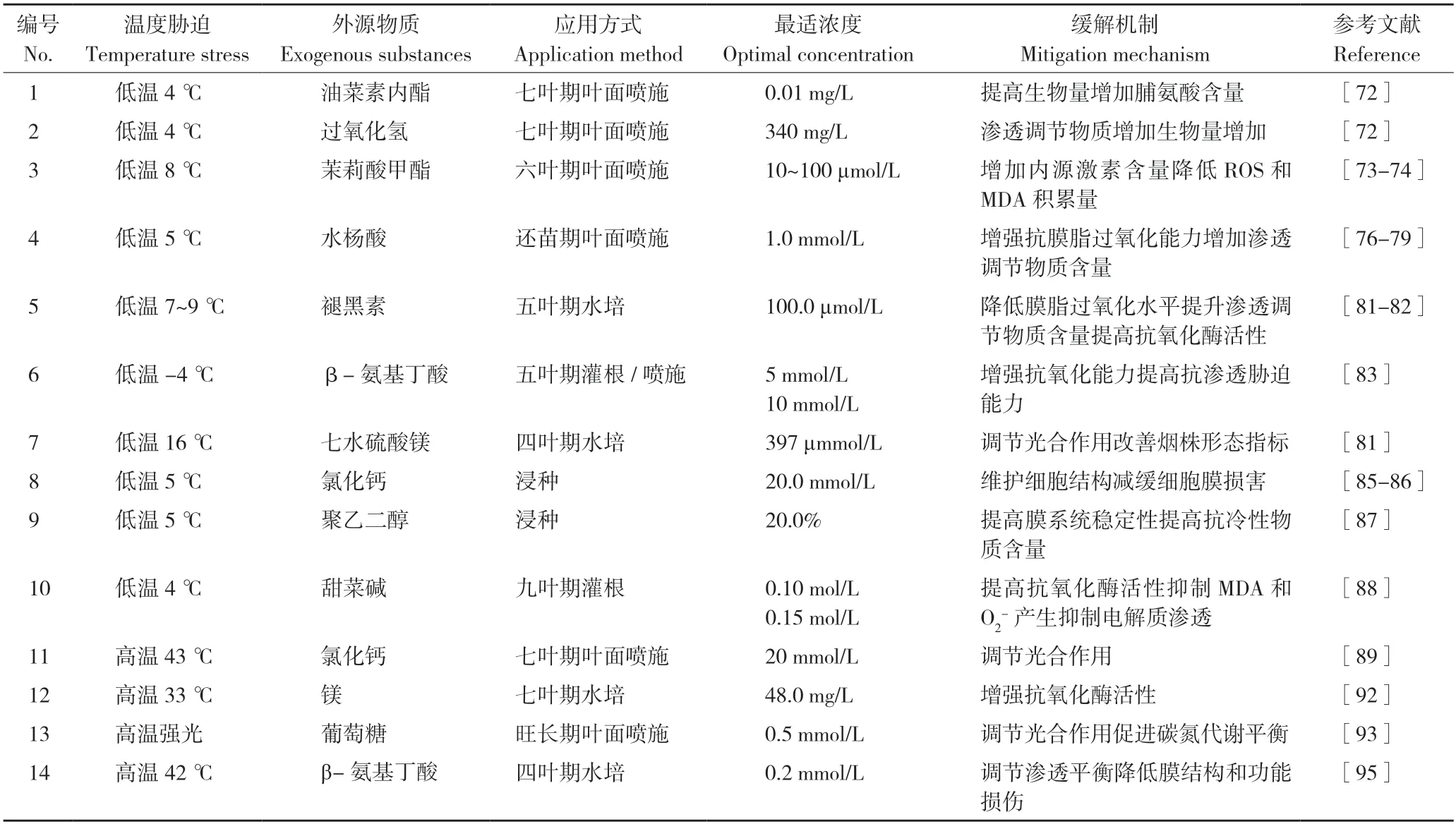
表2 不同外源物质对温度胁迫下烤烟的缓解作用Table 2 Alleviating effects of exogenous substances on Flue-cured Tobacco under temperature stresses
2.2.1 缓解低温胁迫 许金亮等[72]研究发现,喷施油菜素内酯(0.01 mg/L Brassinolide,EBR)能显著恢复烤烟幼苗在冷害中的生长生理进程。前人通过外源施加茉莉酸甲酯(Methyl Jasmonate,MeJA)提高烟草幼苗光合色素的含量,提高抗氧化酶活性,加速抗坏血酸-谷胱甘肽循环(ASA-GSH),降低ROS 和MDA 的积累量,增加可溶性糖和蛋白质的积累,提高内源激素脱落酸(Abscisic Acid,ABA)和生长素(Auxin,IAA)含量,增强烟草抗寒性[73-74],但施加MeJA过量则会导致烟草叶形指数增大、叶片狭长、降低烟叶的商品价值[75]。
水杨酸(Salicylic Acid,SA)是一种植物内源生长调节剂和信号分子,主要从诱导抗氧化防御系统、调控激素稳态和代谢物质等方面缓解非生物胁迫[76-79]。当烤烟遭受低温危害(5 ℃)时,通过喷施不同浓度的SA 可提高叶片中抗膜脂过氧化能力,从而抵御低温危害[80-81]。研究表明低温胁迫下施用适宜浓度的外源褪黑素,可抑制烟草幼苗叶片中ROS 和MDA 的积累,烟叶内脯氨酸、可溶性糖和可溶性蛋白含量增加,膜脂过氧化水平降低,从而提高烟草幼苗抵御低温胁迫的能力[81-82]。此外,外源β-氨基丁酸和镁处理均可有效缓解烤烟低温胁迫的损害[83-84]。
除通过维持光合作用、提高渗透调节物质和抗氧化能力来抵御温度胁迫的伤害外,还可通过稳定细胞膜结构、调控渗透平衡,达到提高抗性的效果。研究表明,CaCl2浸种后烤烟幼苗中结合钙含量明显提高,细胞膜结构稳固性得到提高,烤烟幼苗在低温下损伤有所缓解[85-86];聚乙二醇(PEG)分子能改变各类细胞的生物膜结构,在低温胁迫(5℃)下20%聚乙二醇(PEG)浸种可通过保持烟草幼苗膜系统稳定性提高烟苗抗冷能力[87]。甜菜碱(GB)是一种次生代谢物,在调节渗透平衡中发挥重要作用。顾开元等[88]发现,4℃低温胁迫下,将红花大金元根施0.15 mol/L GB、K326 根施0.1 mol/L GB 能显著缓解低温损伤,提高烤烟耐寒性。此外,将烤烟幼苗喷施外源过氧化氢(340 mg/L H2O2)也能显著提高烟株渗透调节物质含量和增加生物量,从而有效缓解冷害中幼苗的损伤[72]。
2.2.2 缓解高温胁迫 钙对细胞膜、细胞壁的结构功能具有重要维护作用,且作为第二信使的Ca2+参与植物对外界的反应与适应,调节植物细胞对逆境胁迫信号转导过程[89]。研究发现,外源钙和氯化钙均对高温逆境下的烤烟光合作用有重要调控作用[90]。高温43 ℃胁迫处理2 h,外源氯化钙可改善烟叶气孔导度、提高出氧复合体的热稳定性,活性氧积累因此减少,从而达到缓解效果[84]。适当施镁(48.0 mg/L)可通过增加CAT、POD、SOD 等抗氧化酶活性,提高烟株的干物质量和叶片叶绿素合成量,缓解高温强光胁迫下烟叶的日灼斑症状[91]。
叶面喷施氨基酸[92]、葡萄糖[93]后,作物在高温胁迫下光合速率仍保持在较高水平,维持碳氮代谢平衡,促进烟株在温度胁迫下的生长。此外,张清莉等[94]发现,外源施加0.2 mmol/L β-氨基丁酸,通过诱导热激蛋白基因 NtHSP101 的表达提高烟草抗高温胁迫能力。
3 展望
目前,我国大部分烟区存在烤烟春季生长环境温度过低,容易引起烤烟出苗质量下降以及早花问题,而夏季环境温度过高,造成烤烟出现高温逼熟、早衰等现象,仍严重制约烤烟的高效生产和发展。
现阶段,温度胁迫对烤烟的影响机制研究较为薄弱,前人研究多集中于烟株表观和生理响应,以及代谢组学、蛋白组学、转录组学等方面,但也存在一些问题,如未深度挖掘温度胁迫对烤烟各代谢过程的影响;烟株生长环境受温度、水分、光强等综合因素影响,目前研究仅停留于单一胁迫对烟株生理的影响;尚未在烤烟遭受温度胁迫前作出预测响应;针对温度胁迫对烟株的内源激素含量及其体内的动态变化极具研究价值,但目前相关研究仍十分有限,且对于各内源物质是否存在互作进而调控温度胁迫下烟株体内的生理体系仍有待探索。
针对目前与其他农作物(玉米、水稻)的研究[95-97]比较,基于提高烤烟在温度胁迫下抗逆性的角度,未来研究可注重以下方面:挖掘地方种质资源,结合多组学技术鉴定耐热基因;温度胁迫与其他胁迫如干旱胁迫、盐胁迫的交叉研究;植烟土壤微生物、植株根系微生物和内生微生物也可能为提高作物耐高温能力提供新思路;随着高通量测序技术的快速发展,可进一步利用多组学或交叉组学等手段对烤烟温度胁迫响应机制进行深度挖掘;发展对烤烟温度胁迫的预测研究。通过量化田间温度与烤烟性状的相关关系,采用模型模拟、近红外光谱等技术建立烟-田监测预警系统,并形成便携式仪器等产品加以推广应用,以最大程度做出抗逆响应;根据烤烟品种特性,结合当地气象因素和田间水肥状况,制定抗逆栽培管理措施,以有效缓解实际生产中温度胁迫对烤烟生长发育的影响,对于应对气候变化下烤烟稳产优质发展具有重要意义。
