采用脑网络分析的应激诱发脑-机接口运动感知与反馈激活时序规律研究
2023-10-29张凯徐光华李文平李辉杜成航韩丞丞张四聪谢杰仁
张凯,徐光华,2,李文平,李辉,杜成航,韩丞丞,张四聪,谢杰仁
(1. 西安交通大学机械工程学院,710049,西安;2. 西安交通大学机械制造系统工程国家重点实验室,710054,西安)
脑-机接口作为一种新型的人机交互技术,可以在不依赖于脑的正常输出通路(外围神经和肌肉组织)的情况下,使人类大脑直接与计算机或其他外部设备进行通信,进而控制外部设备进行相应的任务并提供反馈[1]。其中,作为其重要工程的应用,运动诱发脑-机接口为脑损伤闭环神经协同康复提供了一种可行的技术手段,具体通过刺激范式有效诱发受试者的运动中枢使其产生神经响应信号,再通过信号处理技术对脑电信号进行实时解码,然后将解码得到的运动意图转化为外部设备的控制指令,辅助受试者完成运动辅助和康复训练[2-4]。该过程有效模拟了运动行为控制的中枢响应过程,实现了神经冲动沿着运动感知环路—神经中枢—运动控制环路—受控肢体的传导,并利用物理接口和双向通信技术从大脑外部构建了一条闭环回路,进而实现运动控制环路的重建[5-6]。
基于脑-机接口技术的神经环路重建有效性受到多种因素的影响,其中,运动感知与控制的时序性对于接口性能与应用效能具有重要的影响。Hebb法则与脉冲时间依赖突触可塑性规律(spike-timing dependent plasticity, STDP)表明:活动时序依赖的突触可塑性满足确定性的时间窗口,直接揭示了有顺序的激活突触前和突触后神经元是修饰环路的关键,从微观层面揭示了时序性对神经环路重塑的重要规律[7-9]。然而,传统的诱发范式仅利用依赖主观认知与弱视觉诱发信息来引导受试者产生自主想象[10-12],其不仅易受到生理状态变化的影响,且难以保证运动感知与控制的同步性,易导致运动诱发脑-机接口的中枢响应过程在不同试次、不同时间具有高度变异性,直接造成整个交互过程中,运动感知神经与运动控制神经的激活过程难以保证同步性和有效时序性(如图1所示),无法满足感知反馈同步性和时效性要求,与STDP所揭示的规律相悖。由此,限制了运动诱发脑-机接口的应用性能,难以保证临床应用中的康复训练与治疗效果。
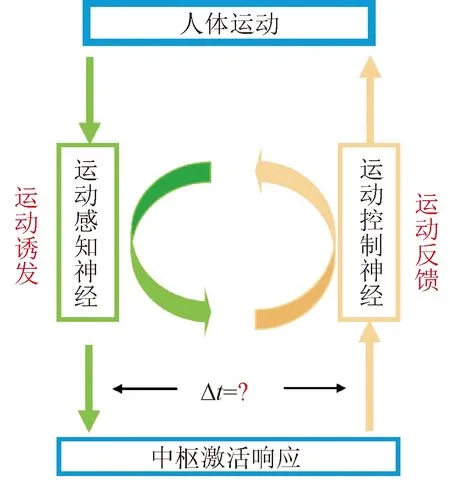
图1 传统运动诱发脑-机接口感知与反馈激活示意Fig.1 The diagram of perception and feedback activation of traditional brain-computer interface based on motor activation
探索运动感知环路与控制环路激活的强匹配诱发模式,对于增强闭环脑-机交互过程的同步性和时序性具有重要启示。中枢激活响应效果往往取决于刺激范式的设计,作为一种典型的刺激模式,应激反应可以定义为各种刺激物(应激源)作用于个体而引起的非特异性反应。特定条件下的应激反应有助于提升自主注意力水平,促进情绪和思维的正向调整,并提升交感神经的激活程度,引起脑电信号功率的增强[13-14]。文献 [15]研究表明,特异性的重心失衡应激刺激会引起运动中枢的激活,且该激活方法具有更强的激活程度和更有序的激活效果。因此,应激状态的出发时刻引起受试者的必然性运动响应,为脑-机交互中运动感知环路与反馈控制环路的有序激活规律研究提供了一种可行的方法。同时,研究发现,大脑的信息加工和处理过程可以通过特定脑区之间信号同步强化效果来进行表征,其中,研究驱动源脑区和神经有向连接的相关规律对于理解大脑活动和神经调控的时序规律有着重要的作用[16]。脑功能网络,是一种将图论和复杂网络理论应用于大脑活动定量分析的研究,作为一种描述大脑皮层不同功能区相互作用一种方法,可以有效帮助研究人员进一步了解脑内信息的加工机制和作用规律[17]。
因此,本文利用应激诱发脑-机接口技术与脑功能网络分析手段,探索了运动激活响应的时序作用规律,研究了闭环脑机时序同步强化方法,为时序同步的脑-机接口设计提供有效理论与技术基础。
1 应激诱发的时序激活响应检测平台设计与搭建
1.1 应激状态触发与运动响应关联分析
应激诱发运动响应通常产生于受试者从非应激态转变为应激态的瞬时时刻,此时刻对应的应激参数定义为应激触发点(用失衡角度θ表示)。重心失衡作为一种典型的运动应激刺激,往往伴随着前庭神经的激活,其作用过程可以描述为:前庭感受器受到偏离重力方向的信息时,神经中枢通过调节四肢的肌张力或控制肢体运动来调整身体的姿态和体位,以维持身体的平衡。如图2所示,基于之前研究的重心失衡应激脑-机接口模型[15],受试者会利用肢体进行支撑动作,来防止重心的完全失衡,而这种运动本质上受到本能行为和自主意识的共同支配,因此具有确定的运动激活性。因此,这种运动响应的强关联模式,有望提供一种可控型意念诱发模式。

(a)重心失衡应激平台
(b)非应激态

(c)应激态
1.2 运动感知与反馈控制激活的时序信息检测平台设计
在运动调控的过程中,有效刺激的产生往往伴随着感知神经的激活,运动动作的发生表征了运动控制神经的激活。对于自然的正常神经环路,运动感知与控制环路的激活几乎是瞬时同步的,而脑-机接口构建的人工神经环路中,两者的激活关系往往不同步,且缺乏有效的激活监测手段。
在真实运动任务下,真实运动的感知与控制反馈近乎同步,因此受试者在应激触发点位置产生的真实运动会引起运动感知与反馈神经同时激活。对于脑-机交互任务,利用外界刺激诱发运动中枢,通过提取脑电激活特征来实施运动控制。由此可以类比得到,有效刺激感知输入的时刻为运动感知环路的激活时刻,脑电激活特征的启动时刻为运动控制环路的激活时刻。
由于受试者存在反应延迟及脑电的演变过程,脑-机接口任务中的感知激活与控制激活往往存在一定的时延。因此,通过记录受试者在应激刺激下应激触发点与脑电特征响应点对应的时刻,有望得到脑-机交互过程中运动感知与控制反馈环路激活的相对时序关系。为此,设计了如图3所示的应激触发点与运动诱发激活检测的多源信息采集系统。

图3 多源生理信息采集系统Fig.3 The acquisition system for multi-source physiological information
陀螺仪位姿传感器通过对重心偏移的角度进行采集以捕捉应激点对应的角度和时刻参数,薄膜压力传感器通过对接触力变化进行监测判断肢体真实运动产生的时刻,脑电信号则是从中枢层面反映运动激活响应的动态变化。
为了减小测量过程的误差,对不同传感器之间时序标定进行统一标准化设置,以保证多源信息采集与任务状态的变化一致性。同时,为保证随动座椅与传感器之间的实时通信,采用TCP/IP与串口通信完成系统控制的信号的实时处理,其具体框架如图4所示。
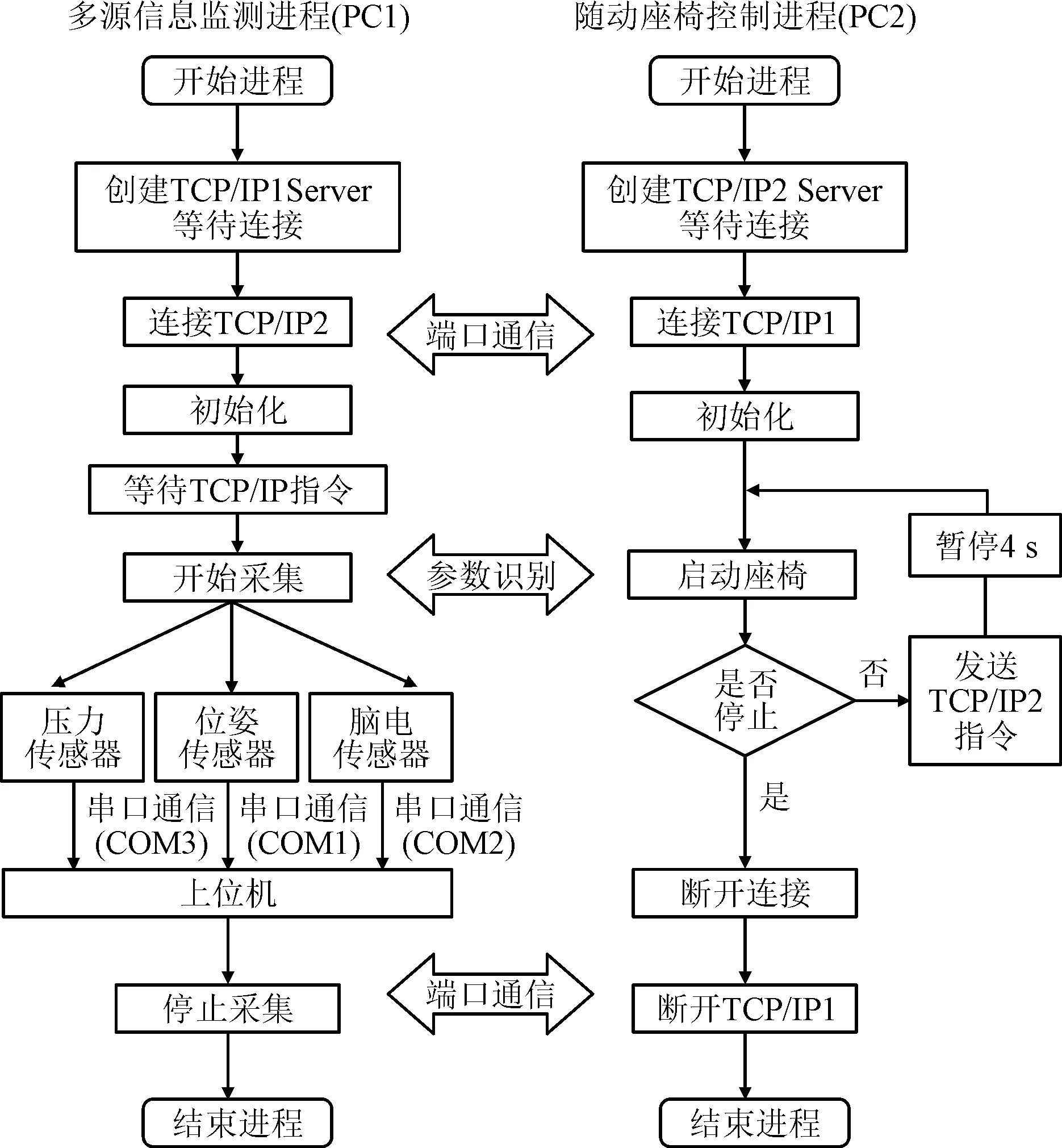
图4 多源信息采集与运动诱发平台软件系统 Fig.4 The multi-source information acquisition and motion induction platform software system
随动座椅与多源信号采集系统采用TCP/IP进行交互通信、接收信号、处理进程并记录受试者生理信号随座椅运动的动态过程。在真实运动或意念激活的信号检测过程中:利用真实运动的接触力变化对应旋转座椅角度变化,可以得到反映感知神经激活的应激点参数;分析计算脑电信号的激活时刻,可以求得运动控制神经的激活时刻。
2 脑-机接口运动感知与反馈激活检测方法设计
2.1 运动应激触发点检测实验设计
为求解应激触发点对应的刺激参数,设计了如图5所示的实验。
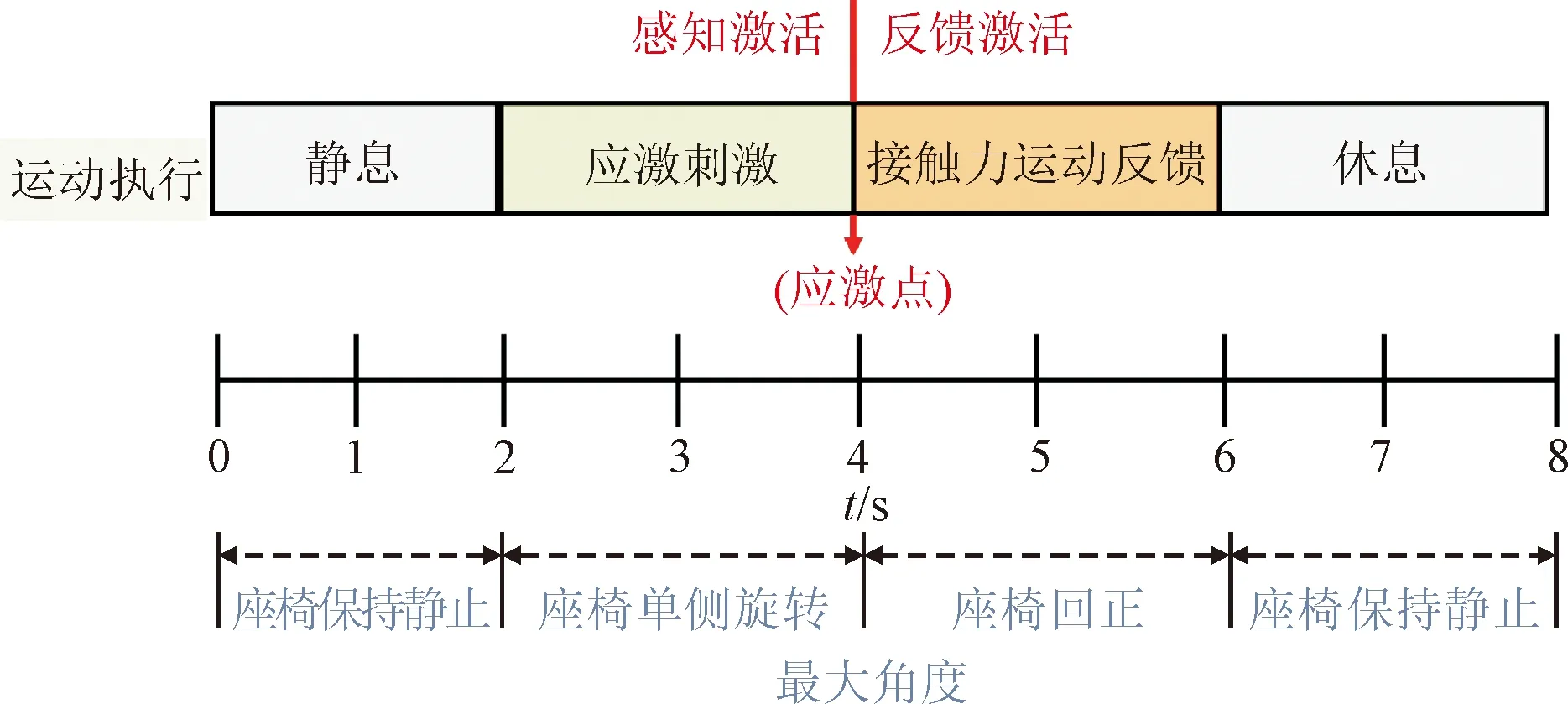
图5 应激诱发-运动执行反馈实验方案示意 Fig.5 The diagram of stress response induction and motor execution feedback
在运动执行实验过程中,前2 s为运动准备阶段,受试者在无物理限位的随动座椅上保持静止。从第2 s起,座椅随机向左/右方向以10 rad/s的速度匀速旋转。期间,要求受试者在无支撑接触力的条件下,尽量保持身体平衡,直到座椅偏转到失衡的临界点,本能反应驱使受试者利用手部支撑座椅扶手,以保持身体平衡,该动作发生过程被布置于支撑点的压力薄膜传感器所记录。在整个过程中,陀螺仪全程记录不同时刻对应的偏移角度。
通过记录支撑接触力产生时刻的对应的偏移角度,即可确定引起应激状态触发的失衡角度θ,即应激点。由于本文的研究对象为左右手运动意念诱发任务,因此通过多次重复左右侧运动偏转实验任务,将叠加平均后的结果定义为单侧手的应激点。
2.2 脑电激活响应检测实验设计
相比真实运动,脑-机交互过程中感知神经的激活通常为外界刺激输入的时刻,控制神经环路激活的判据通常为脑电激活特征的产生时刻,而这种模式下的感知与激活存在一定的时延。
因此,记录应激点产生时刻与脑电特征启动时刻的时延,为量化运动感知与反馈的激活同步关系提供了一种可行的方案。为此,根据运动执行实验得到的应激点参数,设计应激刺激运动诱发实验方案,如图6所示。
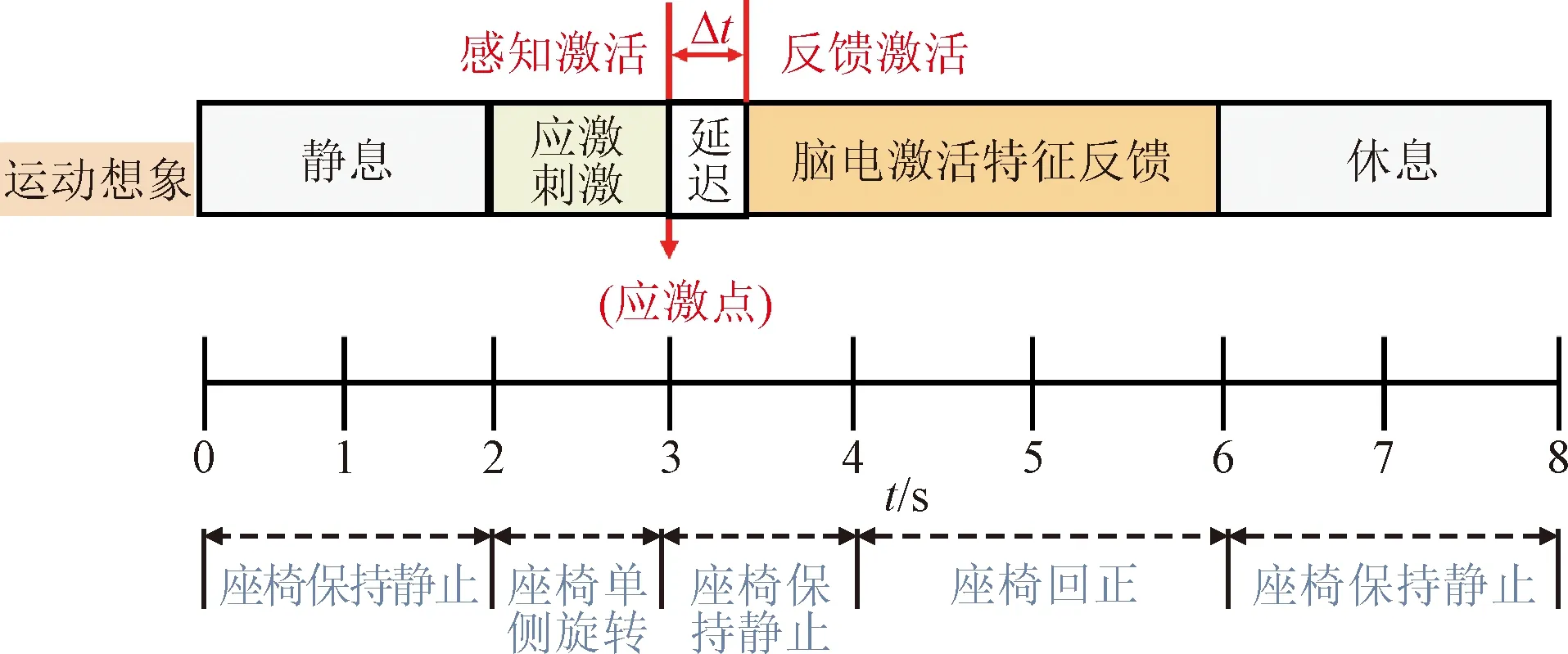
图6 应激诱发-运动激活反馈实验方案示意 Fig.6 The diagram of stress response induction and motor activation feedback
首先,利用运动执行实验所求解的应激点参数对随动座椅的偏移角速度、位移进行设置,具体通过调整随动座椅的偏转角速度ω;然后,从第2 s起,座椅启动并在1 s内偏转到应激触发点的位置,此时采用安全限位装置代替受试者的运动支撑,以保证受试者在实验全程保持静止;从应激点后,受试者执行自主意念诱发任务,直到第6 s,单次实验结束。
2.3 实验条件及脑电数据采集
为了探索时序依赖的脑电响应规律,构建感知同步运动诱发接口,招募了12名受试者(男7女5),身体健康无精神疾病,双眼矫正视力正常,平均年龄(24±2.5)岁。所有受试者都被告知实验的全部程序,并签署知情同意协议。
实验使用奥地利g.tec公司生产的g.Nautilus高精度无线生物电信号采集分析系统,可以用于运动状态下的脑电信号采集,其内置阻抗检测功能可测量电极与头皮接触电阻,精度达 kΩ 级,并具有硬件带通滤波和陷波滤波功能。选取运动区域的相关电位活动作为分析对象,根据10/10 标准电极布置法则,在受试者头部运动皮层相关区域布置电极,其分别位于FC3、C3、CP3、Cz、FC4、C4、CP4、CPz,参考电极位于右耳耳垂(A2),接地电极(GND)位于前额(FPz)。同时,保证所有阻抗保持在5 kΩ以下。脑电图信号的采样率设置为500 Hz,采用50 Hz陷波滤波器和3~100 Hz带通滤波器滤波。在进行信号分析之前,对每个受试者不同任务的脑电信号进行叠加平均,以尽可能减少噪声和异常值对分析结果的干扰。同时,去掉误差较大和不可用的试次,以保证后续分析结果的准确性。
转椅采用数字伺服控制器(Syntron-543AA0),精度为0.1°,配备三轴加速度计(witt-jy901),以记录实际位移运动学。搭载压力薄膜传感器(IMS-S08A),最大量程可达40 kg。所有的运动参数(旋转角度/角速度)都可以手动指定。
实验时,每个受试者要依次参与3类实验:运动执行,应激刺激运动诱发(VSMI)与无应激运动想象(MI)实验。其中,VSMI实验中提供应激刺激和视觉提示,MI实验中仅供座椅旋转的视觉动画作为提示。运动执行实验一共分为3组,每组完成 10次重复实验,每组间隔2 min。两种运动诱发实验各需要完成6组实验,每组开展 10 次重复实验,每组间隔3 min。当受试者在参与脑电采集实验过程中,应尽量避免肢体或者头部运动、吞咽口水、咬牙。实验过程中,实时记录脑电数据、压力数据、位姿数据、时标信息。
3 基于脑网络分析的时序依赖规律量化方法研究
由于EEG能够直接测量神经活动且具有高时间分辨率,探索基于EEG的跨脑区功能连接对理解大脑运动功能的调控机制具有重要的作用。研究表明,特定脑区信号之间的同步强化增强作用可以通过脑功能性连接进行表征[18],而这种连接关系的动态特征能够通过多项指标进行定量评估。在脑电信号分析的领域,研究人员提出了多种算法来定义信号之间的功能连接指标,常见的如皮尔逊相关系数、波谱相干、互信息、相滞指数等[19-20],其中,有向传递函数是一种用多变量自回归模型来表征脑电信号功能性连接强弱的方法,并遵循格兰杰因果关系多变量有效测量规律[21]。其主要通过计算单通道输入信息和输出信息的信息量的关系,来衡量单通道对其他通道的信息贡献量关系,进而定性判断不同脑区响应之间相互联系与影响。本节通过引入脑网络分析方法,利用MATLAB的econnectome工具箱,对应激点诱发下中枢响应时序演化规律进行研究,以揭示应激调控的内在作用机制,验证所提脑-机接口方法对于中枢响应同步性增强的有益效果。
首先,利用多变量的自回归模型(multivariate autoregressive, MVAR)来描述一个N通道的EEG信号[22],X(n)=[x1(n),x2(n),…,xN(n)]。利用静息态和激活态的脑电信号计算激活比E
(1)
(2)
式中:E为激活能量比;A为激活态脑电信号的能量;R为静息态脑电信号的能量;m为运动激活启动时间点;n为单试次总时间点;r=1,2,3,…,p为MVAR的阶数;Ar为系数矩阵;w(n)为高斯白噪声。
然后,将所求系数矩阵与信号表示投影到频域,得到
W(f)=Ar(f)X(f)
(3)
(4)
计算系统传递矩阵D(f)
(5)
式中D(f)元素Dij表示系统中第j个输入与第i个输出之间的关系。
将EEG中第j通道对第i通道的影响关系定义为
(6)

由此,计算单通道的传递函数与所有相关通道的元素平方和,即可得到有向传递函数的表达式
(7)

(8)
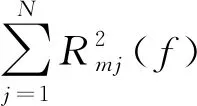
有向传递函数对通道之间信息交流进行了定量的评估,为了进一步挖掘运动激活响应过程脑网络的动态属性,本文采用图分析的方法计算响应的加权网络性质,旨在实现对大脑激活状态时序规律的捕捉。作为一种定量评估脑网络局部信息处理的效率的指标,聚类系数常常被用来表征网络功能分化能力以及评估网络集群程度[23],而运动皮层脑区的激活程度,往往也伴随着局部信息处理效率的增加而增加。因此,选取聚类系数C作为脑激活状态动态评估的指标之一,计算式为
C=
(9)
式中:i、j、h分别为脑网络中的任意节点;ωij为节点i和节点j之间的连通性强度;z为网络节点数;Z为网络节点的集合。
特征路径长度L是一种评估网络功能整合能力的常用指标[24],其定义为网络中任意两个节点之间的平均值。特征路径长度越短,网络功能整合能力就越强,网络连通性越好。特征路径长度计算式为
(10)
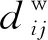
全局网络效率EG作为一种表征网络功能集成属性的指标[25],数值大小与路径长度呈反比,可以更加直观地反映脑网络全局信息的传输能力,计算式为
(11)
作为描述大脑不同脑区之间局部信息传输能力的重要指标[26],局部效率EL可以表征网络功能分离的程度,进而对不同区域活动的规律进行评价,计算式为
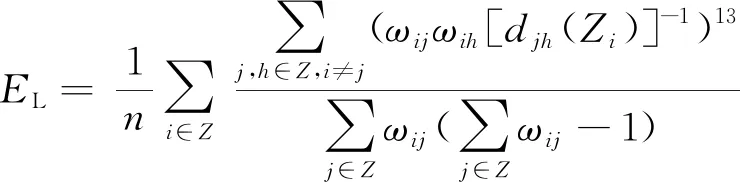
(12)
4 实验与结果
脑网络功能连接能够有效反映运动信息加工处理过程和不同脑区的协同工作机制,以进一步揭示运动行为控制任务下的大脑的工作机制。为此,采用脑网络分析方法进一步探索运动激活响应的动态演化过程。
虽然本文仅对8通道脑电信号进行脑网络分析,但该布置对负责运动皮层的主要通道(FC3、C3、CP3、Cz、FC4、C4、CP4)进行了覆盖,且能保证对运动诱发任务中对侧运动皮层之间的信息交互规律捕捉,因此具有可行性。同时,由于应激刺激对于其他皮层的诱发响应规律尚不明确,为了避免其他脑区激活响应对运动皮层的分析带来干扰,故未扩大通道的选择范围。
4.1 脑功能网络连接分析
脑功能连接反映中枢激活的主要表现形式,可以描述为:核心激活区将作为质心枢纽,与其他相关区域建立连接,协同完成信息加工。在由左右手运动诱发/执行所引起的运动激活过程中,静息态下的两侧脑网络呈现对称态,单侧手运动激活态下的网络呈现偏侧化形态,通常表现为对侧运动皮层激活,与同侧区域建立有向连接。
为了评估本小节所提出方法对运动刺激的激活响应规律,本文通过对应激刺激运动诱发过程的网络连接的动态过程进行可视化,用以分析运动响应的感知同步关系,评估结果如图7所示。

(a)左手运动诱发脑网络激活变化规律

(b)右手运动诱发脑网络激活变化规律

(c)左手运动想象脑网络激活变化规律

(d)左手运动想象脑网络激活变化规律
对同一任务下的脑电信号进行叠加平均,提取μ频段(8~12 Hz)和β频段(13~30 Hz)的脑电信号作为分析对象。如图7(a)所示:在运动准备阶段,左右两侧枕叶皮层产生激活,网络结构呈现对称状态;应激刺激开始,旋转座椅向左侧偏转,运动皮层网络结构由对称态变为偏侧化,右侧运动皮层的CP4、C4通道作为质心通道,控制信息流向其他区域流出,完成对运动信息的协同处理;达到应激点后,停止外源刺激,应激响应占据主导思维活动,此时脑网络连接密度增加,皮层激活逐渐达到峰值;随着激活过程的结束,脑网络由偏侧化恢复到对称态。图7(b)反映了右手运动激活的脑网络变化规律,激活质心为左侧脑区的C3、CP3通道,演化过程与7(a)所示规律具有一致性。由此可以说明,偏侧化重心失衡刺激会诱导受试者产生与运动诱发/运动执行相同的脑网络模式,当失衡点达到应激触发点后,该运动激活引起的网络连接和信息交流会更加密集。由此可以说明,应激触发不仅与传统运动中枢激活具有一致性,而且能有效强化运动脑区之间的信息交流和传递。
4.2 基于脑网络动力学特性的响应规律表征
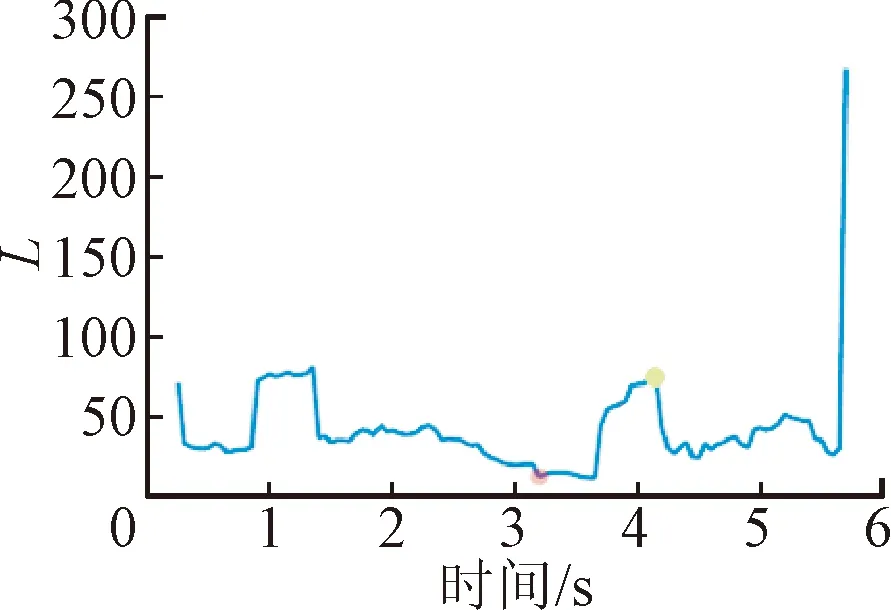
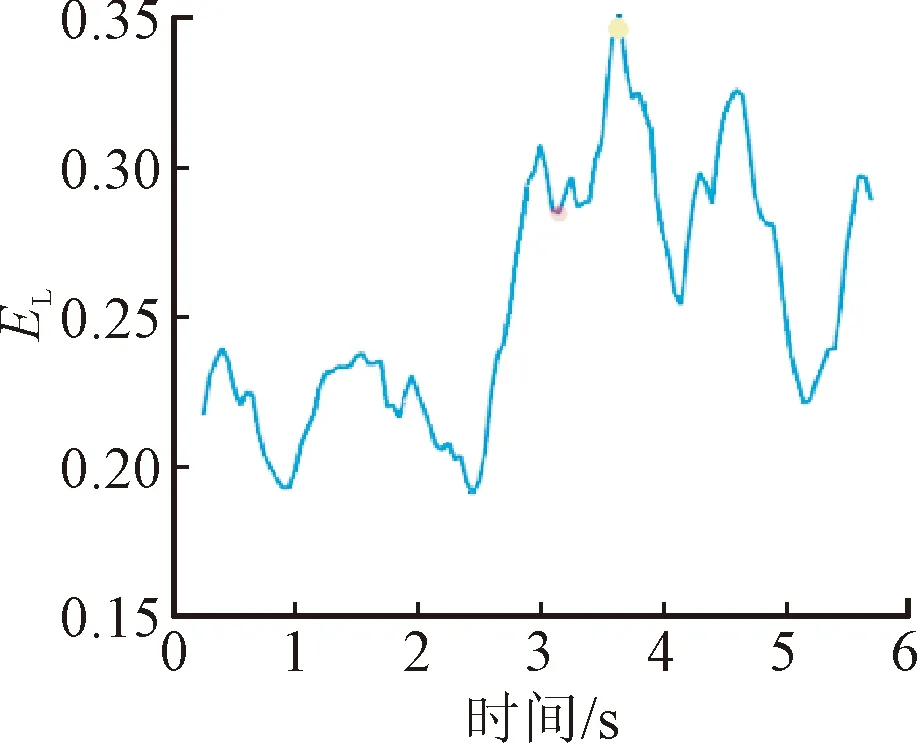
脑网络的拓扑结构可以直观反映中枢响应的演化过程,而网络参数的量化可以实现对中枢响应时序关系的表征。因此,通过量化脑网络的聚类系数、特征路径长度、全局效率与局部效率在诱发任务的动态参数变化来表征运动刺激与中枢激活程度的时序变化规律,具体结果如图8所示。

(a)聚类系数
(b)特征路径长度

(c)全局效率
(d)局部效率
图8给出了1名受试者所有数据叠加平均后的网络参数变化。可以看出,在运动准备阶段(0~2 s),脑网络的聚类程度和信息处理效率较低;随着应激刺激的启动(2~3 s),C、EG、EL曲线先下降而后急剧上升,表明大脑在受到应激事件后,先表现出一定延迟,然后运动皮层之间信息加工在短时间内迅速增加;达到应激点后(3~4 s),运动激活主导此刻的思维活动,该时间段内的C、EG、EL曲线小幅度下降后又持续攀升,并达到峰值,峰值时刻代表了运动皮层激活最强的状态;运动诱发任务结束后(4~6 s),随动座椅回正,大脑信息加工效率逐渐从峰值降低,随着受试者回到静息态,恢复到与运动准备阶段相同的水平。不同于上述3个参数,特征路径长度表征了网络功能整合能力,长度越短,整合能力越强。虽然变化规律与上述3个参数相反,但揭示的规律具有一致性。
4.3 感知反馈激活时序规律的量化统计分析
本小节对12名受试者的应激点参数、激活响应时延以及激活最大持续时间进行统计。应激点参数和运动响应的激活时延由量化分析方法[15]得到,最大激活持续时间通过脑网络动力学参数变化规律得到,即记录激活启动点(图8中红色标识点)与响应峰值(图8中绿色标识点)对应的时间区间。对叠加平均后的信号进行统计分析,结果如表1所示。
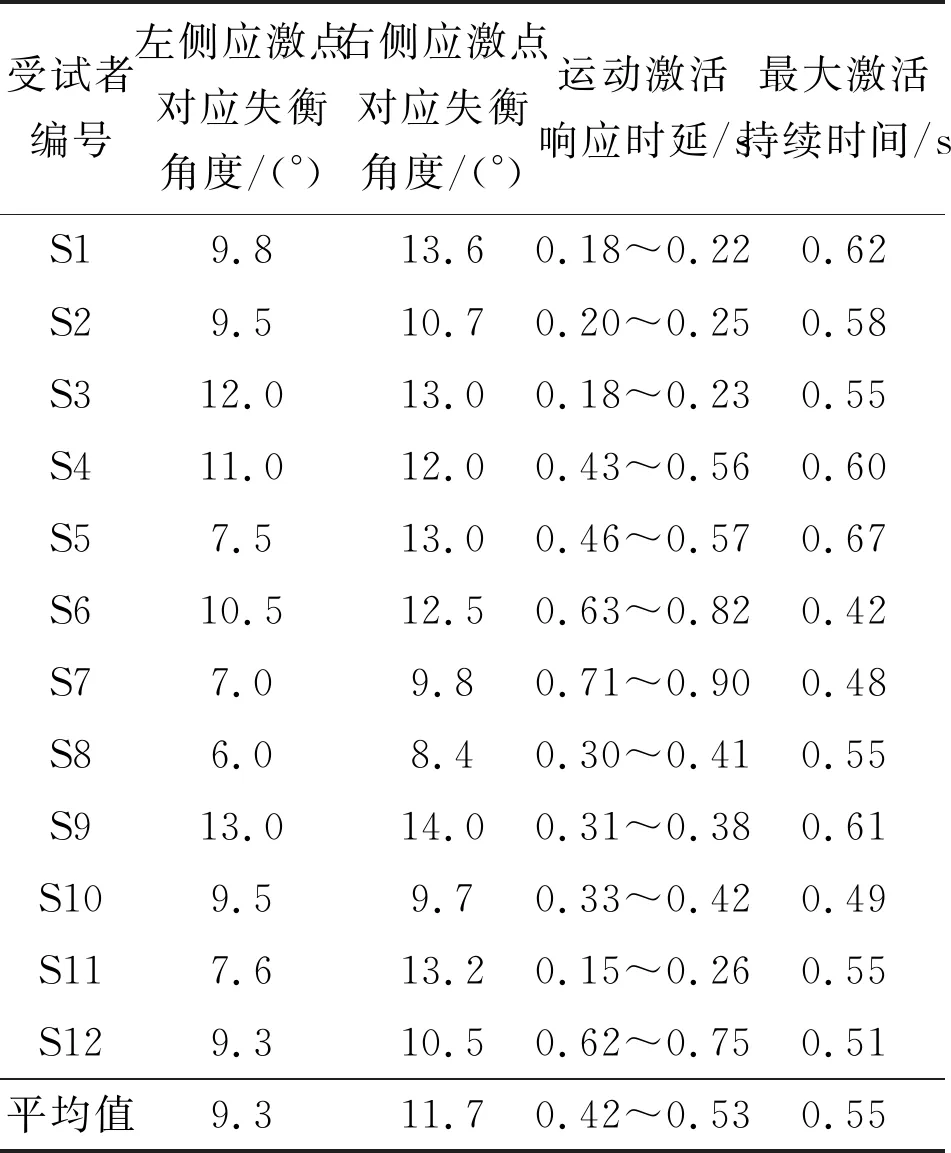
表1 应激刺激中枢响应参数
由统计结果可知,同一受试者的左右侧应激点都具有差异,表明了受试者在失衡刺激下左右侧运动响应水平具有一定的差别,且右侧身体平衡控制能力普遍优于左侧平衡的控制,这可能取决于个体的利手特性。虽然本实验未将利手因素作为受试者筛选的条件,但是由于人群中右利手的人数普遍大于左利手人数,所以受试者群体的右侧平衡控制能力优于左侧平衡控制能力,也符合右利手在人群中数量占优的规律。不同受试者自主意念诱发的响应时延规律具有差异性,即感知与反馈响应的时延为0.15~0.90 s不等,但最大激活响应时间的持续时间具有一致性,为400~600 ms。不难理解,由于受到个体差异的影响,不同受试者对于应激的响应水平与脑-机交互的操作能力都具有一定的差异性,这也决定了外部“神经搭桥”感知-激活的延迟特异性。不过,有效的激活响应量化结果可以指导脑-机接口外源刺激、系统响应,辅助训练参数的更新优化,以达到诱发可控、时序同步的交互效果。
4.4 应激触发激活响应同步增强效果验证
为了验证应激触发点诱发对于闭环调控同步增强的效果,本小节分析对比应激触发运动诱发与无应激运动想象实验结果。实验通过有应激和无应激的外界刺激诱发受试者产生中枢的激活响应,利用不同诱发模式下感知反馈激活响应时延,验证应激触发的同步增强效果,分析结果如图9所示。

图9 不同诱发模式下感知反馈激活响应时延Fig.9 The delay for motor perception and feedback activation in different evoked pattern
由分析结果可知,所有受试者在应激触发点后的1 s内都产生了运动激活响应,12名受试者的感知反馈平均时延为0.629 8 s,而传统的运动想象平均响应时间长达1.782 9 s。除受试者S6以外,两种范式对于其他所有受试者的响应都具有显著性差异(p<0.01),且应激刺激的诱发响应同步性在所有受试者之间都高于传统的运动想象,由此证明了应激刺激对于运动激活响应的有序性调控具有积极的影响。受试者S6在应激诱发任务中产生的效果没有优于传统想象,可能的原因是由于受试者尚未在训练过程中适应失衡应激的刺激模式,进而导致其无法利用应激刺激完成主观运动意念的激活。也可能是在失衡刺激中产生了身体运动,进而给脑电信号带来了较大的串扰。
图9表明,应激运动诱发相较于传统的运动想象具有更强的诱发效果,结合上文中提到应激刺激的作用效果,其可能原因是:在重心失衡应激刺激下,受试者的自主神经运动控制会对重心失衡的状态产生应激反应,进而导致其前庭神经以及负责运动感知和决策的前额叶在应激刺激过程中产生激活,而伴随着受试者主观运动意图的响应,运动皮层随之激活,在前额叶的激活前提下,该运动激活效果可能具有叠加效果,进而产生诱发增强的效果。
5 结 论
本文面向运动诱发脑-机交互过程中运动感知环路与反馈环路激活的时序规律展开研究,通过应激运动诱发脑-机接口平台,开展重心失衡任务下的运动诱发激活时序检测方法,研究基于脑功能网络的激活时序量化规律,揭示了应激刺激脑-机交互过程中,运动感知激活与控制激活的时序规律和脑网络动态演化过程。
脑功能网络分析结果表明,应激刺激运动诱发的网络动态演化规律与传统运动激活响应的演化规律具有一致性,且在达到应激触发点后,运动中枢的网络连接密度和信息交流会变得更加密集,其动力学特性表征的脑激活响应更加强烈。通过检测应激触发点与脑电激活响应点,可以实现脑-机交互任务中,感知神经与运动控制神经激活时延的有效量化。在线实验结果表明,负责运动控制的脑电特征一般会出现于运动感知激活后的0.15~0.90 s内,具体时延在不同受试者之间具有差异性,而其最大激活响应的持续时间为400~600 ms,在不同受试者之间具有一致性。同时,基于应激触发的运动意念诱发模型,可以有效提升感知环路与控制环路神经激活的同步性,且该同步性的增强效果具有显著性。
