Ccr4-Not复合物的生物学功能研究进展
2023-10-28温贵兰
吴 洁, 温贵兰, 赵 静, 胡 璇
(贵州大学动物科学学院,贵州 贵阳 550025)
Ccr4-Not(碳分解阻遏蛋白4复合体)是存在于真核生物中的1种独特、必需、保守的多亚基蛋白复合物,其在真核生物中由核心亚基Not1、Not2、Not4、Not5、Caf1、Ccr4、Caf40组成[1],整体形成1个扁平的L形结构[2]。是P-Body(mRNA processing body)的成分之一,起到去poly(A)尾的作用[3],具有参与转录延伸、mRNA衰变、脱腺苷酸化、蛋白质泛素化等功能[4],并在一些病毒(甲型流感病毒、人类免疫缺陷病毒、猪流行性腹泻病毒等)和疾病(乳腺癌、非小细胞肺癌、骨肉瘤等)中发挥作用。
1 Ccr4-Not复合物的组成
Ccr4-Not在不同生物体中的组成有所不同,某些亚基是不同真核生物共有的,可以认为是核心亚基,包括Ccr4(碳分解代谢物阻遏)、Caf蛋白(Ccr4相关因子)、Caf1、Caf40和Not蛋白(Not1、Not2、Not4、Not5)[1](见图1)。在酵母中除了核心亚基,还包括亚基Caf130和Not3[4]。在黑腹果蝇和人类中还发现了亚基CNOT10和CNOT11[5]。另外,在不同物种中各个亚基的名称也有所不同(见表1)。各亚基分子量为:Not1(237 kDa)、Not2(34 kDa)、Not3(73 kDa)、Not4(54 kDa)、Not5(66 kDa)、Ccr4(76 kDa)、Caf1(37.5 kDa)、Caf40(32 kDa)[6]。

表1 Ccr4-Not复合物亚基在酿酒酵母、人类、黑腹果蝇中的名称和功能[7]
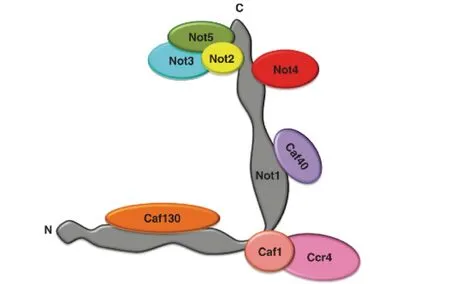
图1 Ccr4-Not复合物的组成和平面结构[6]
2 Ccr4-Not复合物的结构
目前该复合物在粟酒裂殖酵母、酿酒酵母、黑腹果蝇、人类4个物种中的结构已获得详细信息[7]。
2.1 整体构架
Ccr4-Not复合物在低分辨率电子显微镜下显示为1个扁平的L形结构(见图2),具有2个长度相似的臂(180Å和190Å)[2]。
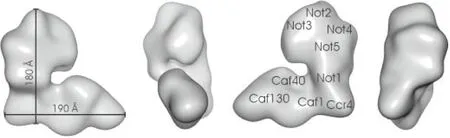
图2 Ccr4-Not复合物的三维结构[2]
2.2 各亚基的结构域和对接位点
在上述4个物种中,该复合物的亚基至少组成3个模块,包括脱腺苷酶模块(Ccr4和Caf1)、E3连接酶模块(Not4)、Not模块(Not2和Not5)[7]。其中最大的亚基Not1形成支架连接其他成分,其C端区域形成1个刚性结构域,由2个垂直堆叠的HEAT样重复序列组成;Not10和Not11结合在Not1的N端结构域上;Ccr4包含EEP(核酸内切酶-核酸外切酶-磷酸酶)和LLR(富含亮氨酸重复序列)结构域,Caf1包含DEDD结构域,Ccr4和Caf1之间相互作用并结合在Not1的MIF4G结构域上;Caf40结合在Not1中央的DUF3819螺旋结构域上;Not4包含N端的RING结构域、中央的RRM(RNA识别基序)结构域和预测为非结构化的C端结构域,并通过C端结构域结合在Not1上[1];Not2和Not5各自包含Not-box结构域并形成异二聚体结合在Not1的C端结构域上[8](见图3)。
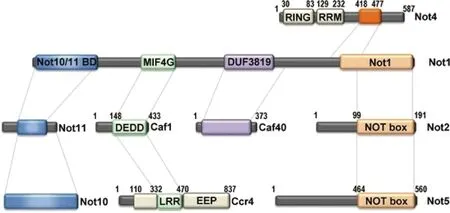
图3 Ccr4-Not复合物各亚基的结构域和对接位点[1]
3 Ccr4-Not复合物的功能
最初该复合物被发现在转录中发挥作用[9],随后在基因调控等方面的功能也逐渐被发现。这些功能可能由复合物的多个亚基提供,也可能由其中的某个亚基提供。
3.1 在基因调控中的功能
Ccr4-Not在基因调控中有广泛的作用,例如转录延伸、mRNA衰变、脱腺苷酸化和蛋白质泛素化等[4]。参与mRNA代谢的不同方面,逐渐成为真核生物中基因表达稳态的重要调节因子[10]。(1)在转录中的功能:Ccr4-Not通过一般转录因子TFIID控制转录起始,并与延伸因子TFIIS协同工作重新激活停滞的RNA聚合酶Ⅱ(RNAPⅡ)[11],从而直接影响转录延伸。此外,在翻译过程中也发挥着作用。Villanyi Z等[12]通过酵母进行2个杂交和免疫共沉淀实验,并通过核糖体分级分离和荧光显微镜检查,表明Not5与Rpb4相互作用将翻译和转录相互联系起来。(2)参与mRNA衰变:mRNA衰变是细胞内维持mRNA代谢稳态的重要机制之一,能够控制基因表达的强度和时机,这个过程由一系列酶参与完成。Ccr4-Not可作为酶参与不同类型的mRNA衰变,通用mRNA脱腺苷酸化、无义介导的mRNA衰变(NMD)、靶向mRNA衰变和脱腺苷化独立脱帽[7]。(3)脱腺苷酸化功能:mRNA的3’ poly(A)尾长短的变化会影响mRNA稳定性和翻译能力,Ccr4和Caf1作为脱腺苷酶可以促进poly(A)尾的缩短[13]。(4)蛋白质泛素化功能:蛋白质泛素化需要一系列特殊的酶发挥作用,包括泛素激活酶(E1)、结合酶(E2)和连接酶(E3)等。CNOT4可以作为E3连接酶发挥作用,并且这个功能存在于其N端的RING结构域中[14]。
3.2 各亚基的功能
(1)CNOT1的功能:其在复合物中主要作为支架连接其他成分,以及许多其他功能。如:充当RNA结合蛋白的平台[15],参与真核生物的mRNA衰变,调节主要组织相容性复合体Ⅱ类(MHCⅡ)基因的表达[16]。另外,在转录水平上介导转录沉默[17]。(2)CNOT2和CNOT3的功能:在结构上相似,有许多相似的功能。如:在各种生理过程中调节mRNA衰变,调节Ccr4-Not的酶活性并促进靶mRNA的降解[15],调节主要组织相容性复合体Ⅱ类(MHCⅡ)基因的表达[16]。(3)Ccr4的功能:是复合物的脱腺苷酶之一,涉及多种生物学功能。如:在秀丽隐杆线虫中,Ccr4的耗尽会使mRNA的poly(A)尾长增加[13]。人类中的CNOT6L具有脱腺苷酶活性,能够选择性地与poly(A)残基相互作用[18]。(4)Caf1的功能:作为复合物的另一个脱腺苷酶,最初在酵母中被确定为所需的转录调节因子[19]。随后,在布氏锥虫中发现Caf1具有脱腺苷酶活性,这对许多mRNA脱腺苷酸化和随后的降解至关重要[20]。另外,CNOT7和CNOT8参与调节多能干细胞(iPSC)的生成[21]。(5)CNOT9的功能:主要作为转录辅助因子发挥作用,并包括许多其他的功能,例如:在哺乳动物组织(尤其是造血系统)的生长和分化过程中发挥作用[15],参与脱腺苷酸化和mRNA衰变[22]。(6)CNOT10和CNOT11的功能:是人类和果蝇的特殊亚基,其本身似乎没有酶活性,但可以稳定RNA底物以刺激脱腺苷酸化[7]。(7)CNOT4的功能:在复合物中充当泛素蛋白连接酶(E3)发挥作用[14],还参与雄性生殖细胞减数分裂过程中的mRNA降解,以及有效的DNA损伤修复和XY染色体交叉[23]。
4 Ccr4-Not复合物与病毒的相关研究
近年来研究发现,该复合物在甲型流感病毒(AIV)、登革热病毒(DENV)、人类免疫缺陷病毒(HIV)、猪流行性腹泻病毒(PEDV)等病毒中也发挥着不同程度的作用。Lin Y C等[24]通过共免疫沉淀测定、体外泛素化测定、串联质谱分析等方法,发现细胞泛素连接酶CNOT4能使AIV核蛋白(NP)泛素化,而NP的泛素化会增强病毒RNA的复制,因此CNOT4可介导AIV核蛋白泛素化促进病毒RNA复制。Liu J等[25]使用RNAi方法在DENV感染的A549、Huh7细胞中筛选Ccr4-Not的亚基,发现CNOT2在DENV感染中起着前病毒作用,CNOT2的下调减少了DENV RNA复制和蛋白质合成,表明CNOT2是Janus激酶信号转导和转录激活因子(JAK-STAT)通路的新型负调节因子,能够支持DENV感染。Matkovic R等[26]通过酵母双杂交(Y2H)筛选,确定CNOT1是人类沉默中枢(HUSH)复合物核心亚基(TASOR)的合作伙伴,并表明TASOR和CNOT1协同作用抑制HIV的表达。Wang J等[27]以兔抗PEDV S1多克隆抗体为诱饵,识别感染或未感染PEDV的猪小肠上皮细胞系(IPEC-J2),使用液相色谱串联质谱(LC-MS-MS)进行检测,表明CNOT2与PEDV S1蛋白相互作用并参与病毒感染,CNOT2的敲除能抑制PEDV的体外复制。
5 Ccr4-Not复合物与疾病的相关研究
随着对该复合物的不断研究和探索,许多研究发现其在乳腺癌、非小细胞肺癌、骨肉瘤等疾病中发挥作用。CNOT2通过调节乳腺癌细胞中的血管内皮生长因子信号,在转移、增殖和血管生成中发挥重要作用,可以作为乳腺癌治疗的有效分子靶点[28];miR-126-5p可能作为肿瘤抑制因子,通过直接结合CNOT7来抑制乳腺癌MCF7细胞系的迁移[29]。非小细胞肺癌CNOT2通过调节转录激活因子3(STAT3)信号通路促进细胞中TNF相关凋亡诱导配体(TRAIL)的敏感性,并且其耗竭和TRAIL的联合治疗可能是克服TRAIL耐药性的有效策略[30]。Zhang B等[31]运用免疫组织化学染色和蛋白质印迹等方法,指出CNOT4通过抑制肿瘤细胞功能(增殖、集落形成、细胞迁移、侵袭)和增加对CTL的敏感性,作为非小细胞肺癌的潜在肿瘤抑制因子发挥作用。Almasmoum H A等[32]发现,非霍奇金淋巴瘤中基因B细胞易位基因1(BTG1)的突变可能通过干扰其抗增殖活性的方式及与CNOT7和CNOT8相互作用的方式促进恶性转化和肿瘤细胞增殖。Wei K等[33]发现甲基化识别蛋白YTH N6-甲基腺苷RNA结合蛋白1(YTHDF1)通过调节CNOT7的m6A水平调控骨肉瘤细胞的增殖、迁移和侵袭。Vicente C等[34]使用不同的黑腹果蝇眼癌模型研究Not3和Ccr4-Not中其他亚基的潜在肿瘤抑制功能,数据显示亚基Not3、Not1、Not2和Pop2的表达减少都会导致肿瘤形成增加,这些数据共同确立了Ccr4-Not可作为肿瘤抑制因子发挥关键作用。
6 展望
近年来对Ccr4-Not复合物的整体组成和结构研究已取得较大进展,但各个亚基的具体结构仍然需要进一步探索。从现有的研究来看,该复合物在部分病毒和疾病中发挥了作用,但作用机制还需深入研究。另外,其与动物病毒相关的研究报道较少,需要进一步拓展这方面的研究。
