草甸草原关键物种功能性状对长期放牧和停牧恢复的响应
2023-10-23司雨凡李辉李子好姜翠霞郭昊南杨培志席杰军闫瑞瑞乌仁其其格山丹辛晓平
司雨凡,李辉,李子好,姜翠霞,郭昊南,杨培志,席杰军,闫瑞瑞,乌仁其其格,山丹,辛晓平
草甸草原关键物种功能性状对长期放牧和停牧恢复的响应
司雨凡1,2,李辉2,3,李子好2,3,姜翠霞2,3,郭昊南2,3,杨培志1,席杰军1,闫瑞瑞2,乌仁其其格4,山丹4,辛晓平
1西北农林科技大学草业与草原学院,陕西杨凌 712100;2呼伦贝尔草原生态系统国家野外科学观测研究站/中国农业科学院农业资源与农业区划研究所,北京 100081;3兰州大学生态学院,兰州 734000;4呼伦贝尔学院农林学院,内蒙古呼伦贝尔 021008
【目的】植物功能性状对环境变化及外部干扰的响应,反映了植物的表型塑性、生存策略及适应能力。通过长期放牧条件下草原植物功能性状的变化及停止放牧后的恢复响应研究,为草原恢复管理的研究提供理论依据。【方法】探究了生长季不同放牧梯度下呼伦贝尔草甸草原6个关键物种(羊草、日阴菅、糙隐子草、斜茎黄芪、裂叶蒿、细叶白头翁)叶片功能性状(叶片干重、叶片含水量、叶面积、比叶面积、叶片氮含量、叶片磷含量、叶片C/N、叶片N/P)的变化,以及生长季停止放牧后的草甸草原关键物种叶片功能性状的即时恢复响应。【结果】(1)6个关键物种的相对生物量和重要值随放牧强度增加而减小,轻度和重度放牧强度停止放牧后的值大于继续放牧。(2)6个关键物种的叶片干重、叶片C/N整体随放牧强度的增加呈减小趋势;叶片含水量、比叶面积、叶片氮含量整体随放牧强度的增加呈增大趋势;除裂叶蒿外各物种叶面积都随放牧强度增加而减小;叶片磷含量除糙引子草外都随放牧强度增加呈增大趋势;随放牧强度增加,羊草、日阴菅、糙隐子草、细叶白头翁的叶片N/P整体呈增大趋势,而斜茎黄芪、裂叶蒿呈减小趋势。(3)停止放牧后,重度放牧的恢复程度高于轻度放牧,6个关键物种的功能性状整体恢复较好,叶片干重、叶片含水量、叶面积、比叶面积、叶片N/P都表现为正响应、叶片磷含量呈现负响应,叶片氮含量、叶片C/N在6个关键物种中正负响应不一,叶片氮含量的正响应少于负响应,叶片C/N的正响应多于负响应。(4)禾草类的叶片形态学性状、杂类草的养分特征对停止放牧后的恢复更敏感,且叶片干重、叶面积和叶片磷含量对放牧强度和物种的交互作用的响应更为显著。停止放牧后,随6个关键种叶面积、叶片氮含量、叶片N/P的增加,叶片干重也在增加。【结论】长期放牧导致草原植物个体小型化,停止放牧后植物功能性状正响应多于负响应,草原植被功能出现明显的恢复响应,草甸草原的地上生物量有所增加。
长期放牧;停止放牧;叶片功能性状;恢复力;草甸草原
0 引言
【研究意义】草地是面积最大的陆地生态系统之一[1],不仅是巨大的陆地碳库、绿色生态屏障,也是草食畜牧业生产基地[2]。呼伦贝尔草原是我国草甸草原保存最完整的地区,也是欧亚大陆东段生物多样性起源中心之一[3],其利用强度比较大。高强度放牧利用造成草原普遍退化,限制了草原畜牧业发展和生态文明建设。长期以来,不同干扰条件下草原退化与恢复机制一直是我国草地生态学研究的重点和热点[4]。放牧压力下的草地退化研究早期强调群落学机制,包括演替理论、草地生产力及生物多样性的干扰响应等,随着植被采样及分析方法的进展,不同放牧压力下草地生态系统多功能性和关键过程的响应,以及器官尺度、个体、种群和群落的功能性状及其多样性水平,成为更好地理解草地放牧系统、解释草原放牧退化与恢复机制的关键[5]。草原生态系统恢复力目前还无法用定量化方法直接测定,但可以通过植被土壤特征与原初状态的距离来进行判断,结合植物器官形态、数量特征和生理养分等功能性状的干扰响应及干扰去除后的恢复响应,可以判断植物个体功能对外部环境的适应性和自我调节能力,为植物功能性状、干扰响应和生态系统功能之间关系研究提供了新的思路[6-8]。因此,植物功能性状在不同放牧压力下及解除放牧压力后的恢复响应,对于维持草地放牧生态系统的多功能及其稳定性、保护草原生物多样性、提高草原生产力、实现草原放牧系统可持续发展具有十分重要的理论和实践意义。【前人研究进展】WRIGHT等[9]首次将叶经济谱概念带入,标准量化了叶片性状和叶片性状间的关系,国内外生态学关于植物功能性状的研究主要集中在叶片性状、根系性状、功能多样性及其与生态系统稳定性之间的关系等领域[10-12],已经从某单一性状变化、多性状组合变化及其之间关系的探讨,发展到基于植物功能性状解释生态系统过程、功能与环境变化的研究[13-14]。有大量研究表明,叶片性状与地上生产力和资源合理利用极其相关,作为在温性草甸、草甸草原到典型草原的关键建群物种,羊草的茎叶性状及其对放牧的响应研究是植物功能性状研究的重点之一,现有研究表明放牧条件下羊草出现了个体矮小化特征[15]。同时叶片的生理和生化特征随环境变化产生了不同的变化,这表明植物可以通过不同功能性状的权衡、通过表型可塑性达到对异质生境的适应[16],例如羊草矮化型变、糙隐子草对于不同放牧强度干扰表现出矮小-密枝化就是对高强度放牧的避牧适应对策[15-19]。邢小青等发现放牧条件下糙隐子草表现为高度降低、叶片数量和叶面积增加,而克氏针茅表现为茎叶高度和比叶面积减小,不同物种的不同功能性状的放牧响应不一致,是其对干扰产生的不同的生态对策[20-21]。草地的不同利用方式会使植物个体的功能性状产生不同变化和策略,以内蒙古典型草原优势物种为对象,研究发现优势物种通过逃避策略来适应放牧干扰,在无干扰下植物碳含量增加,降低干扰后,优势物种改变自身性状从资源获取型向存储型转变[22]。植物群落对干扰的抵抗力和适应能力主要取决于优势物种的植物功能性状,不同长期放牧强度下优势物种的地上生物量、单叶面积是对放牧响应的敏感性状,而叶干物质含量、比叶面积和叶片氮含量则是惰性性状[23]。养分特征对不同放牧强度的响应研究表明,重度放牧降低了C/N,轻度放牧影响较小,随放牧变化N和P产生的响应也不同[24-26]。石明明等在对围封和放牧下高寒草甸植物性状研究发现,不同植物种对放牧的响应模式存在差异,是由于放牧影响物种共存依赖于对多种资源的竞争包括对光资源和土壤养分的竞争[27]。【本研究切入点】植物功能性状及不同功能性状之间关系、功能性状与环境关系的研究是当前植物生态学研究的热点问题之一。草原植物功能性状研究主要集中于不同干扰条件下的植物功能性状及其响应机制和停牧后恢复原始状态的时间。但是,不同放牧强度对植物功能性状的影响程度如何?放牧压力下尤其是重度放牧条件下,放牧解除后植物功能性状能否有效恢复?不同放牧强度下的即时恢复响应如何?目前尚无此类探讨植物及群落功能在不同放牧压力下的恢复力和即时响应的研究报道。本研究遴选草甸草原关键物种、不同叶片形态和生理生化性状,研究长期不同放牧强度下的植物功能性状变化以及放牧解除后植物功能性状的即时恢复响应,探索放牧演替过程中植物个体和群落的适应机制与策略。【拟解决的关键问题】(1)不同放牧强度如何影响关键物种的功能性状?(2)放牧解除后,不同关键物种的功能性状如何响应?(3)不同放牧强度下,哪个关键物种或功能性状恢复能力更强?通过上述研究,旨在为草原合理放牧与恢复管理提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于内蒙古自治区呼伦贝尔市,依托于内蒙古呼伦贝尔草原生态系统国家野外观测研究站长期控制放牧实验平台(49°19′—49°20′N,119°56′—119°57′E),处于大兴安岭西麓丘陵向蒙古高原的过渡区羊草草甸草原的核心地带,海拔670 m左右,属温带半干旱大陆性气候,年平均降水量350—400 mm,年均气温-3—-1 ℃,≥10 ℃年积温为1 580—1 800 ℃,无霜期110d左右,降水期多集中在7—9月[28]。植被类型为羊草+杂类草草甸草原,优势物种有羊草()、贝加尔针茅()、主要物种有糙隐子草()、日荫菅()、裂叶蒿()、伴生种有斜茎黄芪()等。土壤类型为黑钙土或栗钙土。
1.2 研究方法
1.2.1 样地设置 试验选择地形较为平坦,土壤和植被状况较一致的草地,面积为90 hm2,用围栏围成18个等面积小区,各小区面积为5 hm2,根据家畜日食量以及羊草草甸草原牧草利用率情况,当地常年平均载畜率为0.46 AUE/hm2(AUE为家畜单位当量,等于一头500 kg的成年牛)。因此,试验围绕平均载畜率设置了6个放牧强度,即CK(对照,不放牧)、0.23(轻度)、0.34(轻中度)、0.46(中度)、0.69(重度)、0.92(极重度)AUE/hm2,分别记为G0、G0.23、G0.34、G0.46、G0.69、G0.92。按随机区组设计,每个放牧强度设置3个重复不同放牧强度的小区内,分别放牧250—300 kg的育成母牛0、2、3、4、6、8 头,试验用牛组间差异不显著。放牧试验始于2009年,每年生长季6月1日至9月30日连续放牧无补饲,10月开始结束放牧。2021年5月,在每个小区内选取并围封1个20 m×30 m的区域,做停牧恢复处理,如图1所示。
基于多年群落调查数据,选择6个关键物种为对象开展植物功能性状研究:羊草、糙隐子草、日阴菅、斜茎黄芪、裂叶蒿、细叶白头翁。这些物种分属不同功能群,生物量及重要值占对照群落总量的70%以上、不同放牧强度群落总量的40%—70%。

图1 不同放牧强度试验平面图
1.2.2 植物群落调查 2021年8月上旬,地上生物量达到最大时,每个放牧小区和停牧小区采用系统取样法,选取5个50 cm×50 cm样方进行地上群落调查,记录样方内植物种类、高度、多度、盖度,并将各物种地上绿色部分齐地面刈割、称量鲜重,带回实验室105 ℃杀青20 min后65 ℃烘箱烘至恒重并称重。计算每个物种的重要值(IV),公式如下:
=(相对盖度+相对多度+相对地上生物量)/3×100
式中,相对盖度=某一种植物的分盖度/全部植物分盖度的总和;相对多度=某一种植物的多度/全部植物密度总和;相对地上生物量=某一种植物的地上生物量/全部植物地上生物量总和。
1.2.3 植物功能性状测量 2021年8月中下旬,每个放牧小区和停牧小区内,随机选取6个关键物种各10—30株,装在密封袋内,放在装有冰袋的冷藏箱内尽快带回实验室。根据叶片大小,选取10—60片完整无病害的叶片,测量平均叶或叶片的单侧投影表面积(LI-3000C,Li-Cor,Lincoln,NE,USA);叶片样品用万分之一天平称量鲜重,之后105℃杀青20 min后65 ℃烘箱烘至恒重并称重。叶片含水量=(叶片鲜重-叶片干重)/叶片鲜重×100%;比叶面积等于鲜叶单侧面积除以其干物质量。
称重后,将叶片样品充分磨碎后,分别利用重铬酸钾-浓硫酸氧化法、凯氏定氮法、钼锑抗吸光光度法,测定叶片有机碳、全氮、全磷,计算C/N、N/P。
1.2.4 停止放牧后恢复响应的定量评估 本文利用停止放牧后各功能性状的响应程度(response ratio)来定量评估解除放牧后各功能性状的恢复程度,公式如下:
式中:VSG表示停止放牧后各物种的性状值;VG表示长期放牧各物种的性状值。RR值为正,是正响应,代表停止放牧草地的恢复生长>长期放牧草地的自然生长;RR值为负,是负响应,代表长期放牧草地的自然生长>停止放牧草地的恢复生长。
1.3 数据处理与分析
采用 Excel 2019 、SPSS 21.0 、R 4.1.3软件进行数据处理、统计分析和制图。采用单因素方差分析(one-way ANOVA),揭示不同放牧强度下的叶片干重、叶片含水量、叶面积、比叶面积、叶片氮含量、叶片磷含量、叶片C/N、叶片N/P的变化,利用 Duncan 法进行平均值之间的多重比较;利用双因素分析比较和分析放牧强度和物种差异对植物功能性状的影响;对不同放牧强度下和停止放牧后的6个关键物种的8个功能数据利用R绘制1﹕1线图,并利用独立样本T检验进行放牧和停牧的差异分析,不符合正态分布用Mann-Whitney U检验。显著性水平设为<0.05,极显著性水平设为<0.01。
2 结果
2.1 关键物种对不同放牧强度及停止放牧的响应
6个关键物种的相对生物量随着不同的放牧强度整体呈减小趋势。对照组G0.00有最大相对生物量(73.65%),极重度放牧G0.92相对生物量最小(41.37%,图2-a);重要值整体随着不同的放牧强度而减小。对照组G0.00的重要值最大(62.74%),极重度放牧G0.92重要值最小(37.04%,图2-b)。停止放牧后,重度放牧强度(G0.69)样地6个关键物种的相对生物量较放牧增长最多,达到11.93%,但是没有改变总体相对生物量随放牧强度减小的整体趋势(图2-a);重度放牧强度(G0.69)样地6个关键物种的重要值较放牧增长最多,达到6.34%,但没有改变总体相对生物量随放牧强度减小的整体趋势(图2-b)。各物种相对生物量和重要值的停牧恢复的整体变化趋势与继续放牧下基本一致,并轻度和重度停牧大于放牧。
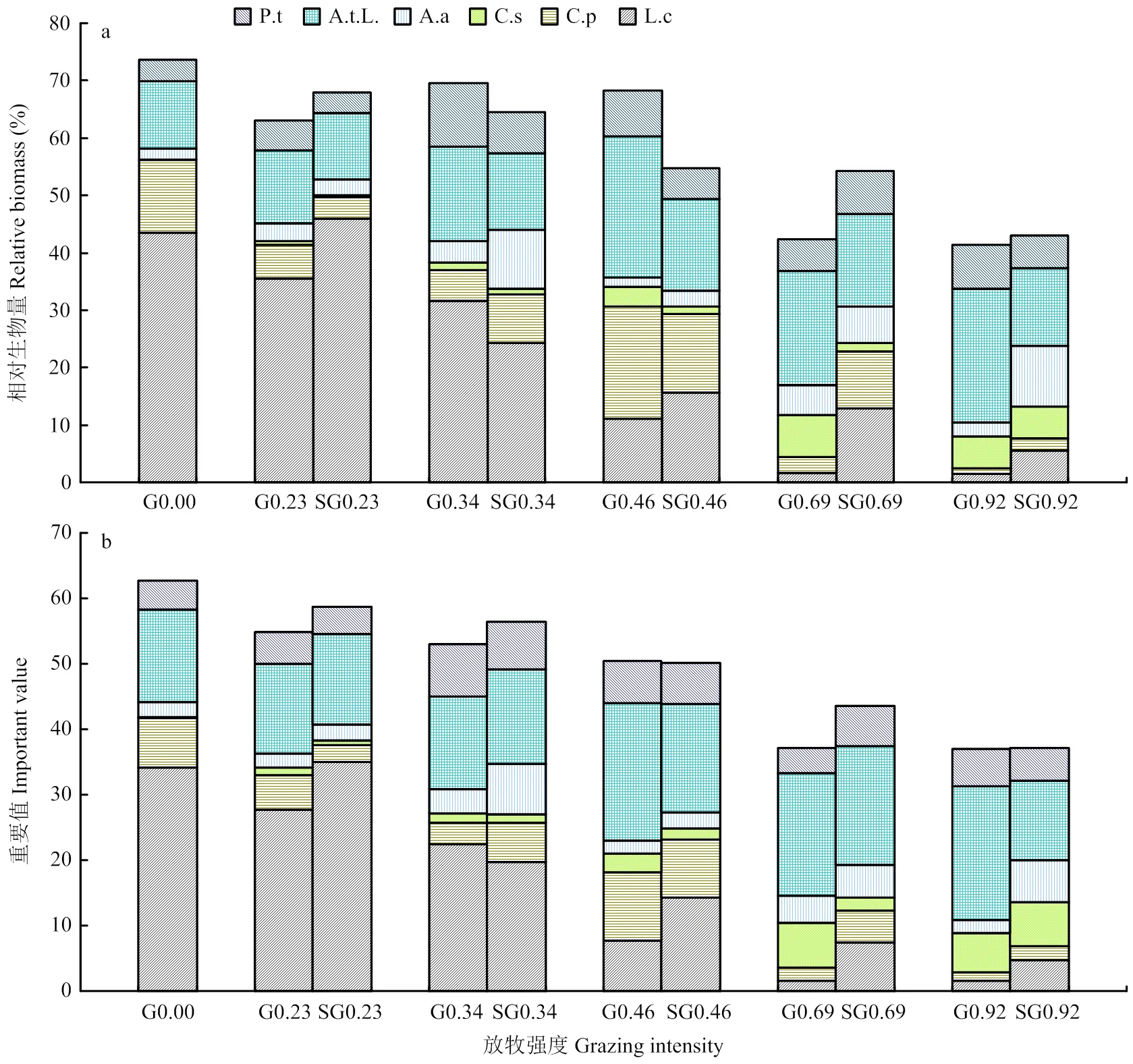
a,b分别是6个关键物种不同放牧强度下停止放牧后的相对生物量(a)、重要值(b),G代表继续放牧,SG代表停牧恢复,P.t、A.t.L.、A.a、C.s、C.p、L.c分别代表细叶白头翁、裂叶蒿、斜茎黄芪、糙隐子草、日阴菅、羊草
2.2 不同放牧强度下关键物种叶片功能性状相对的变化
6个关键物种的叶片干重随放牧强度的增加整体呈减小趋势,羊草、日阴菅、糙隐子草、斜茎黄芪和裂叶蒿在不同放牧梯度下有显著差异(<0.05),细叶白头翁无明显差异(图3-a)。叶片含水量和比叶面积都随放牧强度整体都呈增大趋势,糙隐子草、裂叶蒿的叶片含水量在不同放牧梯度下有显著差异(<0.05),羊草、日阴菅、斜茎黄芪、细叶白头翁无明显差异(图3-b)。羊草、日阴菅、糙隐子草、细叶白头翁的比叶面积在不同放牧强度下有显著差异(<0.05),斜茎黄芪、裂叶蒿无明显差异(图3-d)。叶面积随放牧强度的增加变化不一,羊草、日阴菅、糙隐子草、斜茎黄芪、细叶白头翁随放牧强度增加整体呈减小趋势,裂叶蒿整体呈增大趋势,羊草、日阴菅、糙隐子草在不同放牧强度下有显著差异(<0.05),斜茎黄芪、裂叶蒿、细叶白头翁无明显差异(图3-c)。
6个关键物种的叶片氮含量随放牧强度的增加整体都呈增大趋势,羊草在不同放牧强度下有显著差异(<0.05),日阴菅、糙隐子草、斜茎黄芪、裂叶蒿、细叶白头翁无明显差异(图4-a)。叶片C/N随放牧强度的增加整体呈减小趋势,羊草在不同放牧强度下有显著差异(<0.05),日阴菅、糙隐子草、斜茎黄芪、裂叶蒿、细叶白头翁无明显差异(图4-c)。叶片磷含量和叶片N/P随放牧强度的增加表现出不一致的变化趋势,叶片磷含量在不同放牧强度下除羊草外都有显著差异(<0.05),羊草、日阴菅、斜茎黄芪、裂叶蒿、细叶白头翁的叶片磷含量都呈增大趋势,糙隐子草呈略减小趋势(图4-b)。羊草、日阴菅、糙隐子草、细叶白头翁的叶片N/P整体随放牧强度增大呈增大趋势,斜茎黄芪、裂叶蒿呈减小趋势,叶片N/P在不同放牧强度下无明显差异(图4-d)。
2.3 关键物种叶片功能性状对不同放牧强度停止放牧后的响应
6个关键物种的叶片形态学性状对不同放牧强度停止放牧后的响应如图5所示,各个物种的性状响应不一致。叶片干重、叶片含水量和叶面积的响应的整体趋势为正响应,羊草、糙隐子草在轻度、裂叶蒿在重度、细叶白头翁在轻中度、斜茎黄芪在G0.46和G0.92的叶片干重为负响应,其余均为正响应,细叶白头翁、裂叶蒿在不同放牧强度间有显著差异(<0.05)(图5-a)。日阴菅、糙隐子草、斜茎黄芪在G0.34、裂叶蒿在轻中和极重度、细叶白头翁在轻重度为负响应,其余均为正响应(图5-b)。羊草和细叶白头翁在轻中度、斜茎黄芪在中度和极重度、裂叶蒿在重度为负响应,其余均为正响应,羊草、糙隐子草、裂叶蒿、细叶白头翁在同放牧强度间有显著差异(<0.05)(图5-c)。斜茎黄芪的比叶面积在5个放牧强度停牧后都为正响应,羊草、裂叶蒿在中度和重度、日阴菅在中度、糙隐子草在轻中度、细叶白头翁在轻度为正响应,其余均为负响应(图5-d)。禾草类植物的形态学性状响应变化幅度高于杂类草。
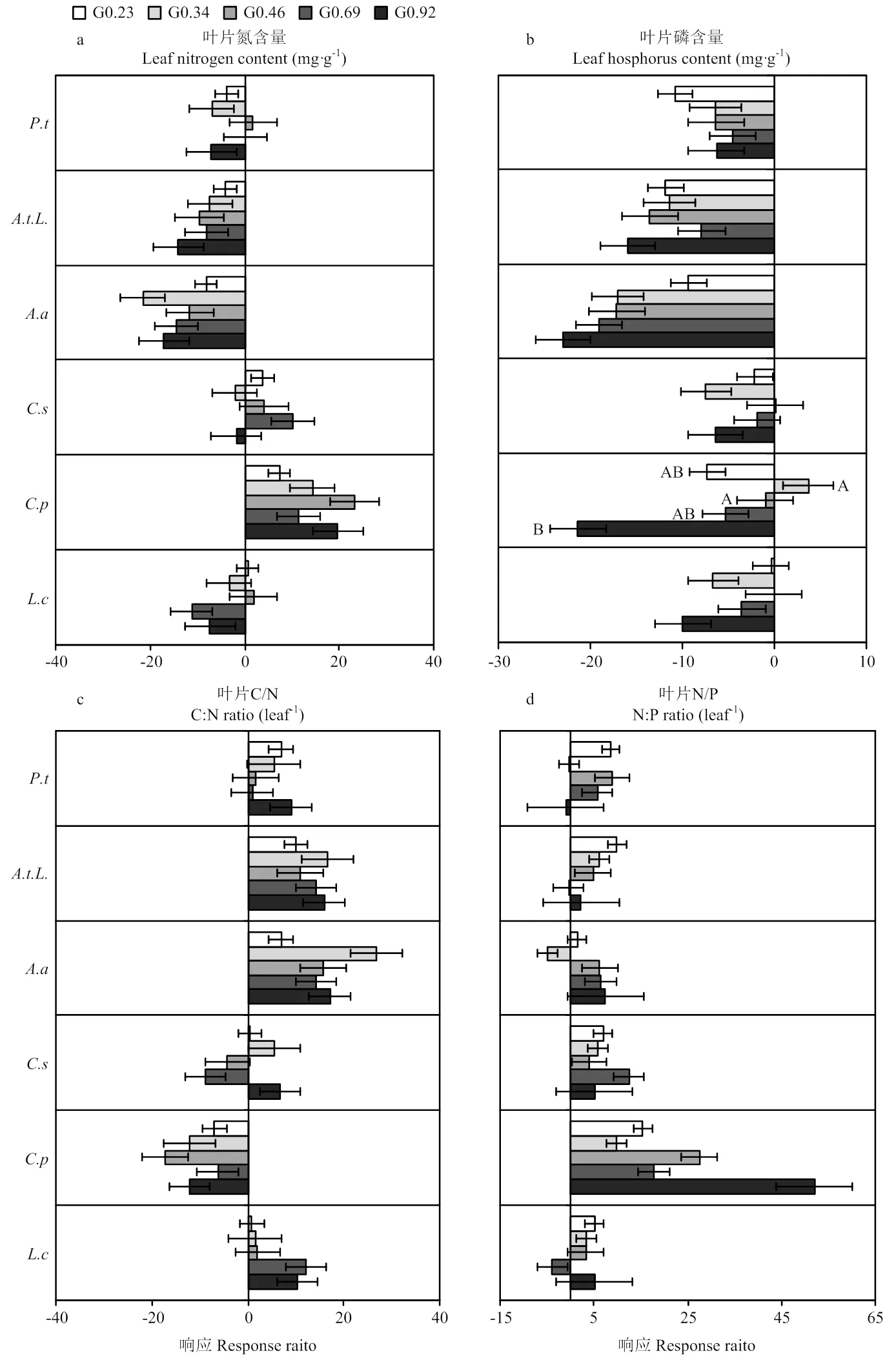
6个关键物种的养分特征对停止放牧后的响应如图6所示,各个物种的养分特征没有一致的响应。叶片氮含量、叶片磷含量的响应的整体趋势为负响应,日阴菅在5个放牧强度停牧后都为正响应,糙隐子草在轻、中、重度为正响应,羊草在轻中度、细叶白头翁在中重度为正响应,其余均为负响应(图6-a)。叶片磷含量除日阴菅、糙隐子草轻中度放牧为正响应,其余均为负响应,日阴菅在同放牧强度间有显著差异(<0.05)(图6-b)。叶片C/N、叶片N/P的响应整体趋势为正响应,日阴菅的叶片C/N在5个放牧强度下都为负响应,糙隐子草在中重度放牧为负响应,其余均为正响应(图6-c)。羊草、裂叶蒿的叶片N/P在重度放牧、细叶白头翁在G0.34、G0.92、斜茎黄芪在轻中度为负响应,其余都为正响应(图6-d)。杂类草植物的养分特征响应变化幅度高于禾草类。
2.4 不同的放牧强度及物种与叶片功能性状的关系
草甸草原不同放牧强度及不同物种对叶片功能性状的分析表明(表1),不同放牧强度对叶片功能性状影响不一,对叶片干重、叶片含水量、叶面积、比叶面积、叶片氮含量、叶片磷含量、叶片C/N有显著影响(<0.05),对叶片N/P无显著影响。不同物种的叶片干重、叶片含水量、叶面积、比叶面积、叶片氮含量、叶片磷含量、叶片C/N、叶片N/P都有极显著差异(<0.01)。放牧强度和物种的交互作用对叶片干重、叶面积、叶片磷含量有显著影响(<0.05)。停止放牧后,6个关键物种的叶片干重随叶面积(<0.01)、叶片氮含量(<0.01)和叶片N/P(=0.011)的增加而增加。6个关键物种的不同功能性状在放牧和停牧后的关系不一致,叶片磷含量在放牧和停牧后有显著差别(<0.05),叶片干重、叶面积杂类草植物停牧恢复大于继续放牧(图7-a, b),叶片氮含量、叶片磷含量杂类草植物继续放牧大于停牧恢复(图7-c, d),叶片C/N禾本科植物继续放牧大于停牧恢复,杂类草植物停牧恢复大于继续放牧(图7-e),叶片N/P无明显一致响应,斜茎黄芪继续放牧>停牧恢复,裂叶蒿和糙隐子草停牧恢复>继续放牧(图7-f)。

a,b,c,d分别是6个关键物种不同放牧强度下叶片干重、叶片含水量、叶面积、比叶面积的变化,L.c、C.p、C.s、A.a、A.t.L.、P.t分别代表羊草、日阴菅、糙隐子草、斜茎黄芪、裂叶蒿、细叶白头翁。图中不同大写字母代表各放牧强度间有显著差异
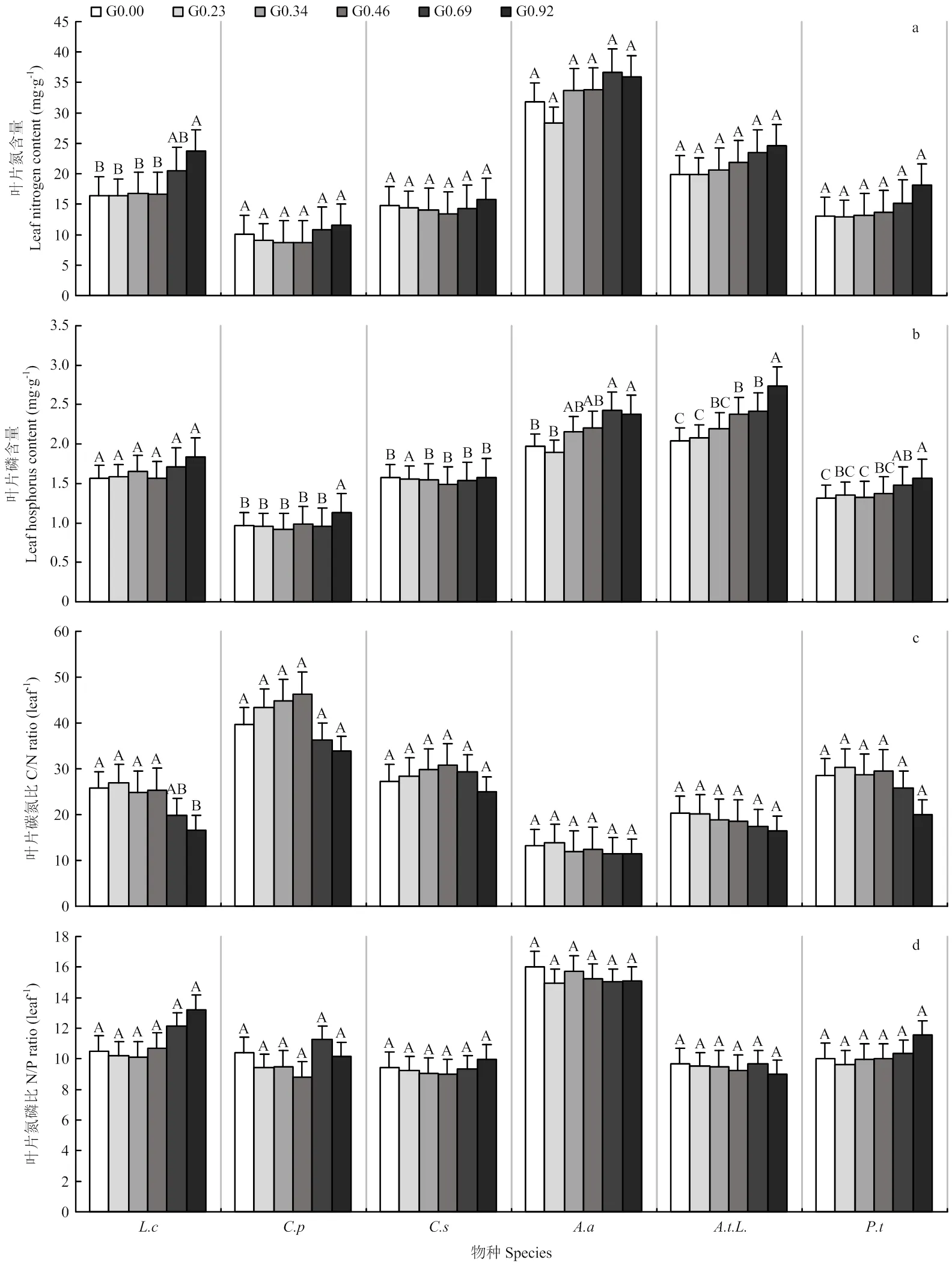
a,b,c,d分别是6个关键物种不同放牧强度下叶片氮含量、叶片磷含量、叶片C/N、叶片N/P的变化,L.c、C.p、C.s、A.a、A.t.L.、P.t分别代表羊草、日阴菅、糙隐子草、斜茎黄芪、裂叶蒿、细叶白头翁。图中不同大写字母代表各放牧强度间有显著差异
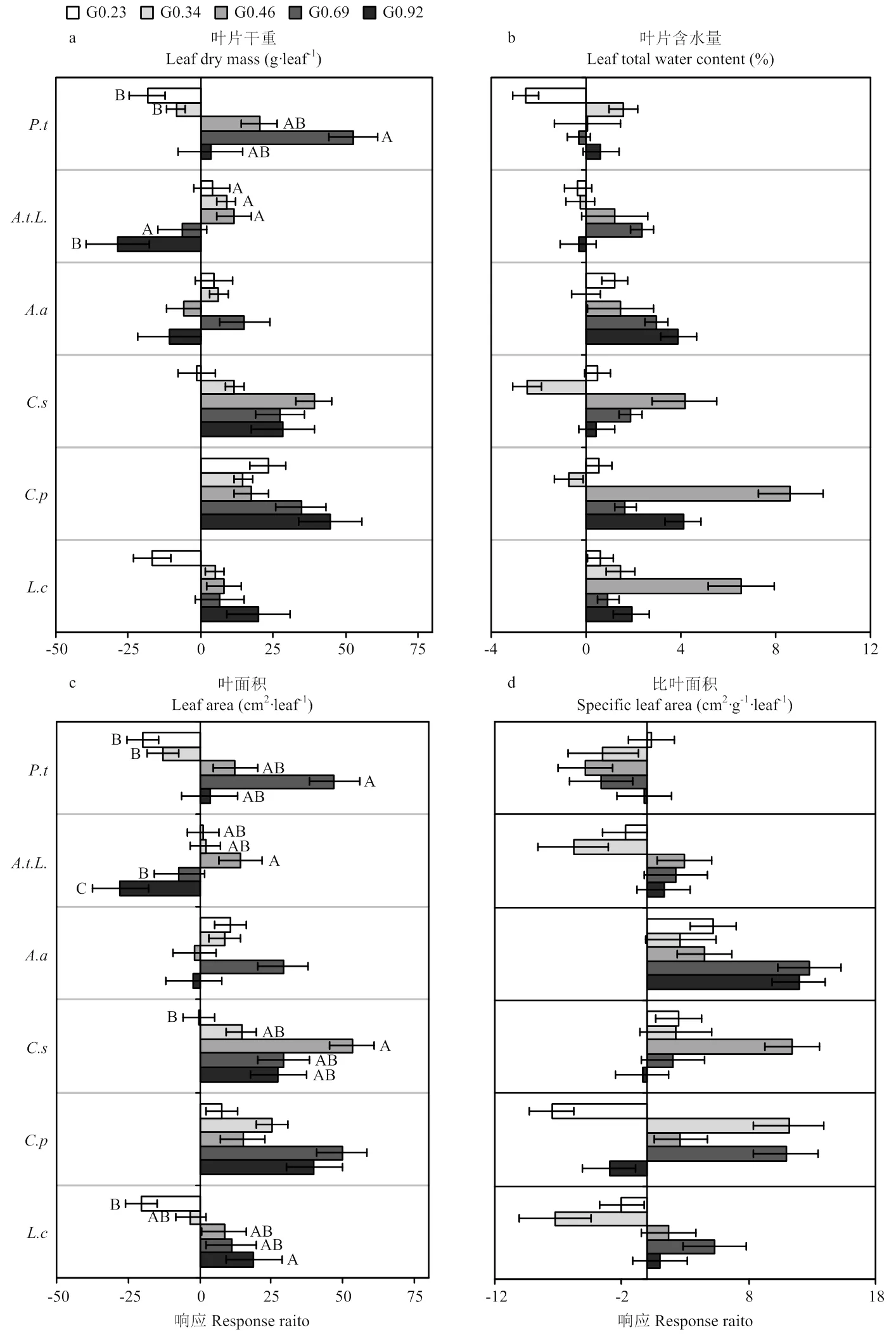
a,b,c,d分别是6个关键物种的叶片干重、叶片含水量、叶面积和比叶面积对停止放牧后的恢复响应,P.t、A.t.L.、A.a、C.s、C.p、L.c分别代表细叶白头翁、裂叶蒿、斜茎黄芪、糙隐子草、日阴菅、羊草。图中不同大写字母代表各放牧强度间有显著差异
a,b,c,d分别是6个关键物种的叶片干重、叶片含水量、叶面积和比叶面积对停止放牧后的恢复响应,P.t、A.t.L.、A.a、C.s、C.p、L.c分别代表细叶白头翁、裂叶蒿、斜茎黄芪、糙隐子草、日阴菅、羊草。图中不同大写字母代表各放牧强度间有显著差异

表1 不同放牧强度与不同物种对叶片功能性状的分析
加粗字体表示在0.01,0.05水平上有显著影响 Bold font indicates that it has a significant effect on 0.01 at 0.05 level
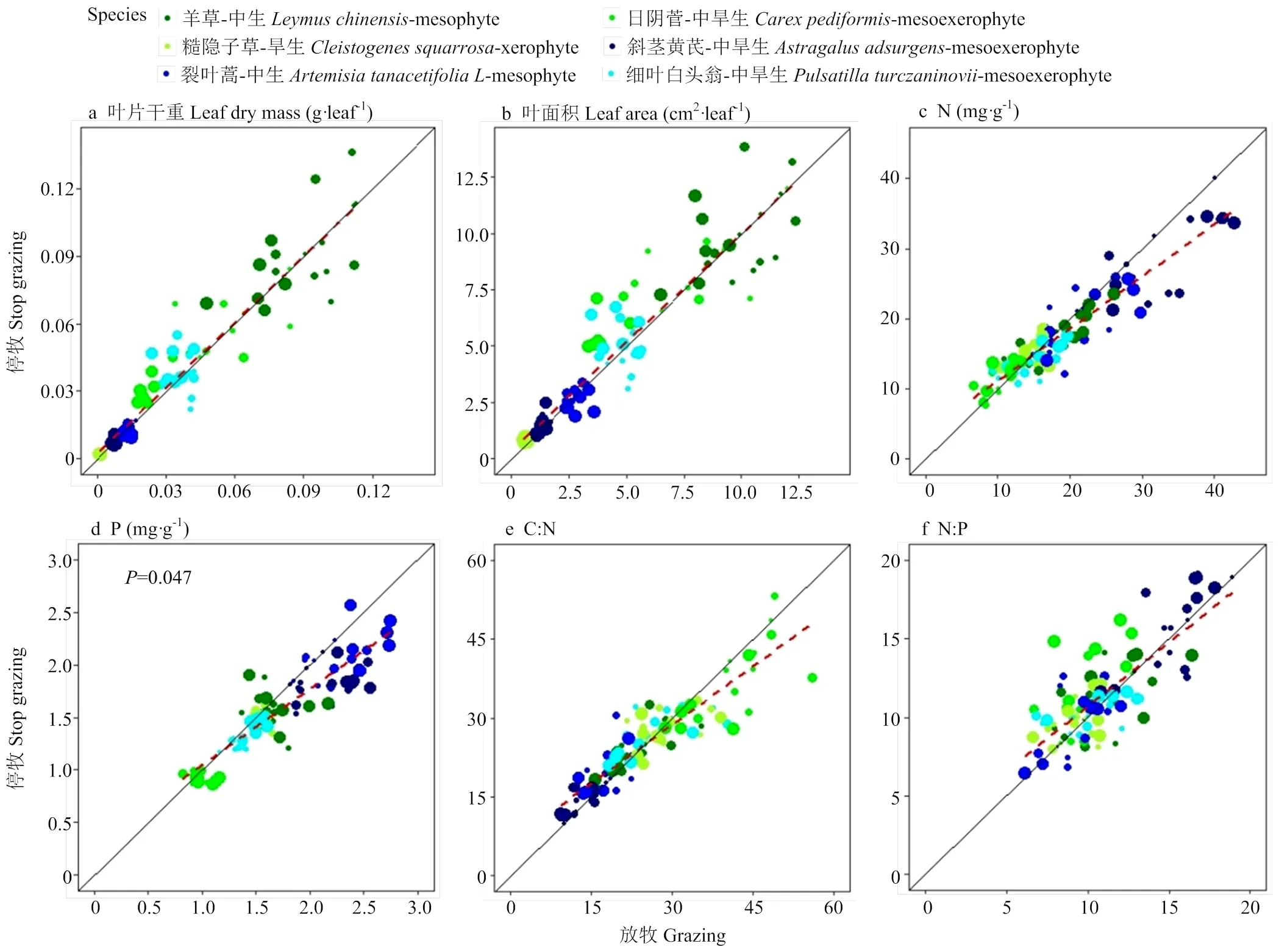
图中不同颜色代表不同物种,点的大小由小到大代表放牧强度由轻度到重度
3 讨论
自20世纪70年代以来,关于恢复力逐渐成为国内外学者研究的热点。最开始恢复力是以弹性,即弹簧特性来表明其稳定性,随后恢复力被美国学者HOLLING[29]首次引入生态学研究来表明系统稳定性,目前关于恢复力的研究很多但没有一个统一的定论,可以概括为生态系统自我恢复的强度和限度,即弹簧的拉伸程度和弹性强度。关于草原生态系统恢复力研究,不同的围封年限成为大家不同的关注点,大部分研究集中在长期围封后的恢复或者长期不同恢复年限下群落植被的恢复情况,部分研究集中在短期围封的恢复上,对短期围封的关注不够。本文所研究的短期围封停牧恢复—即时恢复响应,即弹簧拉伸后回到初始状态这一过程中的回弹瞬间,与贾志锋、朱牛、石明明等[27,30-31]的研究一致,停牧封育并非时间越长越好,短期围封的恢复或即时恢复在一定程度上,依靠草原生态系统自身弹性大大促进了草原的恢复,而长期围封停牧并不能持续增加草原生产力及其功能,不利于草原有效资源的利用。草原放牧相关研究的目的如李向林[32]所说是为了更好草原管理和草原生态系统的发展,所以除了长期的恢复研究外,还应有短期恢复研究,在短期恢复研究基础上进行长期研究,能更全面的为草原恢复和管理提供支持,以便有更好的草原管理。
植物功能性状能够响应生存环境的变化并(或)对生态系统功能有一定影响,其中,叶功能性状与植株生物量和植物对资源的获得和利用的关系最为密切,能够反映植物适应环境变化而形成的生存对策[33-35],通过研究植物叶片功能性状来反映其对环境的适应,具有重要意义。本研究采用基于植物叶片功能性状的方法来检验草甸草原关键物种对不同放牧强度的家畜放牧及停止放牧后的响应。
3.1 不同放牧强度对关键物种功能性状的影响
植物在一定程度上会通过改变自身功能性状来实现植物个体小型化[15],植物个体小型化可以使被采食性降低,增加采食难度,从而达到躲避放牧的效果[36]。本研究中的6个关键物种的花果期除日阴菅和细叶白头翁在5—7月,其他几个物种的花果期整合起来在6—9月,我们的采样时间都在同一时间,可能物候期对植物功能性状影响不大。6个关键物种的叶片干重、叶面积随放牧强度的增加而减少,与其一致,说明了植株小型化。在低放牧强度干扰下,植物可以维持叶片的正常生长,在短时间达到物流平衡暂稳态,随着放牧强度增加,长期过度采食叶片则会导致叶面积指数下降,光合能力下降,叶片的生活力减弱,叶片干重降低[37]。比叶面积的大小能直接反映植物光合能力大小,比叶面积越大,光合能力越强,在一定程度上反映了植物的生长速率[38],也反映了叶片截获光的能力和在强光下的自我保护能力,往往与植物的生长和生存对策紧密相关,能够反映植物在不同生境下的资源获取能力。本研究结果显示,6个关键物种的叶片干重和叶面积随放牧强度的增加减小,而比叶面积整体都呈增大的趋势,表明随放牧强度的增加,叶片虽然变小但变薄了,提高了叶片获取光资源的能力,一定程度上促进了植物的光合作用,提高了植物的生长速率。植物叶片含水量是反映植物组织水分状况的重要指标,是植物正常生理活动的重要因素,水分和营养平衡是随环境变化而变化的[35],当叶片含水量较为充足时,其生理活动较为活跃,代谢较快,本研究关键物种的叶片含水量随放牧强度的增加,整体呈增加趋势,并且与叶片干物质含量呈相反的变化趋势,为叶片的光合作用提供了良好的基础。整体来说,叶片随放牧强度增加而产生的变化叶片变小变薄且含水量增大,可能增加了叶片对CO2的吸收,有利于叶片进行光合作用,提高了叶片光合生产能力。
植物碳氮磷为植物生长的大量元素,氮是蛋白质、核酸、磷脂及其他植物生长发育所必需的有机氮化合物的构成成分,而这些物质是细胞赖以生存的结构或功能成分,因此氮被称为“生命元素”。磷与细胞的能量代谢,各种有机物的合成和分解代谢等几乎所有的生命过程密切相关。植物氮、磷含量都表现为:非禾草>禾草;与非豆科相比,豆科植物相应器官具有更高的氮含量,在组织水平上,禾草比非禾草、非豆科植物比豆科植物具有更高的养分利用效率[39]。本研究结果与此结论整体一致,6个关键物种的氮、磷含量也表现为非禾草>禾草,斜茎黄芪、裂叶蒿、细叶白头翁整体叶片氮、磷含量>羊草、日阴菅、糙隐子草,且豆科植物斜茎黄芪的叶片氮含量最高,叶片氮、磷含量整体随放牧强度的增加表现出增加的变化趋势,这可能是因为高放牧强度的样地内,由于牛采食了植物,刺激幼嫩叶片的快速生长,从而导致了叶片内氮、磷含量的增加,因为幼嫩植物的快速生长需要氮、磷元素。叶片C/N在一定程度上反映了植物的生长速率,生长速率理论表示生长速率较高的生物具有较低的C/N[40],C/N下降表明草地在退化,C/N越低,退化越严重[41]。本研究表明,随放牧强度的增加,6个关键物种的碳氮比都呈减小趋势,从而提高了植物的生长速率,这可能是因为叶片氮含量的增加而导致的。植物氮磷比可作为判定植物营养是否受限的一个指标。KOERSELMAN等[42]在对欧洲湿地植物的研究发现,如果植物氮磷比<14,说明植物受到氮的制约,如果植物氮磷比>16,说明植物受磷的制约。本研究结果显示,整体来看,斜茎黄芪在14—16之间,为氮磷共同限制,其余5个关键物种的氮磷比都<14,说明受到氮的限制,与王淼等[25]的研究结果一致。
3.2 关键物种功能性状对停止放牧的响应
围栏封育是停止放牧的一种方式,其对草地的影响一是直接减少牲畜的采食和践踏,另一个是通过牲畜对土壤结构及土壤水分的干扰而间接影响着植物[43],影响草地生态系统的物质生产和能量分配。本研究中,6个关键物种的相对生物量和重要值在轻度和重度放牧强度恢复较好,功能性状对停止放牧后的响应不一,叶片干重、叶面积的响应较为一致,且叶片的形态学性状对停止放牧的响应都为正响应远大于负响应,物种对性状的影响大于放牧强度,说明解除放牧压力后,植物叶片的形态学性状的恢复生长优于家畜采食后植物的补偿生长。由此得出,轻度放牧在一定程度上,放牧对植物功能的影响大于停止放牧,这可能是植物对放牧躲避的一种策略。整体来看,停止放牧后,植物的叶片形态学性状正响应>负响应,恢复较好,由于植物物种不同,各物种对停止放牧后的响应不同,其中禾草类对停止放牧后的恢复的敏感度大于杂类草。
由于家畜采食,减少了植物的衰老组织,不断刺激植物生长,尤其是幼嫩组织(叶片)的再生,促进氮磷向幼嫩组织的重新分配,同时家畜的粪便也可能是导致放牧区的叶片氮磷含量普遍高于停牧区。本研究结果显示,叶片磷含量对不同放牧强度的负响应多于叶片氮含量,这可能是因为家畜采食促进了植物(尤其是幼嫩叶片)的光合作用,提高了叶片中的叶绿素含量[44-45],而氮元素作为叶绿素的主要组成部分,不仅仅取决于土壤中的氮含量,是植物自身和外部环境共同权衡的,而植物吸收的磷元素,更多来自于土壤中[45],放牧区土壤中的氮磷元素高于停牧区的。碳氮比的值越低代表植物生长速率越快[40],在本研究中,叶片C/N对放牧地响应,杂类草较为一致,都为正响应,即停止放牧后地叶片C/N大于放牧,但禾草类的响应并不一致,羊草不同放牧强度长期放牧停止后的响应为正响应,日阴菅为负响应,糙隐子草在轻度和重度为正响应,在中度为负响应,这可能是因为物种不同,他们的生长速率不同,不同遗传性状的植物在相同条件下,植物体内的含量会有差别。6个关键物种叶片N/P对停止放牧后的响应基本一致,停止放牧后的叶片N/P>放牧下的叶片N/P,这与许雪贇等[46]的研究一致,这可能是因为植物叶片氮含量的相对变化大于叶片磷含量的相对变化。对于养分特征结果得出,停止放牧后恢复较好,促进了植物的再生,杂类草对停止放牧后的恢复的敏感度大于禾草类。总的来说,停止放牧后,植物叶片形态学性状和养分特征的正响应,在一定程度上促进了草甸草原植被功能的恢复,提高了牧草品质。
4 结论
在长期不同强度放牧下,植物出现小型化,叶片随放牧强度增加而变小变薄,叶片氮、磷含量逐渐增加,伴随植物的补偿性生长,植物的生长速率和光合作用随放牧强度的增加而增大,物种间竞争变大。停止放牧后,6个关键物种的叶片功能性状在重度放牧下恢复较轻度放牧好,正响应>负响应,促进了草甸草原的恢复。在放牧与停牧恢复下,植物对有效资源的竞争而出现的变化和不同停牧恢复时间的恢复响应仍需进一步研究,为草原恢复和管理提供一定的数据支持,有助于提高草甸草原的生产力。
[1] 张骞, 马丽, 张中华, 徐文华, 周秉荣, 宋明华, 乔安海, 王芳, 佘延娣, 杨晓渊, 郭婧, 周华坤. 青藏高寒区退化草地生态恢复: 退化现状、恢复措施、效应与展望. 生态学报, 2019, 39(20): 7441-7451.
ZHANG Q, MA L, ZHANG Z H, XU W H, ZHOU B R, SONG M H, QIAO A H, WANG F, SHE Y D, YANG X Y, GUO J, ZHOU H K. Ecological restoration of degraded grassland in Qinghai-Tibet alpine region: degradation status, restoration measures, effects and prospects. Acta Ecologica Sinica, 2019, 39(20): 7441-7451. (in Chinese)
[2] 刘兴元, 龙瑞军, 尚占环. 草地生态系统服务功能及其价值评估方法研究. 草业学报, 2011, 20(1): 167-174.
LIU X Y, LONG R J, SHANG Z H. Evaluation method of ecological services function and their value for grassland ecosystems. Acta Prataculturae Sinica, 2011, 20(1): 167-174. (in Chinese)
[3] 高军靖. 呼伦贝尔草原生态安全评价研究[D]. 北京: 中国环境科学研究院, 2013.
GAO J J. Study on ecological safety assessment in Hulunbeier steppe[D]. Beijing: Chinese Research Academy of Environmental Sciences, 2013. (in Chinese)
[4] 胡志超, 李政海, 周延林, 鲍雅静, 张靖, 黄朔, 元征征, 洪光宇, 张文海. 呼伦贝尔草原退化分级评价及时空格局分析. 中国草地学报, 2014, 36(5): 12-18.
HU Z C, LI Z H, ZHOU Y L, BAO Y J, ZHANG J, HUANG S, YUAN Z Z, HONG G Y, ZHANG W H. The partitioning of degradation and analysis of its spatio-temporal pattern of Hulunbeier grassland, China. Chinese Journal of Grassland, 2014, 36(5): 12-18. (in Chinese)
[5] 贺鹏程, 叶清. 基于植物功能性状的生态学研究进展: 从个体水平到全球尺度. 热带亚热带植物学报, 2019, 27(5): 523-533.
HE P C, YE Q. Plant functional traits: from individual plant to global scale. Journal of Tropical and Subtropical Botany, 2019, 27(5): 523-533. (in Chinese)
[6] MCINTYRE S, LAVOREL S, LANDSBERG J, FORBES T D A. Disturbance response in vegetation–towards a global perspective on functional traits. Journal of Vegetation Science, 1999, 10(5): 621-630.
[7] LAVOREL S, GRIGULIS K, LAMARQUE P, COLACE M P, GARDEN D, GIREL J, PELLET G, DOUZET R. Using plant functional traits to understand the landscape distribution of multiple ecosystem services. Journal of Ecology, 2011, 99(1): 135-147.
[8] 赵娜, 赵新全, 赵亮, 徐世晓, 邹小艳. 植物功能性状对放牧干扰的响应. 生态学杂志, 2016, 35(7): 1916-1926.
ZHAO N, ZHAO X Q, ZHAO L, XU S X, ZOU X Y. Progress in researches of the response of plant functional traits to grazing disturbance. Chinese Journal of Ecology, 2016, 35(7): 1916-1926. (in Chinese)
[9] WRIGHT I J, REICH P B, WESTOBY M, ACKERLY D D, BARUCH Z, BONGERS F, CAVENDER-BARES J, CHAPIN T, CORNELISSEN J H C, DIEMER M, et al. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827.
[10] MOONEY K A, HALITSCHKE R, KESSLER A, AGRAWAL A A. Evolutionary trade-offs in plants mediate the strength of trophic cascades. Science, 2010, 327(5973): 1642-1644.
[11] ORWIN K H, MASON N W H, JORDAN O M, LAMBIE S M, STEVENSON B A, MUDGE P L. Season and dominant species effects on plant trait-ecosystem function relationships in intensively grazed grassland. Journal of Applied Ecology, 2018, 55(1): 236-245.
[12] 晁永芳. 放牧活动对草原植物功能性状及其权衡关系的调控. 当代畜牧, 2018(2): 1-2.
CHAO Y F. Regulation of grazing activities on functional traits of grassland plants and their trade-offs. Contemporary Animal Husbandry, 2018(2): 1-2. (in Chinese)
[13] 张月强. 探究泰山木本植物功能性状间的关系及对环境因子的响应[D]. 济南: 山东大学, 2014.
ZHANG Y Q. Effects of environmental factors on plant functional traits and the relationship between functional traits in Taishan woody plants[D]. Jinan: Shandong University, 2014. (in Chinese)
[14] GARCÍA-CERVIGÓN A I, GARCÍA-LÓPEZ M A, PISTÓN N, PUGNAIRE F I, OLANO J M. Co-ordination between xylem anatomy, plant architecture and leaf functional traits in response to abiotic and biotic drivers in a nurse cushion plant. Annals of Botany, 2021, 127(7): 919-929.
[15] 侯路路, 闫瑞瑞, 张宇, 辛晓平. 放牧强度对草甸草原羊草功能性状的影响. 中国农业科学, 2020, 53(13): 2562-2572.
HOU L L, YAN R R, ZHANG Y, XIN X P. Effects of grazing intensity on functional traits ofin meadow steppe. Scientia Agricultura Sinica, 2020, 53(13): 2562-2572. (in Chinese)
[16] 李西良, 侯向阳, 吴新宏, 萨茹拉, 纪磊, 陈海军, 刘志英, 丁勇. 草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应. 植物生态学报, 2014, 38(5): 440-451.
LI X L, HOU X Y, WU X H, SA R L, JI L, CHEN H J, LIU Z Y, DING Y. Plastic responses of stem and leaf functional traits into long-term grazing in a meadow steppe. Chinese Journal of Plant Ecology, 2014, 38(5): 440-451. (in Chinese)
[17] 王炜, 刘钟龄, 郝敦元, 梁存柱. 内蒙古草原退化群落恢复演替的研究: Ⅰ.退化草原的基本特征与恢复演替动力. 植物生态学报, 1996, 20(5): 449-459.
WANG W, LIU Z L, HAO D Y, LIANG C Z. Research on the restoring succession of the degenerated grassland in inner mongolia──i. basic characteristics and driving force for restoration of the degenerated grassland. Acta Phytoecologica Sinica, 1996, 20(5): 449-459. (in Chinese)
[18] ZIRBEL C R, BASSETT T, GRMAN E, BRUDVIG L A. Plant functional traits and environmental conditions shape community assembly and ecosystem functioning during restoration. Journal of Applied Ecology, 2017, 54(4): 1070-1079.
[19] 吴思雨, 宝音陶格涛, 许宏斌, 张璐. 放牧强度对内蒙古典型草原糙隐子草功能性状的影响. 应用生态学报, 2021, 32(2): 392-398.
WU S Y, BAO Y, XU H B, ZHANG L. Effects of grazing intensities on functional traits ofin a typical grassland of Inner Mongolia, China. Chinese Journal of Applied Ecology, 2021, 32(2): 392-398. (in Chinese)
[20] 邢小青, 张璐, 宝音陶格涛, 杨兆平. 放牧对糙隐子草地上个体功能性状的影响. 中国草地学报, 2019, 41(6): 116-122.
XING X Q, ZHANG L, BAO Y, YANG Z P. Effect of grazing on the aboveground functional traits of. Chinese Journal of Grassland, 2019, 41(6): 116-122. (in Chinese)
[21] 邢小青, 郝匕台, 齐丽雪, 张璐, 宝音陶格涛. 放牧对克氏针茅功能性状的影响. 生态环境学报, 2019, 28(1): 57-64.
XING X Q, HAO B T, QI L X, ZHANG L, BAO Y. Effect of grazing on functional traits of. Ecology and Environmental Sciences, 2019, 28(1): 57-64. (in Chinese)
[22] 张景慧, 王铮, 黄永梅, 陈慧颖, 李智勇, 梁存柱. 草地利用方式对温性典型草原优势种植物功能性状的影响. 植物生态学报, 2021, 45(8): 818-833.
ZHANG J H, WANG Z, HUANG Y M, CHEN H Y, LI Z Y, LIANG C Z. Effects of grassland utilization on the functional traits of dominant plants in a temperate typical steppe. Chinese Journal of Plant Ecology, 2021, 45(8): 818-833. (in Chinese)
[23] YE R H, LIU G F, CHANG H, SHAN Y M, MU L, WEN C, TE R, WU N T, SHI L, LIU Y H, WANG H M, YUN X J, LIU G X, LI F. Response of plant traits ofto grazing intensity and fluctuation in annual precipitation in a desert steppe, Northern China. Global Ecology and Conservation, 2020, 24: e01237.
[24] BAI Y F, WU J G, CLARK C M, PAN Q M, ZHANG L X, CHEN S P, WANG Q B, HAN X G. Grazing alters ecosystem functioning and C: N: P stoichiometry of grasslands along a regional precipitation gradient. Journal of Applied Ecology, 2012, 49(6): 1204-1215.
[25] 王淼, 张宇, 李瑞强, 辛晓平, 朱晓昱, 曹娟, 周忠义, 闫瑞瑞. 放牧强度对羊草草甸草原植物器官及群落氮磷化学计量的影响. 中国农业科学, 2022, 55(7): 1371-1384.
WANG M, ZHANG Y, LI R Q, XIN X P, ZHU X Y, CAO J, ZHOU Z Y, YAN R R. Effects of grazing disturbance on the stoichiometry of nitrogen and phosphorus in plant organs ofmeadow steppe. Scientia Agricultura Sinica, 2022, 55(7): 1371-1384. (in Chinese)
[26] HE M A, ZHOU G Y, YUAN T F, GROENIGEN K J, SHAO J J, ZHOU X H. Grazing intensity significantly changes the C: N: P stoichiometry in grassland ecosystems. Global Ecology and Biogeography, 2020, 29(2): 355-369.
[27] 石明明, 牛得草, 王莹, 袁晓波, 贺磊, 韩炳宏, 宗文杰, 傅华. 围封与放牧管理对高寒草甸植物功能性状和功能多样性的影响. 西北植物学报, 2017, 37(6): 1216-1225.
SHI M M, NIU D C, WANG Y, YUAN X B, HE L, HAN B H, ZONG W J, FU H. Effect of fencing and grazing management on the plant functional traits and functional diversity in an alpine meadow on the Tibetan Plateau. Acta Botanica Boreali-Occidentalia Sinica, 2017, 37(6): 1216-1225. (in Chinese)
[28] 闫瑞瑞, 辛晓平, 张保辉, 闫玉春, 杨桂霞. 肉牛放牧梯度对呼伦贝尔草甸草原植物群落特征的影响. 中国草地学报, 2010, 32(3): 62-67.
YAN R R, XIN X P, ZHANG B H, YAN Y C, YANG G X. Influence of cattle grazing gradient on plant community characteristics in Hulunber meadow steppe. Chinese Journal of Grassland, 2010, 32(3): 62-67. (in Chinese)
[29] HOLLING C S. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics, 1973, 4: 1-23.
[30] 贾志锋, 马祥, 徐成体, 刘文辉, 魏小星, 雷生春. 短期封育对贵南县轻度退化高寒草甸植被特征的影响. 草业科学, 2019, 36(11): 2766-2774.
JIA Z F, MA X, XU C T, LIU W H, WEI X X, LEI S C. Effects of short-term enclosure on the vegetation characteristics of a lightly degraded alpine meadow in Guinan County. Pratacultural Science, 2019, 36(11): 2766-2774. (in Chinese)
[31] 朱牛, 孙建, 石凝, 王金牛, 张林, 罗栋梁, 申承, 盖艾鸿. 短期围栏封育对高寒草甸植物群落及土壤理化性质的影响. 草地学报, 2023, 31(3): 834-843.
ZHU N, SUN J, SHI N, WANG J N, ZHANG L, LUO D L, SHEN C, GAI A H. Effects of short-term fence enclosing on plant community and the physical and chemical properties of alpine meadow soils. Acta Agrestia Sinica, 2023, 31(3): 834-843. (in Chinese)
[32] 李向林. 草原管理的生态学理论与概念模式进展. 中国农业科学, 2018, 51(1): 191-202.
LI X L. Advances in ecological theories and management models regarding rangeland management. Scientia Agricultura Sinica, 2018, 51(1): 191-202. (in Chinese)
[33] GARNIER E, SHIPLEY B, ROUMET C, LAURENT G. A standardized protocol for the determination of specific leaf area and leaf dry matter content. Functional Ecology, 2001, 15(5): 688-695.
[34] 刘金环, 曾德慧, Don Koo Lee. 科尔沁沙地东南部地区主要植物叶片性状及其相互关系. 生态学杂志, 2006, 25(8): 921-925.
LIU J H, ZENG D H, LEE D K. Leaf traits and their interrelationships of main plant species in southeast Horqin sandy land. Chinese Journal of Ecology, 2006, 25(8): 921-925. (in Chinese)
[35] VENDRAMINI F, DÍAZ S, GURVICH D E, WILSON P J, THOMPSON K, HODGSON J G. Leaf traits as indicators of resource- use strategy in floras with succulent species. New Phytologist, 2002, 154(1): 147-157.
[36] ZHENG S X, REN H Y, LAN Z C, LI W H, WANG K B, BAI Y F. Effects of grazing on leaf traits and ecosystem functioning in Inner Mongolia grasslands: scaling from species to community. Biogeosciences, 2010, 7(3): 1117-1132.
[37] SMITH S E. Variation in response to defoliation between populations ofvar.(Poaceae) with different livestock grazing histories. American Journal of Botany, 1998, 85(9): 1266-1272.
[38] LAPORTE M A, GARNIER E. ThesauForm—Traits: a web based collaborative tool to develop a thesaurus for plant functional diversity research. Ecological Informatics, 2012, 11: 34-44.
[39] TJOELKER M G, CRAINE J M, WEDIN D, REICH P B, TILMAN D. Linking leaf and root trait syndromes among 39 grassland and savannah species. The New Phytologist, 2005, 167(2): 493-508.
[40] 牛得草, 董晓玉, 傅华. 长芒草不同季节碳氮磷生态化学计量特征. 草业科学, 2011, 28(6): 915-920.
NIU D C, DONG X Y, FU H. Seasonal dynamics of carbon, nitrogen and phosphorus stoichiometry in. Pratacultural Science, 2011, 28(6): 915-920. (in Chinese)
[41] 李丹, 康萨如拉, 赵梦颖, 张庆, 任海娟, 任婧, 周俊梅, 王珍, 吴仁吉, 牛建明. 内蒙古羊草草原不同退化阶段土壤养分与植物功能性状的关系. 植物生态学报, 2016, 40(10): 991-1002.
LI D, KANGSARULA, ZHAO M Y, ZHANG Q, REN H J, REN J, ZHOU J M, WANG Z, WU R J, NIU J M. Relationships between soil nutrients and plant functional traits in different degradation stages ofsteppe in Nei Mongol, China. Chinese Journal of Plant Ecology, 2016, 40(10): 991-1002. (in Chinese)
[42] KOERSELMAN W, MEULEMAN A F M. The vegetation N: P ratio: a new tool to detect the nature of nutrient limitation. The Journal of Applied Ecology, 1996, 33(6): 1441.
[43] 聂莹莹, 辛晓平, 徐丽君, 杨桂霞. 封育措施对呼伦贝尔草甸草原区草地生产力的影响. 中国农业资源与区划, 2022, 43(8): 74-82.
NIE Y Y, XIN X P, XU L J, YANG G X. Effects of enclosure measures on grassland and productivity in Hulunbeier meadow steppe. Chinese Journal of Agricultural Resources and Regional Planning, 2022, 43(8): 74-82. (in Chinese)
[44] 翟夏杰, 黄顶, 王堃. 围封与放牧对典型草原植被和土壤的影响. 中国草地学报, 2015, 37(6): 73-78.
ZHAI X J, HUANG D, WANG K. Effects of fencing and grazing on vegetation and soil in typical grassland. Chinese Journal of Grassland, 2015, 37(6): 73-78. (in Chinese)
[45] NIU K C, HE J S, ZHANG S T, LECHOWICZ M J. Tradeoffs between forage quality and soil fertility: lessons from Himalayan rangelands. Agriculture, Ecosystems & Environment, 2016, 234: 31-39.
[46] 许雪贇, 曹建军, 杨淋, 杨书荣, 龚毅帆, 李梦天. 放牧与围封对青藏高原草地土壤和植物叶片化学计量学特征的影响. 生态学杂志, 2018, 37(5): 1349-1355.
XU X Y, CAO J J, YANG L, YANG S R, GONG Y F, LI M T. Effects of grazing and enclosure on foliar and soil stoichiometry of grassland on the Qinghai-Tibetan Plateau. Chinese Journal of Ecology, 2018, 37(5): 1349-1355. (in Chinese)
Response of Functional Traits of Key Species in Meadow Steppe to Long-Term Grazing and Grazing Exclusion

1College of Prataculture and Grassland,Northwest A&F University,Yangling 712100, Shaanxi;2National Field Scientific Observation and Research Station of Hulunbuir Grassland Ecosystem / Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences, Beijing 100081;3College of Ecology, Lanzhou University, Lanzhou 730000;4School of Agriculture and Forestry, Hulunbuir University, Hulunbuir 021008, Inner Mongolia
【Objective】The response of plant functional traits to environmental changes and external disturbances reflects the phenotypic plasticity, survival strategies and adaptability of plants. The study on the changes of functional traits of grassland plants under long-term grazing and the recovery response after stopping grazing can provide a theoretical basis for grassland restoration management.【Method】In this study, the changes of leaf functional traits (leaf dry mass, leaf total water content, leaf area, specific leaf area, leaf nitrogen content, leaf phosphorus content, leaf C/N raito, leaf N/P ratio) of six key species () in Hulunbuir meadow steppe under different grazing intensities during growing season were studied. The immediate recovery response of leaf functional traits of key species in meadow steppe after stopping grazing during growing season was discussed.【Result】(1) The relative biomass and importance values of the six key species decreased with the increase of grazing intensity, and the values of light and heavy grazing intensity after stopping grazing were greater than that after continuing grazing. (2)The leaf dry mass and leaf C/N ratio of six key species decreased with the increase of grazing intensity; the leaf total water content, specific leaf area and leaf nitrogen content increased with the increase of grazing intensity; the leaf area of all species exceptdecreased with the increase of grazing intensity; the leaf phosphorus content increased with the increase of grazing intensity except; with the increase of grazing intensity, the leaf N/P ratio ofshowed an increasing trend, while that ofL. decreased. (3) After stopping grazing, the resilience of heavy grazing was higher than that of light grazing, and the functional traits of 6 key species recovered better as a whole, leaf dry mass, leaf total water content, leaf area, specific leaf area and leaf N/P ratio showed positive response, leaf phosphorus content showed negative response, leaf nitrogen content and leaf C/N ratio showed different positive and negative responses among six key species, the negative response of leaf nitrogen content was less than positive response, and the positive response of leaf C/N was more than negative response. (4) The leaf morphological traits of grasses and the nutrient characteristics of miscellaneous grasses were more sensitive to recovery after stopping grazing, and the responses of leaf dry mass, leaf area and leaf phosphorus content to the interaction between grazing intensity and species were more significant. After stopping grazing, leaf dry mass of leaves increased with the increase of leaf area, leaf nitrogen content and leaf N/P of the six key species.【Conclusion】Long-term grazing led to individual miniaturization of grassland plants. After stopping grazing, the positive response of plant functional traits is more than negative response, and the grassland vegetation function shows an obvious recovery response, which increases the productivity of the grassland. Above-ground biomass of the meadow grassland increases.
long-term grazing; stop grazing; leaf functional traits; resilience; meadow steppe
10.3864/j.issn.0578-1752.2023.18.016
2022-10-22;
2023-06-15
国家自然科学基金重点项目(32130070)、国家重点研发计划(2021YFD1300500)
司雨凡,E-mail:yufan@nwafu.edu.cn。通信作者辛晓平,E-mail:xinxiaoping@caas.cn
(责任编辑 林鉴非)
