基于SSR 标记的100 份国内外小麦种质遗传多样性分析及DNA 指纹图谱构建
2023-10-20王健胜侯桂玲王二伟马爱锄程世平
王健胜侯桂玲王二伟马爱锄程世平
(1. 平顶山学院,河南平顶山 467000;2. 河南省生态经济型木本植物种质创新与利用重点实验室,河南平顶山 467000;3. 平顶山市农业科学院,河南平顶山 467001)
小麦(Triticum aestivumL.)具有较强的环境适应性,我国从北方到南方均有种植,具有分布范围广、栽培面积大、总产量高等特点,是我国的主要粮食作物之一,对保障国家粮食安全具有重要战略意义[1]。 优良的品种是保障我国小麦高产稳产的重要基础。 但在育种实践中,为了获得农艺性状优良的小麦品种,育种工作者习惯于过分注重对少数优良小麦种质的利用,这使得育成小麦种质的遗传基础日益狭窄,小麦育种水平及进度提高缓慢,故种质资源研究作为小麦新品种选育的重要基础性工作越来越受到科研工作者的重视[2-3]。 因此,开展种质遗传多样性分析,掌握现有种质的遗传基础及亲缘关系,发掘并利用优异种质材料,对拓宽小麦育种的遗传基础、加快育种进程和提高育种水平具有重要意义。
已有学者开展了小麦遗传多样性研究[4-8],如张凡等[4]对620 份小麦种质农艺性状的遗传多样性进行了研究,发现其在质量性状和数量性状上均具有丰富的遗传多样性,并据此将这620 份小麦种质分为10 个类群;通过综合评价,筛选出91 份较为优异的种质材料,可作为亲本材料用于黄淮麦区小麦育种改良中。 近年来,分子标记技术快速发展,因其具有多态性丰富、遗传信息量大、不受外界环境条件影响等优点而在小麦遗传多样性研究中得到较为广泛的应用[9]。 例如卢丽斌等[10]利用AFLP 技术对安徽省小麦赤霉病菌4 个地理群体的68 个供试菌株进行了群体遗传多样性分析,结果发现4 个群体间的遗传距离与地理分布没有明显的相关性,遗传变异主要存在于群体内,群体间的遗传变异较小;韩芳等[11]采用SRAP 分子标记对70 份黄淮麦区小麦品种的遗传多样性进行了分析,发现河南省小麦品种的多样性指数最高,江苏省小麦品种多样性指数最低;罗琴等[12]利用42 条ISSR 引物对11 份人工合成小麦和3 个普通小麦材料进行了遗传多样性分析,发现材料间的平均遗传相似系数为0.790,表明这14 份小麦材料亲缘关系较近。
SSR(simple sequence repeats)作为一种重要类型分子标记,具有分布位点多、多态性丰富、易于操作、可覆盖整个基因组等优点,目前已被广泛应用于小麦遗传多样性研究中[13-14]。 赵檀等[15]利用覆盖小麦全基因组的SSR 标记对河北省1949 年至2012 年审定的169 份小麦品种进行遗传多样性分析,结果认为,自1949 年以来,虽然品种的等位基因频率在下降,但多样性水平却呈现上升趋势;根据遗传相似系数,供试品种可划分为5 大类。
虽然前人利用分子标记技术对小麦遗传多样性进行了较多研究,但由于试验材料和研究方法不同,研究结果并不具有通用性,且许多研究还存在试验材料来源单一且只针对国内小麦种质的问题。 本试验以来自不同国家及我国不同区域的小麦品种(系)和地方农家种等为材料,利用SSR 标记对其遗传多样性进行研究,以全面揭示这些材料间的遗传差异及亲缘关系,同时构建每个小麦种质的DNA 指纹图谱,以期为小麦种质资源保护及新品种培育提供有效材料支撑。
1 材料与方法
1.1 供试材料及田间种植
试验材料为来自国内外的100 份小麦种质(表1),其中,国外种质11 份,包括澳大利亚的4份,法国的4 份,保加利亚、罗马尼亚和墨西哥的各1 份;国内种质89 份,包括河南的47 份,陕西的8 份,北京的6 份,江苏的6 份,山东的6 份,河北的5 份,山西的3 份,四川的2 份,贵州的2 份,安徽的1 份,宁夏的1 份。

表1 本研究所用小麦种质信息

表2 本研究所用SSR 引物的信息
2021 年10 月将这100 份小麦材料种植于河南省平顶山市农业科学院小麦基地。 每个材料设置3 个重复,每个重复种3 行,每行行长2 m,行距25 cm。 田间管理与普通大田相同。
1.2 基因组DNA 提取及PCR 扩增
在小麦苗期采集叶片材料,用SDS 法提取小麦基因组DNA[16],TE溶解后用SSR引物进行PCR 扩增。 PCR 扩增反应总体积为15 μL,包括1 μL 基因组DNA、2 μL 上下游引物、5 μL 2×BlueTaqPCR Mix、7 μL ddH2O。 在Eppendorf PCR 扩增仪上进行SSR-PCR 扩增,扩增程序如下:94 ℃预变性3 min;94 ℃变性1 min,Tm 复性1 min,72 ℃延伸1 min,35 个循环;72 ℃延伸5 min。SSR 引物一部分来自网站http:/ /wheat.pw.usda.gov/ggpages/DEM/ggtabledefs.html,另一部分来自文献[17],均由上海生工生物工程有限公司合成。
1.3 电泳检测及带型统计
PCR 扩增产物用8%聚丙烯酰胺凝胶电泳检测,保存胶片并进行带型统计,处于同一位点的有带记为“1”、无带记为“0”、缺失记为“9”,获得由“0、1、9”组成的数据矩阵,用于下一步分析。
1.4 数据统计分析
采用软件NTSYS 中的非加权平均法(UPMGA)计算小麦种质间的遗传相似系数,并对小麦种质进行聚类分析;统计分析不同引物的扩增总条带数、多态性条带数、多态性条带百分比,利用POPGEN 32 软件估算SCoT 引物和ISSR 引物的遗传参数,包括Nei’s 基因多样性指数、有效等位基因数、Shannon’s 信息指数等。
2 结果与分析
2.1 国内外小麦种质资源SSR 多态性分析
首先利用表型性状差异较大的两个小麦种质对125 对SSR 引物进行筛选,最终选择条带清晰、多态性丰富的23 对引物用于对100 份小麦种质进行检测分析,示例如图1,结果详见表3。 23对SSR 引物对100 份小麦种质扩增共获得235 个基因位点,其中多态性位点232 个,占比达到98.72%。 每对SSR 引物检测位点数介于6~18 之间,平均等位位点数为10.09 个;其中,Xcfa2153扩增出的位点最多,为18 个,而SSR26 和Xgwm539 扩增的位点最少,均只有6 个。 SSR 引物的多态性位点百分比表现较为突出,除Xwmc256、Xwmc335、Xgwm499 分别为87. 5%、88.9%、91.7%外,其余引物的多态性位点百分比均为100%,每对引物多态性位点百分比平均为98.61%。 有效等位基因数变幅为1.16 ~1.69,平均为1.35;Nei’s 基因多样性指数分布在0.14 ~0.39之间,平均为0.22;Shannon’s 信息指数的变化范围是0.26 ~0.58,平均值为0.36。 综合来看,本研究选择的SSR 引物在检测小麦遗传差异方面表现均较好。

图1 SSR144 对部分小麦种质的扩增情况
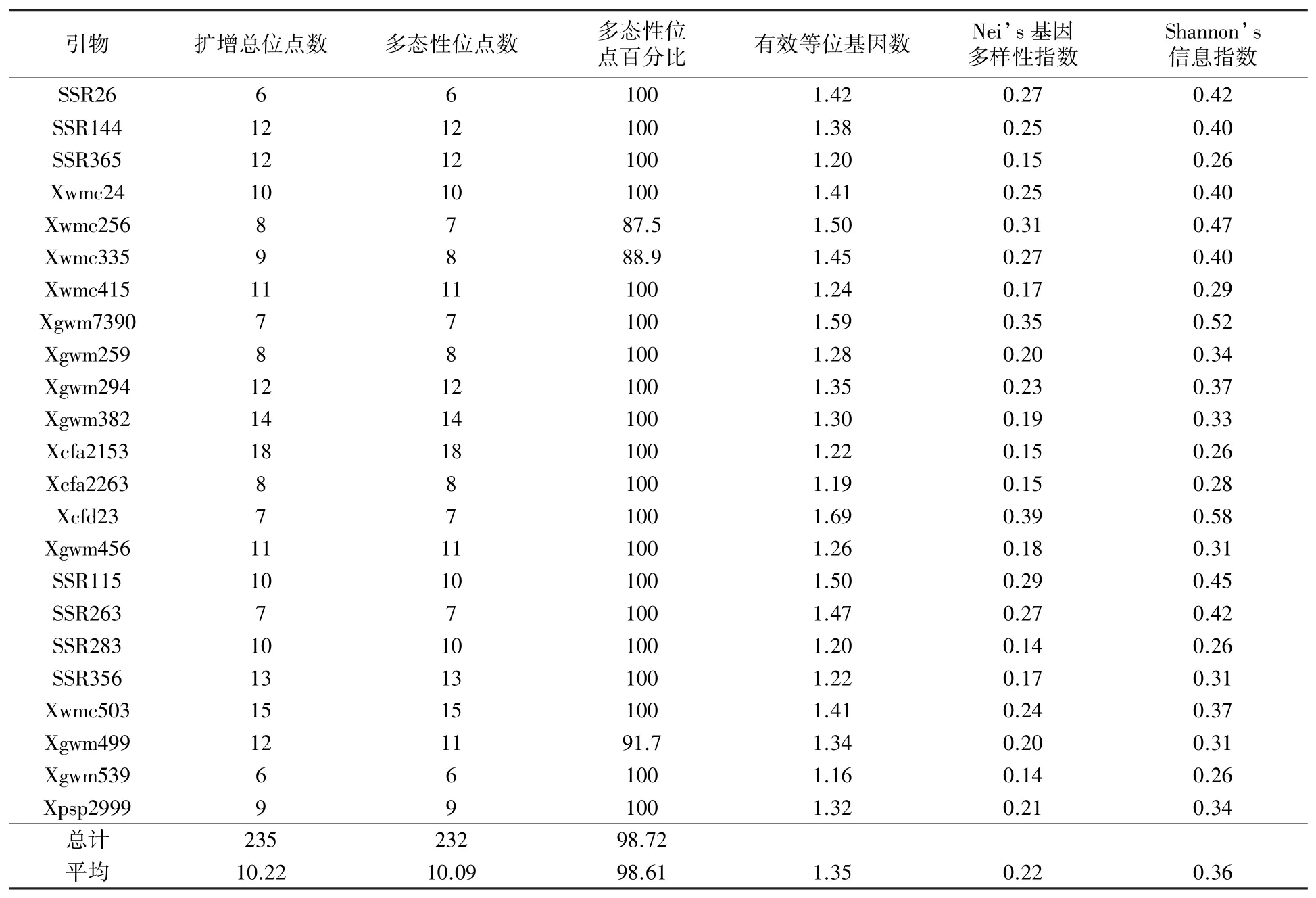
表3 本研究所用SSR 标记的遗传多样性参数
2.2 国内外小麦种质资源亲缘关系分析
遗传相似系数可以反映不同小麦种质间亲缘关系的远近。 基于SSR 引物检测到的不同小麦种质基因型数据,获得了100 份小麦种质间的Nei’s 遗传相似系数,变幅在0.56 ~0.96,平均为0.73,表明100 份小麦种质存在较丰富的遗传变异。 其中,来自甘肃的种质09N37 与来自江苏的种质10EW137 之间的遗传相似系数最小,说明这两个种质间的亲缘关系最远;而最大的遗传相似系数来自于种质洛麦21 号与偃展4110 之间,意味着两者间的亲缘关系最近,可能与它们均来自河南有关。 另外,超出平均遗传相似系数的小麦种质数占参试种质总数的50.99%,而低于平均遗传相似系数的小麦种质占参试品种总数的48.98%。
2.3 国内外小麦种质资源聚类分析
利用SSR 检测数据获得小麦种质间的遗传距离,以此构建100 份小麦种质的遗传聚类图谱,见图2。 可以看出,100 份小麦种质可被划分为6个类群,不同类群包含的小麦种质数存在较大差异。 第Ⅰ类群包含3 个小麦种质,即宿553、贵农17 和开麦21;第Ⅱ类群包含的小麦种质数最少,仅1 个,为来自河北的衡观35;第Ⅲ类群包含2个小麦种质,均来自国外,分别是来自澳大利亚的CD87 和来自法国的B08;第Ⅳ类群包括5 个小麦种质,分别是西峰9 号、临汾10 号、郑麦004、西农979 和04 中3804;第Ⅴ类群包含的小麦种质较多,共有27 个,占参试种质总数的27%,其中来自河南的小麦种质较多,其数量达到该类群总数的51.9%;第Ⅵ类群包含的小麦种质数最多,达到62个,占参试种质总数的62%,其中来自河南的种质占到46.8%。 另外发现,来自北京的5 个小麦种质被聚在同一个类群,来自法国的法国麦、Belero 和B20 也被划为一类,而且源自国外的小麦种质也基本被划分在同一类群。

图2 100 份小麦种质基于遗传距离的聚类分析结果
2.4 国内外小麦种质资源DNA 指纹图谱的构建
为了更有效地保护和利用小麦种质资源,从23 个SSR 引物中选取了多态性比较丰富的5 个SSR 引物,即Xwmc335、Xgwm294、Xcfd23、Xgwm499、Xwmc24,构建了100 份小麦种质的DNA指纹图谱,该DNA 指纹图谱共包括了47 个扩增变异位点,详见表4。 可以看出,每个小麦种质拥有自己特有的DNA 指纹特征谱带,这些谱带可以作为“身份证”用于对小麦种质进行有效区分、保护和利用。
3 讨论
随着生活水平的不断提高,人们对现代小麦生产提出了更高的要求,在保证高产的同时,优质、抗逆等特性也逐步成为育种专家追求的重要目标,这就要求小麦育种实现新的突破。 为了实现此目标,除了利用先进的育种技术外,丰富的种质资源是重要的前提和基础。 同时,随着国内外小麦种质资源交流的日益频繁,种质资源利用过程中经常会出现“异种同名”和“异名同种”等问题,影响了小麦种质资源的保护和利用。 因此,开展小麦种质资源研究已经成为一项重要课题。
遗传相似性系数可以直接反映不同小麦种质间亲缘关系的远近,可为小麦种质在下一步的遗传研究及育种实践中利用提供依据。 虽然前人已利用分子标记对小麦种质遗传多样性进行了较多分析[18-22],但由于研究材料和方法的不同,其遗传多样性程度差异较大,研究结果间的通用性较差,对其它研究参考价值有限。 如陈天青等[20]研究发现,贵州小麦育种核心种质的遗传相似系数为0.13 ~0.63,平均为0.33;Masmoudi 等[21]研究发现,突尼斯小麦种质遗传相似系数介于0.36 ~0.79 之间;徐志等[22]对四川小麦品种遗传多样性的研究发现,品种间遗传相似系数变化范围为0.62~1.00;而本研究对100 份国内外小麦种质的遗传多样性分析发现,供试小麦种质间的遗传相似系数变幅为0.56~0.96,平均为0.73。
另外,前人研究多存在材料来源单一等问题,而本研究选用的100 份小麦种质材料来源丰富,来自6 个国家,其中我国的小麦种质来自12 个不同省份或地区。 但从本研究与前人研究的比较中也发现,小麦种质遗传多样性水平与其来源地不存在必然的直接联系,即来源越丰富其遗传多样性水平不一定越高。 该结论也得到了聚类分析结果的支持,即不同来源小麦种质经常被划分在同一类群,而来自同一地区的小麦种质也会被划分在不同类群,这在一定程度上表明小麦种质亲缘关系远近与其是否来自同一地区不存在必然的联系。 这与喻俊杰[23]、李鲜花[24]等的研究结果存在一定的差异。
DNA 指纹图谱可以为小麦种质的科学保护和利用提供有效支撑,但前人开展该图谱构建的研究相对较少,如李红琴等[25]利用SSR 标记构建了青海省审定小麦品种的分子身份证。 本研究利用变异位点丰富的5 个SSR 标记构建了100 份小麦种质的DNA 指纹图谱,且图谱更加精细,更有利于对小麦种质进行区分和保护。 综上,本研究结果可为供试小麦种质深入研究及有效利用提供有用信息。
4 结论
本研究利用SSR 分子标记分析了国内外100份小麦种质的遗传多样性。 选用的23 对SSR 引物在扩增位点数、多态性位点百分比、有效等位基因数、Nei’s 基因多样性指数和Shannon’s 信息指数等方面表现均较好。 这100 份小麦种质中存在较丰富的遗传变异,不同种质间的遗传相似系数变幅为0.56 ~0.96,平均为0.73,基于遗传距离可将其划分为6 个类群。 同时,利用多态性比较丰富的5 个SSR 引物,构建了这100 份小麦种质的DNA 指纹图谱。 本研究结果可为供试小麦种质的科学利用和保护提供一定依据。
