实蝇科昆虫对寄主植物的选择和响应研究进展
2023-10-16郭腾达秦誉嘉李志红
郭腾达, 秦誉嘉, 李志红
(中国农业大学植物保护学院植物生物安全系,农业农村部植物检疫性有害生物监测防控重点实验室,北京 100193)
实蝇属双翅目Diptera实蝇科Tephritidae,种类繁多,目前世界已知约500属4 500余种,其中约1 500种实蝇为害植物各种果实,250余种实蝇可直接为害种植类果蔬[1-3]。实蝇科昆虫为植食性昆虫,幼虫为潜食性,可为害寄主植物的各个部位,例如叶、芽、茎、果实、种子及部分寄主植物的花序[4]。尤其是为害寄主果实的实蝇,其成虫产卵于果皮下,幼虫孵化后直接取食果肉,从而导致果实腐烂、早落,造成极大的经济损失,严重时田间果实受害率高达100%[5-6]。
随着全世界物流业快速发展,实蝇的这些特性使其卵和幼虫极易随贸易运输在地区间转移及远距离传播,实蝇对果树、蔬菜等寄主植物的侵害严重威胁果蔬产业持续健康发展[3,7]。实蝇科昆虫较强的入侵能力使其寄主范围也在不断扩大,在世界范围内广泛分布,很多种类已成为世界性检疫害虫,引起世界各国的高度重视[6]。本文对实蝇科昆虫对寄主的选择、响应以及机制的研究进展进行综述, 以期为实蝇科昆虫与寄主植物的相互作用机制和实蝇防控技术的进一步研究提供参考。
1 实蝇对寄主植物的选择
1.1 实蝇的多食性
通常来说,专食性与多食性取决于成虫可以产卵、幼虫可以生长的寄主种类数。植食性昆虫中大约98%的种类是专食性的,仅以一种或少数近缘植物为食;而在实蝇科中,多食性的现象比较常见,尤其在已经公布寄主的果实蝇属的200多种实蝇中,超过40%的种类具有两种或更多寄主植物。重要经济实蝇——橘小实蝇Bactroceradorsalis、地中海实蝇Ceratitiscapitata的寄主种类超过200种[8-11]。为什么在实蝇科中存在较多的多食性种类?通常认为有以下几个原因[10]:
第一,植食性昆虫和它们的寄主植物之间存在共同进化,寄主植物被昆虫取食、产卵时,会产生相应的防御机制,而昆虫为了克服植物的防御也有相应的反防御机制,这样会使得植食性昆虫变得越来越专食[12]。而试验表明,实蝇幼虫取食对寄主植物的种子数量和萌发都没有负面影响,甚至认为果实蝇属幼虫取食对寄主植物有益,推测可能是实蝇幼虫有助于果肉更快地分解,从而增加了种子萌发的数量和速率[13]。多食性实蝇幼虫的发育可能会受到寄主的轻微影响,但在不同的寄主果实中,幼虫存活率仍然很高,所以实蝇对寄主植物专食性的进化需求似乎很少[14]。
第二,大多数植食性昆虫专食性的原因很大程度上是其在复杂的环境中需要准确地对寄主植物进行定位,这也是实蝇科中很多实蝇专食性的主要原因。通常大多数实蝇会优先选择成熟的寄主果实[15],而与成熟有关的是芳香酯的产生,芳香酯是成熟水果的典型气味标志。Cunningham等证明,3种酯(乙酸乙酯、丙酸乙酯、丁酸乙酯)的混合物对昆士兰果实蝇Bactroceratryoni的吸引力与复杂的全水果气味混合物相同或更大,当将酯混合物注射到不太受欢迎的南瓜寄主体内时,对产卵期雌性成虫的吸引力接近于其高度偏好的桃[16]。因此,对许多潜在寄主中常见挥发物的选择适应性使实蝇能够快速定位多个寄主植物。
第三,天敌也是影响实蝇多食性的一个重要原因,由于实蝇的为害特点,很少会受到其他捕食性昆虫的侵害,而寄生蜂是实蝇最重要的天敌之一[17-18]。寄生蜂可以寄生实蝇的卵、幼虫和蛹,依赖植物源气味寄生蜂会精准定位实蝇经常为害的植物,即便是在生理上不合适寄生的幼虫,寄生蜂仍会定位果实并试图寄生,而实蝇将卵产在寄生蜂不能定位的寄主上时,幼虫就可能因避开了寄生蜂而存活下来[19]。随着时间的推移,寄生蜂可能会识别并定位于实蝇的新寄主植物[20]。这可能会导致实蝇再次重新选择寄主,寄主范围也因此扩大。
1.2 实蝇的寄主选择
实蝇通常将卵产在完好的果实中,幼虫蛀食为害。幼虫的发育受到单个寄主果实的限制,因此实蝇对寄主植物的选择是由可以自由飞行和主动寻找寄主的雌成虫决定的,雌成虫对寄主植物的选择会影响其本身以及后代的生长发育[21]。当实蝇进入新的地区后,其对当地寄主植物的选择能力也是该种实蝇能否成功入侵的关键[1]。实蝇科昆虫对寄主植物的选择主要体现在以下几个方面:
1.2.1对不同种类寄主植物的选择
已发现多种实蝇的寄主非常广泛,但是多食性实蝇对不同寄主植物的选择具有明显差异。橘小实蝇在不同水果上的产卵量由多到少依次为:柑橘、香蕉、杧果、枣、石榴、苹果、桃子、梨、李、葡萄、柿,在不同蔬菜上的产卵量由多到少依次为:丝瓜、豇豆、苦瓜、黄瓜、茄子、南瓜、青椒、番茄[22]。番石榴果实蝇Bactroceracorrecta对7种供试水果的选择性由大到小依次为番石榴、杧果、杨桃、香蕉、梨、苹果、柑橘[23]。
李磊等采用绝对寄主适应度和相对寄主适应度模型评估了瓜实蝇Bactroceracucurbitae对39种寄主的适应性表现[24],结果表明,瓜实蝇可以在所有供试寄主上产卵,但其幼虫不能在苹果、牛油果、佛手瓜以及百香果上完成发育。对于能完成世代发育的35种寄主可分为两类,其中笋瓜、葫芦瓜、南瓜、黄瓜、香瓜为瓜实蝇嗜好寄主,其他30种为普通寄主。其他研究[25-29]中实蝇对不同寄主的选择见表1。

表1 不同实蝇对寄主产卵偏好Table 1 Egg-laying preferences of different fruit flies for hosts
1.2.2对寄主植物不同部位的选择
不同实蝇会选择寄主植物的不同部位产卵。大部分实蝇会选择寄主植物的果实,例如橘小实蝇、番石榴果实蝇、瓜实蝇等会在多种水果和蔬菜的果实中产卵,枣实蝇Carpomyavesuviana多选择将卵产于枣果背阴面和枣果底部[30-33]。也有少数实蝇选择寄主植物非果实部位产卵,泽兰实蝇Procecidocharesutilis为专性寄生昆虫,雌成虫将卵产在紫茎泽兰生长点部位,初孵幼虫蛀入到紫茎泽兰植株幼嫩部位取食[34]。荚果实蝇Acanthiophilushelianthi将卵产于花蕾中,并在花蕾中度过从卵到成虫的整个生命周期[35]。防风草实蝇Euleiafratria雌成虫将产卵器穿过寄主植物叶片表面进行产卵[36]。芹菜实蝇Euleiaheraclei通过产卵器在芹菜等寄主植物叶片的下表面产卵,幼虫孵化后开始啃食叶片组织[37]。芦笋实蝇Plioreoceptapoeciloptera雌成虫于4月到6月将卵产在土中新生的芦笋叶中,孵化的幼虫会取食叶片,导致光合作用降低[38]。
1.2.3对寄主植物不同品种的选择
对于同种寄主的不同品种,实蝇的选择也是不同的。橘小实蝇寄主植物广泛,在对杧果品种选择试验中其对‘红象牙杧’和‘三年杧’的选择率远远大于‘金凤凰杧’‘胭脂杧’‘台农1号’;在对5个火龙果品种的选择试验中其偏好在‘普红’‘白肉桂热心’和‘台湾大红’上产卵;而在不同品种的苹果上其产卵量从多到少为‘金帅’‘富士’‘新红星’[39-41]。
杧果也是番石榴果实蝇的嗜食寄主,在对5个杧果品种的选择试验中,番石榴果实蝇选择率由高到低为‘三年杧’‘台农1号’‘虎豹牙’‘胭脂杧’‘鹰嘴杧’[42]。枣实蝇对几种枣果的产卵选择性表现为偏好‘相枣’‘灰枣’‘梨枣’,对‘鸡心枣’和‘骏枣’选择性较差[32]。通过对没有采取防控措施的受害柑橘园调查发现,柑橘大实蝇Bactroceraminax为害不同柑橘品种引起的虫果率由高至低为‘脐橙’‘酸橙’>‘冰糖橙’‘天草’‘蜜橘’>‘沙田柚’>‘椪柑’[43]。
2 实蝇对寄主植物的响应
2.1 实蝇对寄主的生长发育响应
通常实蝇雌成虫会倾向于选择完好的植物果实产卵,完好的果实有利于其后代的生长发育。而由于不同寄主果实在营养成分、化学物质等方面存在较大差异,实蝇在不同的寄主上产卵会造成其后代生活史等方面发育的差异。
墨西哥按实蝇Anastrephaludens在柚、橙、杧果、桃4种水果上卵期分别为3.8、4.3、4.6、3.8 d,幼虫期分别为27.4、29.7、22.3、70.3 d,蛹期分别为17.1、13.7、14.2、13.0 d,在桃上产卵量大于其他3种水果[44]。枣实蝇在‘骏枣’上的蛹期、雌虫怀卵量、性成熟历期、交配时间以及雌雄成虫寿命均略大于‘酸枣’[45]。瓜实蝇和南亚果实蝇在不同寄主(黄瓜、丝瓜、西葫芦)上的蛹历期和羽化率明显不同[46]。
同种寄主不同品种也会影响实蝇的发育参数,周宇翔等选择3个杨梅品种研究橘小实蝇取食后生长发育相关的性状,结果表明,幼虫历期以取食‘荸荠种’时最短,而蛹历期和蛹重以取食‘水梅’时最短和最轻;雄成虫以取食‘东魁’时最重,幼虫、幼虫-蛹、幼虫-成虫的存活率以取食‘荸荠种’时最高,‘水梅’时最低[47]。
寄主的营养成分是影响实蝇寄主选择的一个重要因素,碳水化合物、脂肪、蛋白质等重要物质的种类和比例对实蝇的生长发育和生殖有直接的影响[48-49]。Hafsi等研究了7种实蝇幼虫在22种寄主植物上的表现,结果表明,不同寄主对幼虫存活率、发育时间和蛹重均有显著影响,且在高存活率果实上饲养的幼虫具有较高的蛹重和较短的发育时间;在碳水化合物、纤维和脂肪含量较高的果实中,多食性实蝇种类的存活率高于寡食性种类[50]。橘小实蝇幼虫的发育参数(长度、干重和发育历期)会随寄主果实适宜性的增加而增加,取食香蕉、杧果和桃的橘小实蝇幼虫三项参数均最佳,而取食苹果和番茄时幼虫体长最短和干重最轻,并且世代发育时间延长,这些结果也表明寄主果实质量是决定幼虫发育能否完成及发育历期的关键因素[51]。
2.2 实蝇对寄主的行为响应
实蝇依靠视觉、触觉等器官对植物的物理、生理等特征和性状作出判断,以决定是否将其作为适宜的产卵寄主,依靠嗅觉系统对寄主散发的气味进行选择。不同寄主由于颜色、形态、气味不同会导致其对实蝇的吸引力有显著差异[52]。
寄主植物的颜色和形状会影响实蝇成虫的选择。室内试验发现,橘小实蝇雌虫对颜色的选择依次为橘黄色、绿色、黄色、蓝色、黑色、红色、紫色、白色,对形状的选择依次为球形、半球形、圆柱形、三角锥形、长方形、方形[22]。Wu等通过色纸测试橘小实蝇颜色偏好,结果表明,紫外线(300~380 nm)和绿色(500~570 nm)刺激会增强彩色纸对橘小实蝇的吸引力,而蓝色(380~500 nm)刺激会降低吸引力[53]。柑橘大实蝇成虫对黄色和橙色基质的食物趋性最强,其次为绿色,对白、蓝、黑、红色基质的食物趋性较弱[54]。
寄主植物中的化合物会影响幼虫的生长发育,幼虫的发育通常会影响成虫对寄主植物的选择。幼虫发育与寄主植物中的化合物、营养成分之间的关系对实蝇寄主范围起着决定性作用,Varikou等连续两年利用4个橄榄品种评估了油橄榄实蝇Bactroceraoleae与果实酚类化合物浓度以及果实大小的关系。果肉中橄榄苦苷、总酚类化合物和邻二酚类化合物的浓度与实蝇对品种的敏感性呈负相关关系,实蝇倾向于选择酚类化合物浓度较低且果实较大的品种[55]。
实蝇还会通过杂交和寄主转移现象提高其适应性,Schwarz等研究表明白浆果实蝇Rhagoletiszephyria和蓝莓实蝇Rhagoletismendax可以通过杂交产生杂交种,其杂交种出现寄主转移现象,即该杂交种不选择亲本的寄主植物作为寄主,而选择其他植物作为为害对象,该现象可以帮助杂交种摆脱与亲本的寄主竞争,该结果对进一步研究实蝇对适应寄主的行为响应及寄主选择具有重要意义[56]。
2.3 实蝇对寄主挥发物的响应
昆虫依靠识别环境中的气味分子来做出响应,气味分子主要包括植物产生的挥发物以及昆虫产生的信息素等[57]。植物挥发物是由植物自身合成释放的化合物,由挥发性有机化合物组成,在常温下通常是低分子量、易气化的亲脂性液体,主要包括醇类、烃类、酮类、有机酸、有机硫、含氮化合物等[58-59]。而昆虫灵敏的嗅觉系统能够很好地识别植物挥发物[60-61]。不同的寄主植物的挥发物在种类、总量等方面上都存在一定的差别,因此昆虫能够对不同的寄主植物进行选择[62]。
柑橘大实蝇成虫对14种板栗挥发物的触角电位(EAG)反应和嗅觉行为反应的研究表明,柠檬烯、α-蒎烯、苯乙酸甲酯、异丁醛、芳樟醇、β-石竹烯和苯乙醛对初羽化柑橘大实蝇成虫具有显著或极显著的引诱作用,戊醛、乙偶姻、乙酸芳樟酯和水杨酸甲酯对柑橘大实蝇成虫表现出显著或极显著的驱避作用,壬醛对柑橘大实蝇雌虫具有极显著的驱避作用、对柑橘大实蝇雄虫具有极显著的引诱作用[63]。‘红江橙’青果挥发物中的柠檬烯、4-萜烯醇和柠檬醛活性成分能引起柑橘大实蝇雄虫的EAG反应,而柑橘大实蝇雌虫仅对柠檬烯和壬醛有EAG反应[64]。
橘小实蝇对不同的寄主挥发物表现出不同的趋性,吴健等测试了8种寄主植物挥发物(β-紫罗兰酮、乙酸异戊酯、罗勒烯、苯乙醇、己酸乙酯、乙酸叶醇酯、乙酸丁酯、苯甲醛)对橘小实蝇的引诱效果,除苯甲醛外,7种挥发物对性成熟的橘小实蝇雌雄成虫都具有显著的引诱作用,且对雌雄虫的引诱效果无明显差异[65]。此外,从3种可以引诱橘小实蝇产卵的寄主果实(番石榴、橙子和杧果)提取共有挥发物β-石竹烯,利用Y型嗅觉仪和四臂嗅觉仪测试均表明低浓度β-石竹烯(9 μg/mL)对橘小实蝇雌成虫具有引诱作用,但高浓度β-石竹烯对橘小实蝇没有引诱作用,增加寄主番石榴果实中β-石竹烯的含量也会显著降低果实对雌虫产卵的诱集效果[66]。
在枣果挥发物中棕榈酸乙酯、月桂酸乙酯、肉豆蔻酸与枣实蝇产卵选择密切相关,在田间试验中,枣实蝇成虫对肉豆蔻酸的选择率较强[67]。具条实蝇Zeugodacusscutellata对寄主植物挥发物3-辛酮的EAG响应显著高于诱蝇酮、覆盆子酮和姜酮;EPG(刺探电位图)响应则相反,对诱蝇酮、覆盆子酮和姜酮的响应显著高于3-辛酮[68]。
3 实蝇对寄主植物的选择和响应机制
3.1 实蝇的感受器官
目前已知实蝇科昆虫常见触角感受器主要有5种:毛形感受器、锥形感受器、腔锥形感受器、刺形感受器、棒状感受器。其他比较少见的感受器包括:钟形感受器、瓶形感受器、柱形感受器、栓锥形感受器、沟槽形感受器等[69]。张国娜运用扫描电镜,描述了6种实蝇(瓜实蝇、黑颜果实蝇Zeugodacusdiaphora、橘小实蝇、柑橘大实蝇、具条实蝇和南亚果实蝇)感受器的类型、形态和分布。在实蝇类昆虫下颚须上共有3种类型的感受器,分别为刺形感受器、微毛形感受器和锥形感受器;跗节上有3种类型感受器:毛形感受器、刺形感受器和锥形感受器;产卵器上共分布有3种类型的感受器,分别为毛形感受器4、毛形感受器5和钟形感受器,这些感觉器官能帮助雌成虫寻找合适的产卵地点[70]。
在双翅目同一科甚至同一属昆虫之间成虫触角感受器类型也存在一定的差异,在橘小实蝇中,成虫触角由柄节、梗节及鞭节组成,鞭节上着生具毛触角芒;触角上有毛形、锥形、刺形和柱形4类9种感受器,其中具弯钩的毛形感受器首次在橘小实蝇成虫触角上被观察到[71-72]。而Awad 等发现桃实蝇Bactrocerazonata则没有上述锥形感受器和柱形感受器,但其具有橘小实蝇触角没有的钟形感受器[73]。不同地理种群的同一种实蝇触角感受器也会存在明显的适应性变化。位于我国台湾的橘小实蝇触角上发现有栓锥形感受器和棒状感受器[74],然而我国大陆的橘小实蝇皆未发现以上两种感受器,但大陆的橘小实蝇触角上发现有刺形感受器、腔锥型感受器或柱形感受器;此外,不同地理种群的橘小实蝇在亚型和部分感受器形态上也存在一定差异[71,75-77]。触角感受器的数量及分布是与生存环境相互作用中自然选择压力等综合作用的结果,与食性、习性和栖境相关[78]。因此可以认为,不同地理种群的橘小实蝇由于其偏好寄主和栖息环境的不同造成了触角感受器的差异。
在其他实蝇中,南亚果实蝇的触角外观形态和所含的感受器类型与橘小实蝇大致相同,包括毛形感受器、柱形感受器、锥形感受器和刺形感受器等[76];枣实蝇触角上有5种感受器类型:鬃形感受器、毛形感受器、锥形感受器、腔锥形感受器和棒状感受器[79];在番石榴果实蝇中化学类感受器主要由毛形感受器、锥形感受器、棒形感受器、沟槽形感受器组成,其表面具孔或者沟槽,挥发性化学分子可以通过[80]。对柑橘大实蝇产卵器的超微结构观察可以看到,柑橘大实蝇产卵器由产卵器基节、翻缩膜和产卵针3部分组成,上有毛形、腔锥形和钟形3种类型感受器[81]。
3.2 实蝇化学感受基因
昆虫有一套复杂而精细的嗅觉感受系统,其利用多种类型的嗅觉感受器与化学感受基因,对环境中的气味分子做出不同的行为与生理反应,例如可进行寄主定位、寻找产卵场所、躲避天敌等[82-83]。近年来昆虫的嗅觉识别过程相关的基因功能特征正在逐步被揭示,包括气味结合蛋白(odorant binding proteins, OBPs)、气味受体(odorant receptors, ORs)、味觉受体(gustatory receptors, GRs)、离子型受体(ionotropic receptors, IRs)、气味降解酶(odorant degrading enzymes, ODEs)、化学感受蛋白(chemosensory proteins, CSPs)和感觉神经元膜蛋白(sensory neuron membrane proteins, SNMPs)等的基因[84-87]。
在实蝇科中,与化学感受相关机制已有大量研究,目前多种实蝇的化学感受基因已被鉴定(表2),有助于了解相关通路中基因的潜在功能。例如,橘小实蝇和番石榴果实蝇中化学感受基因有比较明显的扩张现象,对于入侵物种的种群建立和快速传播至关重要[83]。
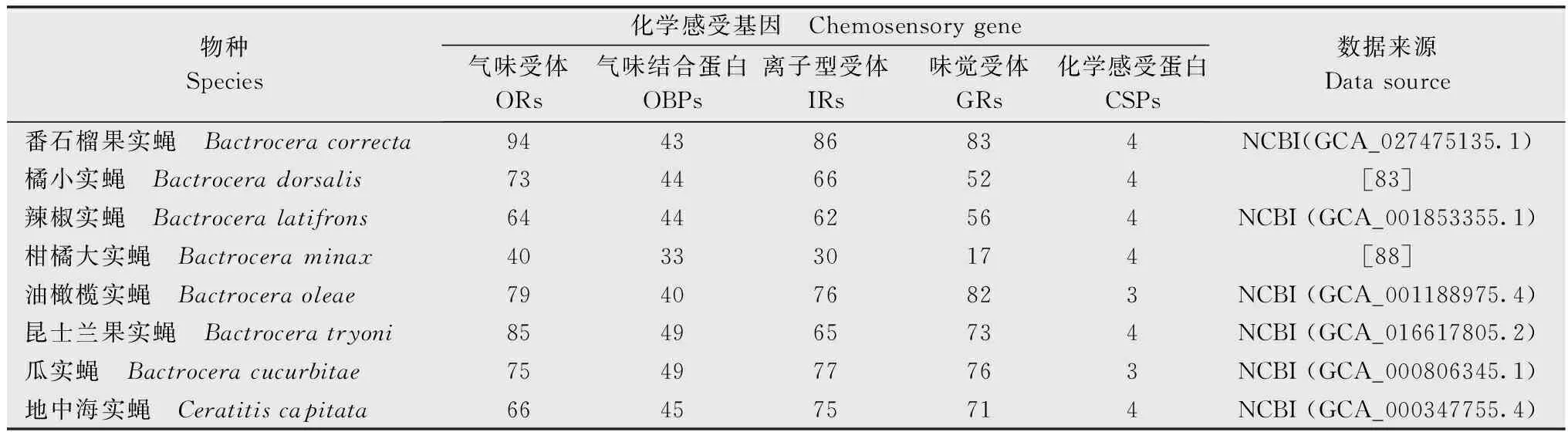
表2 主要实蝇的化学感受基因数量Table 2 The numbers of chemosensory genes in major fruit fly species
对柑橘大实蝇4 个气味受体基因的研究表明4 个 BminORs 都能被一种植物挥发物激活,BminOR3、BminOrco仅对1-辛烯-3-醇的刺激具有强烈的电生理反应;BminOR12、BminOrco则对多种植物挥发物的刺激具有电生理反应[89]。柑橘大实蝇BminOR24在雌、雄成虫触角中高表达,当与嗅觉受体共受体BminOrco在非洲爪蟾卵母细胞中共表达时,细胞对芳樟醇反应强烈[90]。此外,柑橘大实蝇气味结合蛋白BminOBP9可以和11种柑橘挥发物进行结合[91],化学感受蛋白基因BminCSP和气味结合蛋白基因BminOBP21参与调控柑橘大实蝇感受柑橘寄主挥发物D-柠檬烯[92]。
BdorOBP83b 和 BdorOBP56f-2是橘小实蝇中感知甲基丁香酚的气味结合蛋白[93-94]。BdorOBP13仅在橘小实蝇雄性和雌性成虫中表达,在触角中表达量较高,RNAi结果显示,注射BdorOBP13 dsRNA后,橘小实蝇对甲基丁香酚(雄性引诱物)和丙位辛内酯(雌性引诱物)的电生理反应显著降低,表明该基因可能参与了雄性实蝇对甲基丁香酚的感知和雌性实蝇对丙位辛内酯的感知[95]。甲基丁香酚诱导的表达谱和体外结合测定显示 BdorOBP69a 也可以参与对甲基丁香酚的感知,并以强亲和力直接与其结合[96]。橘小实蝇BdorOrco也参与调控性成熟雄虫对植物源诱剂甲基丁香酚的感知过程[97],BdorOR82a和BdorOR13a分别参与调节橘小实蝇雌虫对植物挥发物乙酸香叶酯和1-辛烯-3-醇的识别,促进其交配后定位产卵寄主[98]。
3.3 实蝇抗性相关基因
植物在与昆虫共存的过程中表现出非常复杂的互作关系,为抵御害虫的为害,许多植物已进化出多种防御机制,例如改变植物形态、合成次生代谢物、释放趋避性挥发物等[99-101]。为了对抗植物次生代谢物,昆虫进化出一套完整的解毒系统,包括重要的解毒酶(细胞色素P450、谷胱甘肽S-转移酶等)和功能基因家族(UDP-葡萄糖醛酸转移酶和ABC转运蛋白)来抵御各种毒素的影响[102-104]。
实蝇科强大的适应能力来自其具有复杂的解毒系统。研究表明,解毒基因的数量与寄主的适应有关[105](表3)。橘小实蝇和番石榴果实蝇的入侵扩张可能与P450和ABC转运蛋白家族的扩张相关,使其在极端环境中具有较强的抗性和生存能力。

表3 主要实蝇的抗性相关基因数量Table 3 The numbers of resistance-related genes in major fruit fly species
Zhang等通过对植物代谢组学和昆虫转录组学的联合分析,探讨了未成熟柑橘果实与柑橘大实蝇的相互作用[106]。在未成熟柑橘果实代谢组中鉴定出18种次生代谢产物,主要包括黄酮类、生物碱类和苯丙素类,它们会在幼虫取食过程中积累。与成熟柑橘相比,取食未成熟柑橘果实幼虫的体内有3种与解毒相关的基因(1个P450基因,2个 ABC转运蛋白基因)高表达。分别利用不同次生代谢产物处理幼虫,会使一种ABC转运蛋白基因显著上调,利用RNAi 技术使该 ABC转运蛋白基因沉默后,幼虫死亡率达到51%。
通过比较泽兰实蝇对原产地和入侵地紫茎泽兰植株的选择,寄生在入侵地紫茎泽兰上的泽兰实蝇的羧酸酯酶活性低于原产地,但谷胱甘肽S-转移酶在雌虫中的活性则相反,这个结果可能是由于原产地与入侵地紫茎泽兰中次生物质的区别导致的[107]。
4 展望
实蝇作为一种主要为害果实的害虫,与寄主植物之间的复杂相互作用一直是研究者关注的焦点。一方面,实蝇能够通过感知和评估寄主植物特征来选择和定位寄主,这一过程涉及多个感觉器官的协同作用,包括视觉、嗅觉和触觉等[40, 63, 108]。另一方面,寄主植物可以启动一系列防御反应来抵御实蝇的为害,这些防御反应可以通过激活抗虫化学物质的合成和积累等方式干扰实蝇幼虫取食和成虫产卵行为,从而减少实蝇的为害[109-110]。最后,实蝇也能通过逃避、解毒等方式适应或降低寄主植物的防御反应[106](图1)。
近几十年来,随着技术的不断发展,在分子生物学层面上研究人员已经开始探索寄主植物与实蝇之间的相互作用,例如通过分析寄主植物做出防御反应时代谢物的变化,可以了解植物对实蝇入侵的应答机制[106]。基因组学、蛋白组学、代谢组学等研究手段也为揭示实蝇对寄主植物选择和响应的分子机制提供了重要途径。越来越多的实蝇基因组数据的公布和基因功能研究的发表,为多组学联合分析比较实蝇对不同寄主植物的响应如感受器官的发育和功能等研究奠定了基础。电生理技术被广泛应用于实蝇的寄主选择和行为响应研究,通过记录实蝇感觉器官的电生理反应,可以揭示实蝇对寄主植物刺激的感知和响应过程,更好地了解实蝇响应寄主植物的机制。
尽管实蝇与寄主植物互作研究已经取得了一定进展,但仍有许多问题需要进一步解决。首先,在实蝇与寄主的分子互作机制研究中,目前的研究主要集中在特定基因的鉴定与功能研究,还缺乏对多种基因协同作用的机制研究等。其次,对于实蝇选择寄主的重要性状,如对寄主外形特征的识别和挥发性化合物的感知等,仍需进行更深入的研究。此外,实蝇产卵方式和幼虫取食会诱导寄主植物产生防御反应,不同寄主植物的抗性机制对实蝇产卵和幼虫发育的影响还需要进一步阐明。在实际应用方面,当前针对实蝇的防治措施主要是基于化学农药,但其效果有限且存在环境风险。因此,基于实蝇与寄主的互作关系研发安全、协调和高效的实蝇防控新技术仍然是一个迫切的需求。
综上,笔者对实蝇及其寄主选择与响应相关内容进行了综述,以期提高对实蝇与寄主相互作用的理解。实蝇与寄主植物之间的研究仍有许多待探索的领域,通过深入研究实蝇的感知机制、寄主植物的防御反应和共进化关系,可以为开发更稳定、高效的实蝇防治措施提供理论依据,降低实蝇对农作物产量和品质的损害,促进农业的可持续发展。
