施氮量对滴灌春小麦茎鞘NSC积累与转运的影响
2023-10-10车子强蒋桂英王海琪王荣荣尹豪杰
车子强,蒋桂英,王海琪,王荣荣,尹豪杰
(石河子大学农学院,新疆 石河子 832000)
促进光合产物向籽粒转运是禾谷类作物高效生产的关键,阐明光合产物向籽粒转运,特别是花前和花后临时贮藏在营养器官的光合产物向籽粒转运的调控机制,对解决目前小麦生产上存在的高施氮水平下茎鞘中光合产物向籽粒转运率低、籽粒灌浆慢等问题具有十分重要的理论和实践意义[1]。小麦开花前和开花后,在茎鞘临时贮存并可转运的光合产物称为非结构碳水化合物(Nonstructural carbohydrate,NSC),其主要成分是果聚糖,含量占茎杆总干质量的40%以上,果聚糖的代谢调控茎鞘NSC积累和再运转[2],对于缓和植株源叶片光合产物供应与籽粒库光合产物需求之间的矛盾,维持较高的籽粒灌浆速率具有重要作用[3]。
施用氮肥是协调作物源-库关系、影响作物产量形成的重要栽培措施。合理的氮肥供给有利于促进小麦籽粒灌浆中后期茎鞘中积累的NSC向籽粒输出,给籽粒灌浆提供所需营养,影响其粒重和产量[4]。研究发现,开花前小麦茎鞘中贮藏物质对产量的贡献率为3%~30%,开花后小麦茎鞘中的暂贮物对产量的贡献占籽粒干物质的10%~25%[5]。潘俊峰等[6]研究发现,长江中下游麦区低氮处理(50 kg·hm-2)增加了茎鞘花前NSC的积累以及花后NSC向籽粒的转运;Liang等[7]研究发现,在中国北部冬麦区,适宜的施氮量(180 kg·hm-2)显著提高了花前茎鞘中NSC含量,促进弱势粒库强和灌浆速率,而过量施氮显著降低了花前茎鞘NSC积累量,抑制了小麦弱势粒库强和灌浆速率。茎鞘中存在着聚合度(Degree of polymerization, DP)不同的果聚糖[8-9],其中DP≥4的果聚糖聚合度较高,其含量与聚合程度可反映营养器官中光合产物积累与贮藏的能力[10]。当植株茎鞘干物质量降低时,蔗糖果糖基转移酶(SST)催化蔗糖合成果聚糖(DP=3),此为果聚糖合成的第一步[11],同时也是控制碳素向果聚糖库分配的关键[12],果聚糖果糖基转移酶(FFT)则通过使果聚糖链长增加,进一步催化DP≥4的果聚糖合成,其后在果聚糖外水解酶(FEH)催化下分解转运至作物籽粒[13]。姜东等[11]在华北麦区发现,当施氮量介于210~330 kg·hm-2时,小麦茎鞘中SST活性随施氮量的增加而增大,有利于茎鞘中DP≥4果聚糖含量的积累,但FEH酶活性受到抑制,使成熟期小麦茎鞘中果聚糖分解与转运受阻。Guo等[14]研究也发现施氮量过多会导致茎鞘中果聚糖积累与转运受到抑制,而适宜的氮肥施用量可促使茎鞘中果聚糖的积累及转运。可见,施氮量可不同程度调控茎鞘中NSC和果聚糖代谢及籽粒产量形成。
滴灌是新疆麦区的主要灌溉技术,因施肥和灌水方式不同,滴灌麦区的田间气候等条件与其他麦区存在差异,光合产物的积累与转运也明显不同[15],且新疆小麦生产中长期追求高肥高产,氮肥施用量常年保持在300 kg·hm-2,氮肥的持续高投入不仅使种植成本增加,还造成了环境污染[16-17]。在新疆田间滴灌条件下,如何协调施氮量、果聚糖、NSC以及小麦产量之间的关系,促进茎鞘中果聚糖和NSC向小麦籽粒高效转运以提高小麦产量仍未有定论。因此,本试验以当地主栽小麦品种为材料,通过设置不同施氮水平,利用HPLC技术,探究滴灌春小麦茎鞘中DP≥4果聚糖和NSC的代谢特征,分析花后小麦茎鞘中DP≥4果聚糖和NSC积累与转运及其对籽粒产量贡献的动态规律,以期为新疆滴灌小麦氮肥优化施用提供科学依据。
1 材料与方法
1.1 试验区概况
试验于2021年4—7月在新疆石河子大学农学院实验站(85°59′E,44°18′N)进行,该地区多年平均气温为7.5~8.2℃,多年平均降水量208 mm,多年平均蒸发量1 660 mm,属于典型的大陆性气候。2021年4—7月小麦生育期间平均日最高气温、日最低气温和逐日降水量变化如图1所示。供试土壤类型为灌溉灰漠土,供试0~40 cm土层土壤基本性状如表1。

表1 供试土壤基本理化性质Table 1 Physical and chemical characteristics of the experimental soil
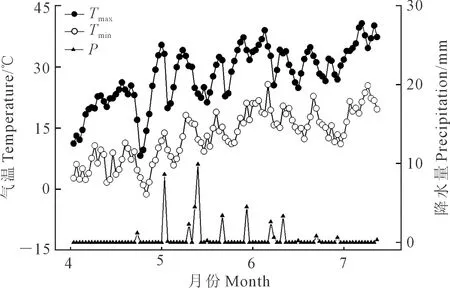
图1 小麦生育期平均日最高气温(Tmax)、平均日最低气温(Tmin)和逐日降水量(P)变化Fig.1 Changes of mean daily maximum temperature (Tmax),mean daily minimum temperature (Tmin) and daily precipitation (P) during wheat growth period
1.2 试验设计
供试小麦品种为强筋型‘新春37号’(XC37,蛋白质含量16.3%)和中筋型‘新春6号’(XC6,蛋白质含量13.5%)。试验采用裂区设计,品种为主区,氮素为副区,设置4种氮肥处理:CK1(300 kg·hm-2)、A1(255 kg·hm-2)、B1(210 kg·hm-2)、CK2(0 kg·hm-2),各处理具体氮肥施用量如表2所示。
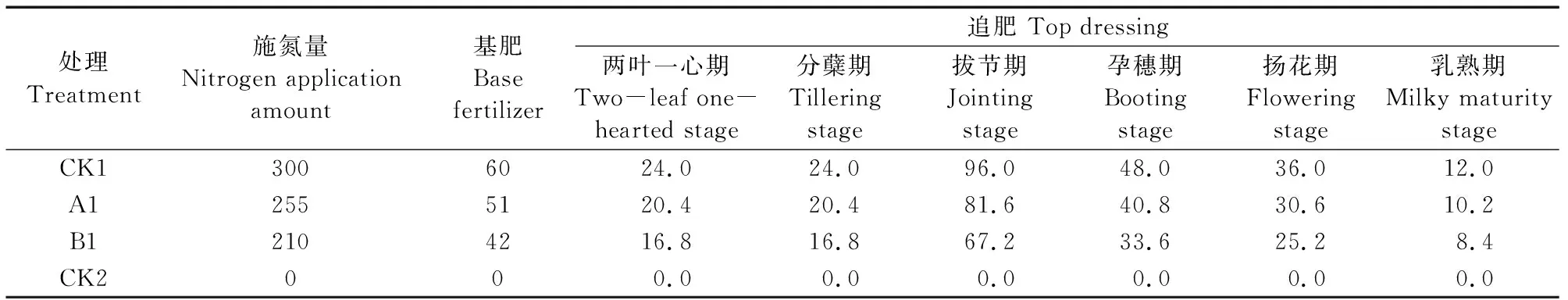
表2 不同处理氮肥施用量/(kg·hm-2)Table 2 Nitrogen fertilizer application amount of different treatments
小区种植面积为12 m2(3 m×4 m),各小区间埋置100 cm深度的防渗膜,防止肥料外渗。氮肥基追比例为2∶8,播前各小区将20%氮肥(尿素,N 46%)和120 kg·hm-2的P2O5(过磷酸钙,P2O512%)作基肥翻耕于土壤;其余氮肥按不同比例在各生育期随滴灌水施入土壤。
生育期间灌溉量为6 000 m3·hm-2,共灌水9次,各个时期的灌水量通过水表精确控制。小麦于2021年4月4日播种,播种量为345 kg·hm-2,采用“宽窄行”、“一管4行”的种植方式,行间距为12.5 cm+20 cm+12.5 cm+15 cm,滴灌带(管径16 mm,滴头间距30 cm,流量2.6 L·h-1)放置在20 cm的宽行;2021年7月7日收获,其他田间管理同大田生产。
1.3 测定项目与方法
1.3.1 茎鞘果聚糖代谢酶活性的测定 于开花期、乳熟期、蜡熟期和成熟期取同一日开花的小麦,每个处理选取5株,将茎鞘按穗下节间、倒二节间和其余节间3个节位分样,鲜样置入液氮,-80℃下保存,用于测定果聚糖代谢酶活性。蔗糖果糖基转移酶(SST)和果聚糖外水解酶(FEH)活性测定参照Verspreet等[18]的方法。
1.3.2 茎鞘果聚糖(DP≥4)和NSC含量的测定及计算 取样时间同1.3.1小节。每个处理选取10株单茎,分别剪取茎鞘倒二节间和其余节间,5株用于测定果聚糖(DP≥4)含量,另外5株用于测定NSC含量。果聚糖(DP≥4)和NSC含量使用高效液相色谱法(HPLC)测定,具体参照姜东等[19]的方法。测定使用安捷伦1200系列液相色谱仪,色谱柱为艾杰尔Bonshell ASB C18(150 mm×2.1 mm,1.7 μm),以0.1%三乙胺-乙腈溶液进行梯度洗脱流动相,柱温为28℃,流速为1 mL·min-1,漂移管温度为75℃,气体压力为40 psi,喷雾器模式加热,动力水平为60%。参照马召朋等[20]的方法计算果聚糖以及NSC花前、花后转运、贡献率和绝对含量,果聚糖各指标计算方法如下:
花前转运率(%)=(花后当天果聚糖绝对含量-成熟期果聚糖绝对含量)/花后当天果聚糖绝对含量×100%
花后转运率(%)=(最大果聚糖绝对含量-花后当天果聚糖绝对含量)/最大果聚糖绝对含量×100%
花前贡献率(%)=(花后当天果聚糖绝对含量-成熟期果聚糖绝对含量)/(1000×主穗粒重)×100%
花后贡献率(%)=(最大果聚糖绝对含量-花后当天果聚糖绝对含量)/(1000×主穗粒重)×100%
绝对含量(mg·g-1)=果聚糖浓度×干物质量
NSC花前和花后转运率及其贡献率计算公式同果聚糖。
1.3.3 茎鞘干物质量测定及相关指标的计算 于小麦开花期和成熟期分别取具有代表性的10株小麦,按照穗下节、倒二节和其余节3个节位分别置于105℃烘箱杀青30 min,70℃烘干至恒重后称重,即为茎鞘不同节位干物质量。茎鞘中干物质的积累与转运及其对小麦产量的贡献率按照王茂莹等[21]的方法进行计算。
花前贮藏物质运转量(kg·hm-2)=开花期干质量-成熟期干质量
花前干物质转移效率(%)=花前贮藏物质运转量/开花期干质量×100%
花前物质运转贡献率(%)=花前贮藏物质运转量/成熟期籽粒干质量×100%
花后物质积累量(kg·hm-2)=成熟期籽粒干质量-花前贮藏物质运转量
花后干物质转移效率(%)=花后贮藏物质运转量/开花期干质量×100%
花后积累贡献率(%)=花后物质积累量/成熟期籽粒干质量×100%
1.3.4 产量的测定 于小麦成熟期每个处理小区随机选取1 m2的样方,将植株全部收割后自然晾干,称籽粒重量,计算产量;同时测定1 m2样方内小麦穗数,从中随机选取20株测定穗粒数和千粒重,重复3次。
1.4 数据分析
使用Microsoft Excel 2016和Origin 2018进行数据处理和图表绘制,利用SPSS进行统计分析,运用Duncan及LSD方法进行显著性检验。
2 结果与分析
2.1 不同施氮处理下春小麦茎鞘果聚糖代谢关键酶活性的变化
由图2可知,不同施氮处理下两个品种不同茎鞘节位SST活性均随生育进程推进呈先升后降变化趋势,乳熟期最大。节位、施氮量和品种不同,SST活性变化也不同。各品种相同施氮量下,A1处理其余节SST活性最高,分别比倒二节、穗下节高15.02%~53.44%、52.11%~106.26%。各品种相同节位下,除开花期和蜡熟期外,其余生育时期各施氮处理SST活性均显著高于CK2,表现为A1>CK1>B1>CK2;各生育时期A1处理其余节SST活性均显著高于CK1、B1、CK2处理,增幅分别为5.59%~10.13%、8.97%~13.16%、20.48%~30.24%。同一节位不同品种之间,A1处理其余节中SST活性XC37比XC6高2.20%~9.67%。综上可知施氮量能显著影响小麦各节位SST活性变化,影响效果因品种、节位和生育时期而异;两个品种(XC37和XC6)SST活性均为A1处理(施氮量为255 kg·hm-2)在整个生育期综合表现较优。
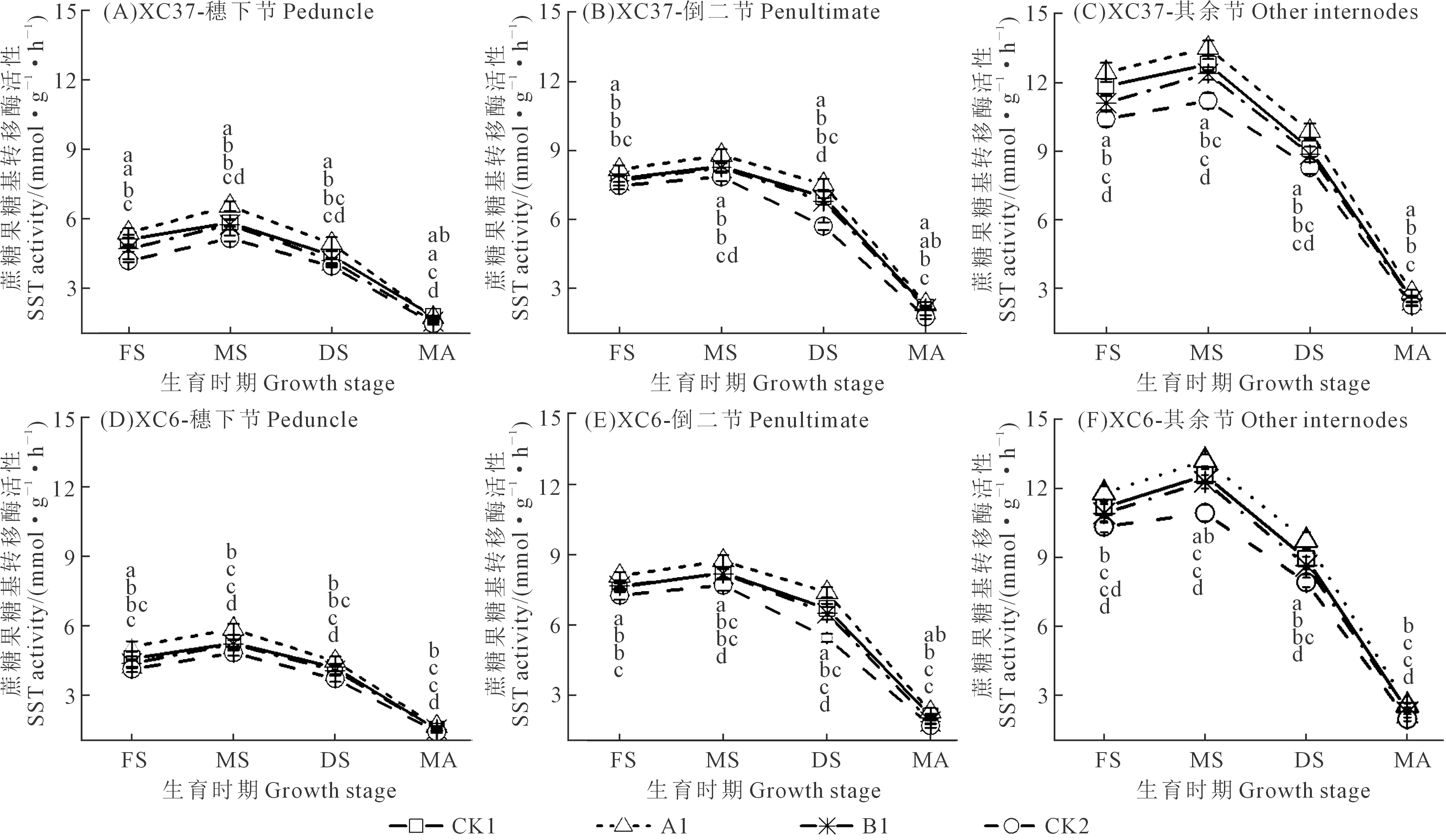
注:FS:开花期;MS:乳熟期;DS:蜡熟期,MA:成熟期。图中不同小写字母代表两品种在同一时期不同处理间差异显著(P<0.05)。下同。Note:FS: Flowering stage; MS: Milky maturity stage; DS: Dough stage; MA: Maturity stage. Different lowercase letters represent significant differences (P<0.05) between two varieties in different treatments at the same time. The same as below.图2 施氮处理对滴灌春小麦不同节位SST活性的影响Fig.2 Effects of different nitrogen application on SST activity at different nodes of spring wheat under drip irrigation
由图3可知,不同施氮量下,两个品种各茎鞘节位的FEH活性均呈“谷-峰-谷”的变化趋势,乳熟期、成熟期低谷,蜡熟期达峰值。同一生育时期各节位FEH活性随施氮量降低呈先升后降变化,A1处理在蜡熟期可保持较高的果聚糖分解能力(FEH活性为26.43~85.69 mmol·g-1·h-1),CK1处理则使FEH活性显著降低(20.57~77.86 mmol·g-1·h-1)。节位、施氮量和品种不同,FEH活性变化也有所不同。各品种相同施氮量下,A1处理FEH活性在其余节最高,分别比倒二节和穗下节提高11.38%~29.78%和37.87%~57.39%。各品种相同节位下,除开花期外,A1处理其余节FEH活性均显著高于CK1、B1、CK2处理,增幅分别为10.06%~21.50%、13.61%~30.87%、20.96%~44.84%。同一节位相同施氮量下,XC37品种A1处理其余节FEH活性比XC6高2.67%~5.88%。综上可知,两个品种FEH活性均为A1处理在整个生育期综合表现较优。
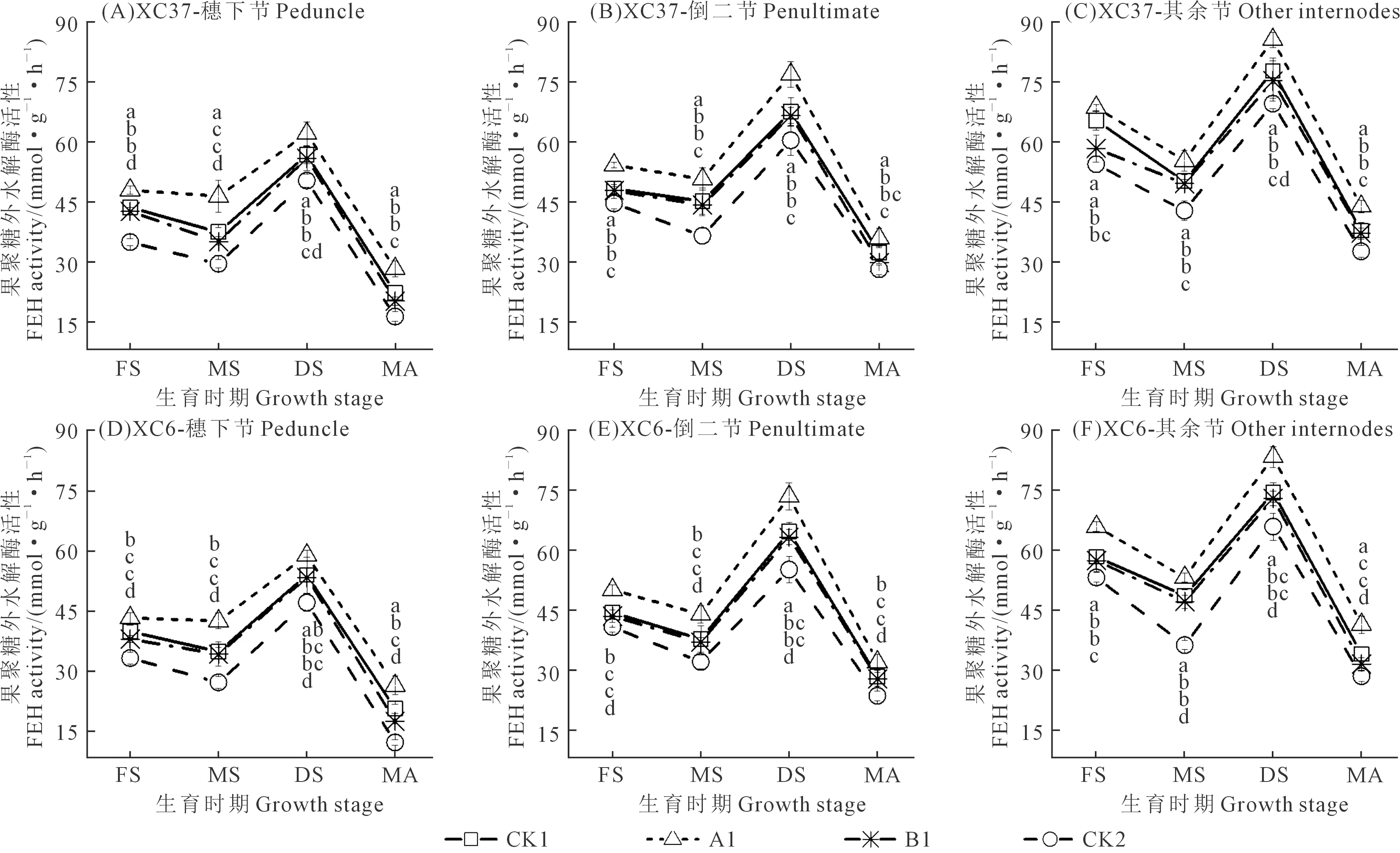
图3 施氮处理对滴灌春小麦不同节位FEH活性的影响Fig.3 Effects of different nitrogen application on FEH activity at different nodes of spring wheat under drip irrigation
2.2 不同施氮处理下春小麦茎鞘果聚糖含量和NSC含量的变化
不同施氮量下,倒二节和其余节中DP≥4果聚糖含量(图4)和NSC含量(图5)均随施氮量的增加先增后降,表现为A1>CK1>B1>CK2(乳熟期除外),说明A1处理可有效提高茎鞘中果聚糖供应,CK1处理则导致果聚糖供应水平降低。节位、施氮量和品种不同,果聚糖和NSC含量变化也有所不同。
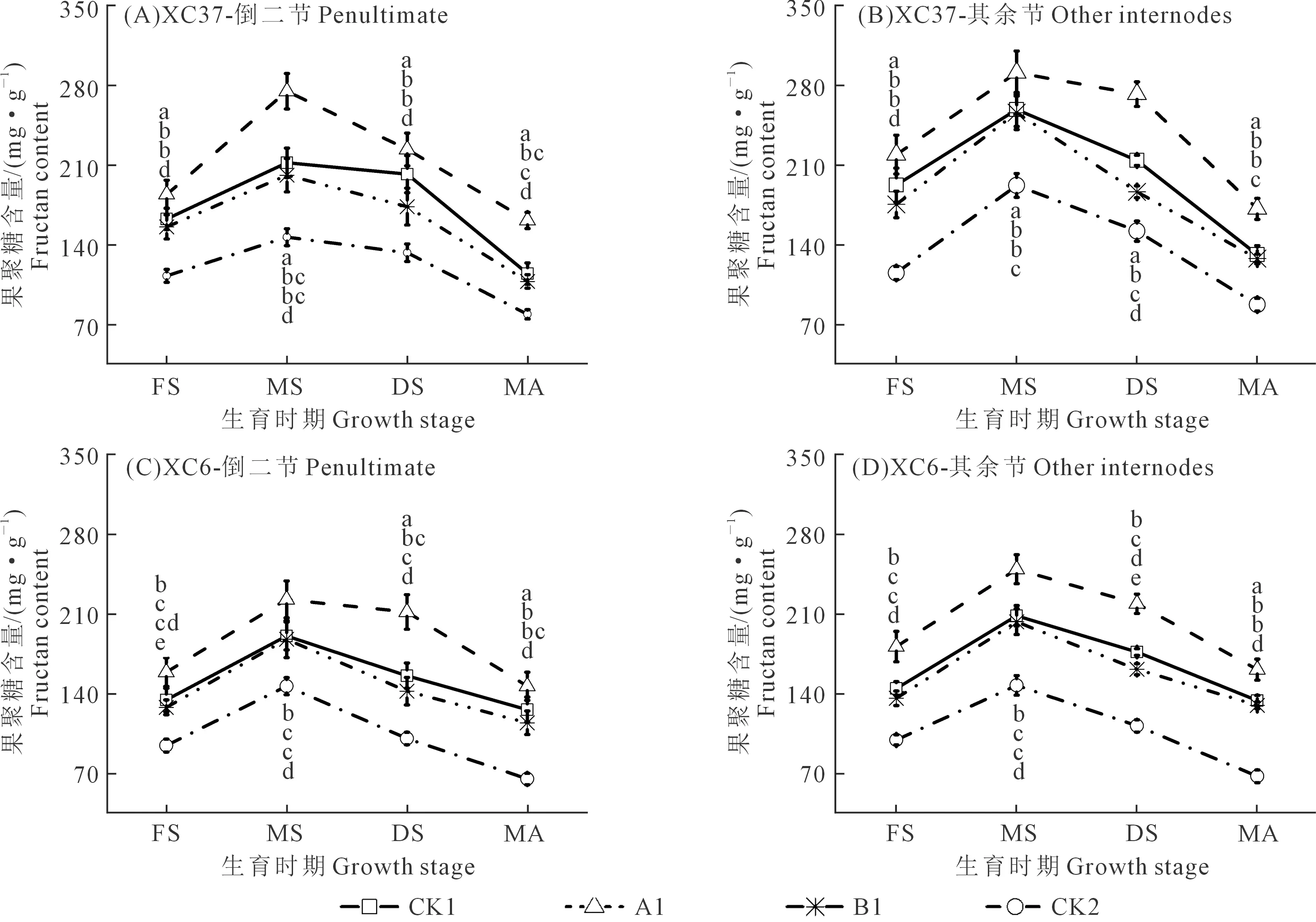
图4 施氮处理对滴灌春小麦不同节位果聚糖含量(DP≥4)的影响Fig.4 Effects of different nitrogen application on fructan content (DP≥4) at different nodes of spring wheat under drip irrigation
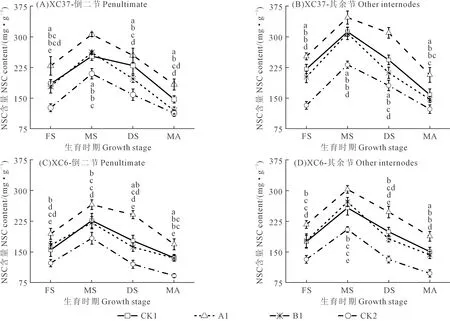
图5 施氮处理对滴灌春小麦不同节位NSC含量的影响Fig.5 Effects of different nitrogen application on NSC content at different nodes of spring wheat under drip irrigation
如图4所示,茎鞘中DP≥4果聚糖含量随生育时期推进呈先增后降的倒“V”趋势,乳熟期最高,成熟期最低。各品种相同施氮量下,A1处理开花~成熟期其余节DP≥4果聚糖含量均高于倒二节,增幅为5.89%~9.59%,且在开花~蜡熟期差异明显,成熟期差距逐渐缩小。各品种相同节位下,A1处理其余节中果聚糖含量分别比CK1、B1、CK2显著高12.46%~22.18%、13.74%~25.51%、51.35%~139.14%,倒二节中分别高27.62%~29.61%、35.75%~36.57%、87.13%~124.01%。同一节位相同施氮量下,XC37品种A1处理其余节果聚糖含量比XC6高6.80%~17.01%。
如图5所示,适当降低施氮量可提高茎鞘中NSC供应,过高或过低施氮量则降低NSC含量。各品种相同施氮量下,A1处理各节位NSC变化规律与果聚糖相同,表现为整个生育期其余节高于倒二节,增幅为12.17%~13.45%,开花~蜡熟期两节位差异较大,成熟期差异较小。各品种相同节位下,其余节中A1处理NSC含量分别比CK1、B1、CK2显著高11.12%~24.63%、12.52%~31.49%、49.55%~91.31%,倒二节中分别显著高20.97%~24.24%、16.84%~40.43%、45.55%~82.81%。同一节位相同施氮量下,XC37品种A1处理其余节NSC含量比XC6高9.37%~12.43%。综上可知,两个品种果聚糖和NSC含量均为A1处理在整个生育期综合表现较优。
2.3 不同施氮处理下春小麦茎鞘干物质量及产量的变化
由图6可知,不同施氮量下各节位茎鞘干物质量均随生育进程推进呈先升后降变化,乳熟期最大,此时A1处理各节位茎鞘干物质量均显著高于CK2,穗下节、倒二节、其余节分别高20.34%~23.18%、22.53%~26.50%、46.54%~47.55%。各品种相同施氮量下,A1处理茎鞘干物质量以其余节最高,倒二节其次,穗下节最低,前者较后两者分别高24.41%~44.13%、52.09%~60.57%。各品种相同节位下,除成熟期外,A1处理其余节茎鞘干物质量显著高于CK1、B1、CK2,增幅分别为10.09%~24.31%、12.47%~30.08%、46.54%~66.68%;同一节位相同施氮量下,A1处理其余节茎鞘干物质量XC37比XC6高4.63%~6.32%。综上可知,两个品种在整个生育期茎鞘干物质量均为A1处理综合表现较优。
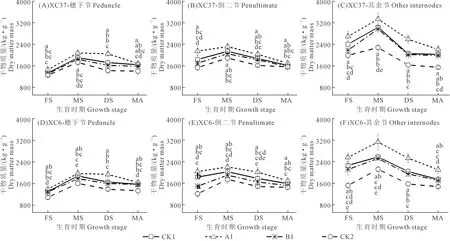
图6 不同施氮处理对滴灌春小麦不同节位干物质量的影响Fig.6 Effects of different nitrogen treatments on dry matter weight at different nodes of spring wheat under drip irrigation
由表3可知,品种对小麦穗数及产量有显著影响,施氮量对小麦产量及其构成因子有极显著影响(P<0.01),施氮量与品种互作对两品种产量及其构成因子影响显著(P<0.05)。千粒重、穗数、穗粒数变化趋势一致,XC37和XC6在A1及CK1处理下产量及其构成因子最大。两品种产量及其构成随施氮量减少呈先增后减的变化,CK2和B1导致产量及其构成因子的降低,CK1增产效果不明显。与CK1相比,XC37在A1处理下千粒重、穗数、穗粒数、产量分别提高3.12%、4.84%、2.06%、3.98%,XC6分别提高3.77%、4.23%、1.93%、6.09%,XC37比XC6分别提高0.89%、1.81%、0.97%、2.97%。经多项式拟合,籽粒产量与氮肥施用量呈二次曲线关系:y21=-0.0168x2+9.4535x+5658.6(XC37,R2=0.9547);y21=-0.0263x2+12.4420x+5376.5(XC6,R2=0.9550)。即当两品种小麦施氮量x为255 kg·hm-2时,XC37的产量为6 976.82 kg·hm-2;XC6的产量为6 839.05 kg·hm-2。
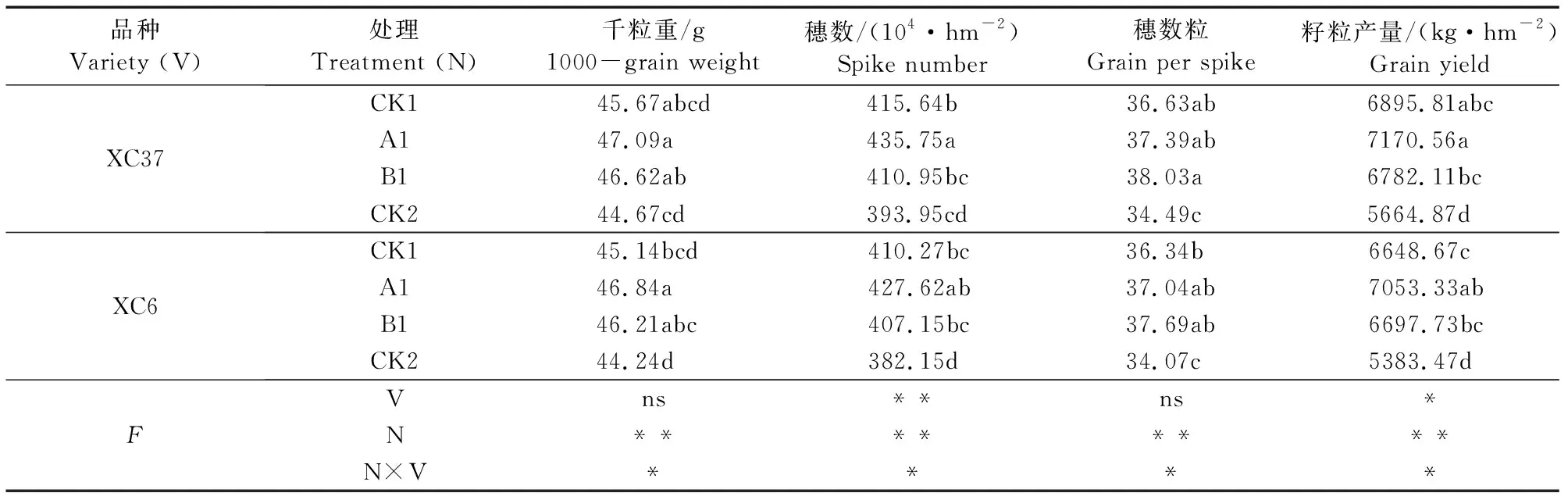
表3 不同施氮处理对滴灌春小麦产量及其构成因子的影响Table 3 Effects of different nitrogen treatments on spring wheat yield and yield composition factors under drip irrigation
2.4 不同施氮处理下春小麦茎鞘果聚糖、NSC、干物质转运对产量的贡献
就果聚糖转运而言,品种对小麦倒二节和其余节的花后转运率无显著影响,施氮量对各节位花前和花后转运率、贡献率均存在极显著影响,品种及施氮量互作对花前、花后转运率和贡献率均有显著影响(其余节花后转运率、贡献率除外)(表4)。不同施氮处理下,两个品种A1处理不同节位果聚糖花前转运率均显著低于CK2(P<0.05),果聚糖花后转运率、贡献率及花前贡献率均高于CK2。果聚糖转运率变化范围为16.14%~49.38%,贡献率为4.60%~35.77%。除B1处理,其余节果聚糖贡献率高于倒二节。两品种花后果聚糖对籽粒贡献率均高于花前,XC37花前和花后对籽粒贡献率较XC6增幅分别为44.75%~149.80%和15.37%~62.52%。
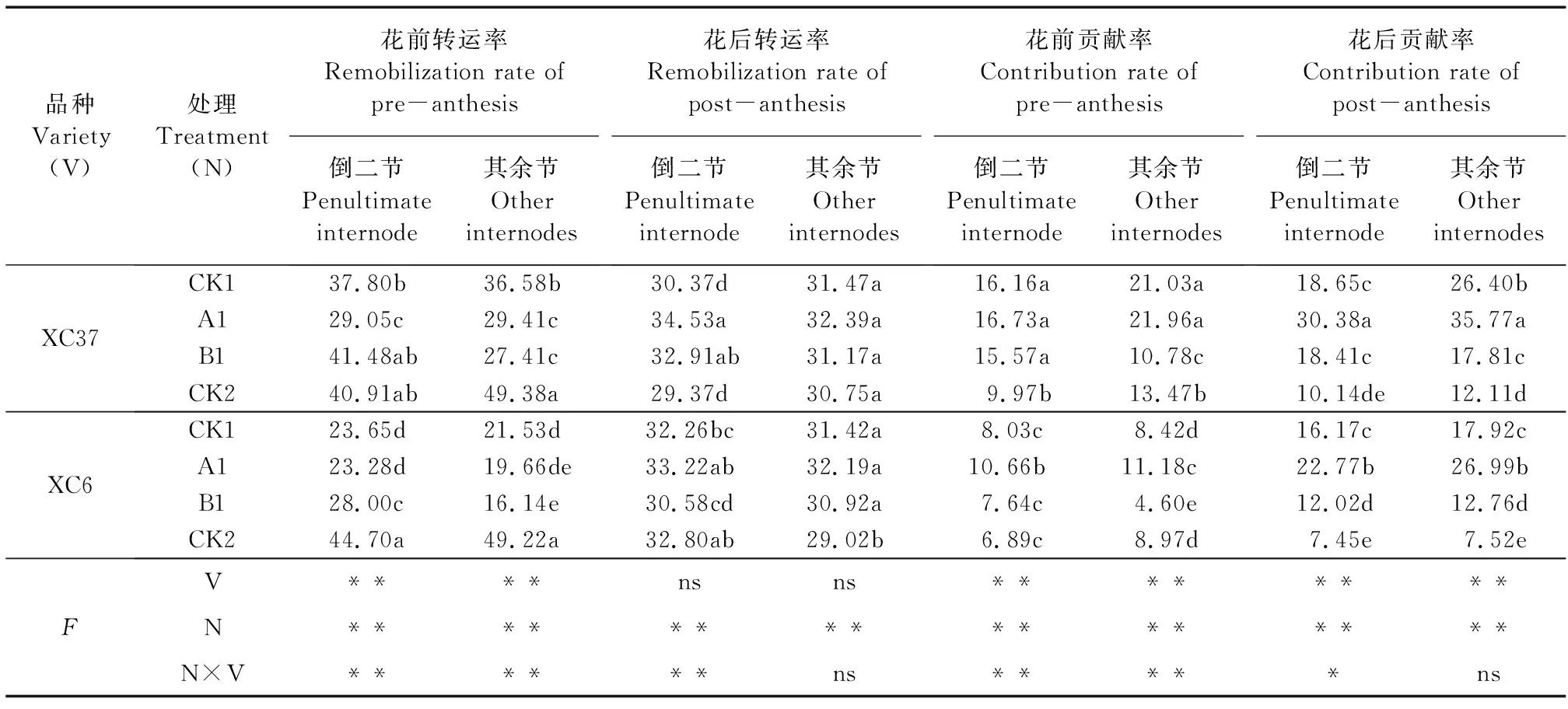
表4 不同施氮处理对滴灌春小麦各节位花前和花后果聚糖转运率、贡献率的影响/%Table 4 Effects of different nitrogen treatments on remobilization rate and contribution rate of fructan in penultimate and other internodes of spring wheat during the pre- and post-anthesis under drip irrigation
由表5可知,施氮量对两节位花前、花后NSC转运率和贡献率有极显著影响(其余节花后转运率除外),品种及施氮量互作对两节位NSC花前转运率、贡献率及倒二节花后转运率、其余节花后贡献率均有显著影响。不同施氮量下各节位NSC花后转运率及花前、花后贡献率均为A1处理最大,均显著高于CK2;花前和花后NSC贡献率表现为A1>CK1>B1>CK2,各处理NSC转运率、贡献率分别为21.43%~44.37%、9.91%~45.65%。不同节位NSC转运率、贡献率略有差异,其余节的NSC花后转运率及其对籽粒贡献率高于倒二节。两品种花后NSC转运率及其对籽粒贡献率均高于花前,提高了55.11%~74.99%;XC37花前、花后NSC对籽粒贡献率比XC6分别提高9.67%~72.83%、30.60%~81.71%。
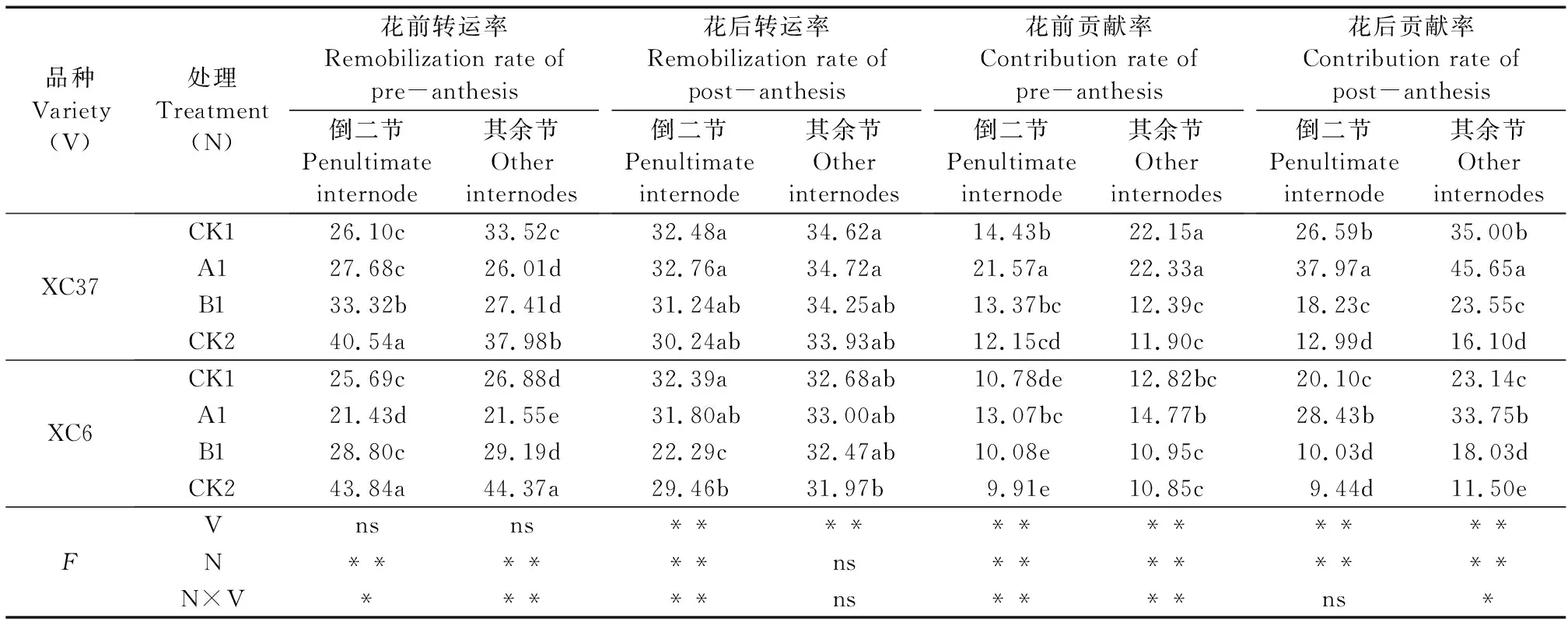
表5 不同施氮处理对滴灌春小麦各节位花前和花后NSC转运率、贡献率的影响/%Table 5 Effects of different nitrogen treatments on remobilization rate and contribution rate of NSC in penultimate and other internodes of spring wheat during the pre- and post-anthesis under drip irrigation
不同施氮量下茎鞘各节位干物质转运率、贡献率存在差异,品种、施氮量均对两节位转运量、转运率和贡献率有显著影响,二者互作对穗下节籽粒贡献率有显著影响(表6)。茎鞘干物质转运率为11.14%~33.83%,对籽粒贡献率为4.28%~27.44%。A1处理两品种转运率、贡献率均比CK2显著提升(P<0.05)。各处理其余节转运率、贡献率较高,倒二节次之,穗下节最低。两品种干物质转运率及对籽粒的贡献率均表现为XC37略高于XC6。
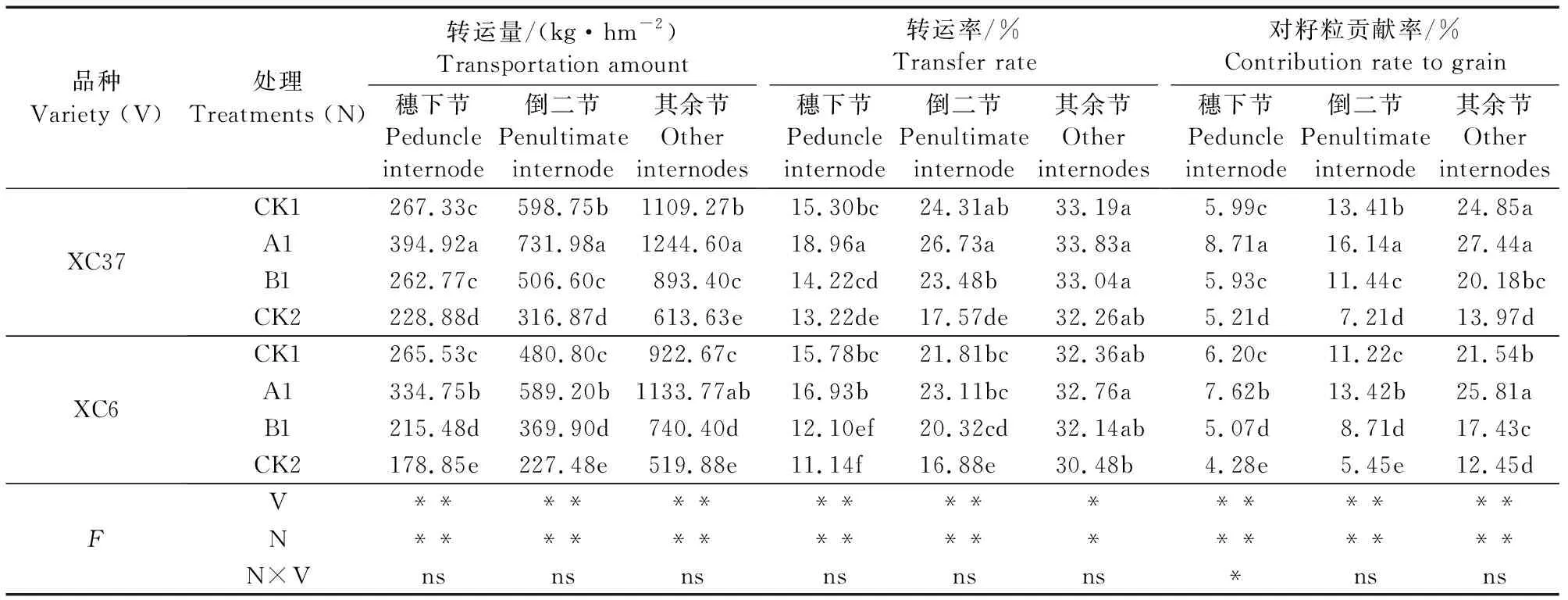
表6 不同施氮处理对滴灌春小麦茎鞘干物质转运量、转运率、贡献率的影响Table 6 Effects of different nitrogen treatments on transfer and contribution rate of dry material in the stem and sheath of spring wheat under drip irrigation
3 讨 论
3.1 施氮量对滴灌春小麦茎鞘中果聚糖代谢酶活性、果聚糖含量及其积累与转运的影响
果聚糖主要储存在小麦茎鞘中,是NSC的主要成分[9],其代谢受SST和FEH两种关键酶调控,SST与果聚糖的合成密切相关,而FEH则与果聚糖的降解有关[10-11]。姜东等[11]认为,在开花后约15~20 d(乳熟期),小麦茎鞘中的果聚糖含量达最大,SST活性也相对较高,开花后25~30 d左右(蜡熟期)SST活性最高。本研究与其结论较为一致,随生育进程推进,小麦茎鞘SST活性于乳熟期达到最大,成熟期最小,而FEH活性在蜡熟期达到最大,成熟期最小;说明生育前期小麦茎鞘SST活性较高,促进茎鞘果聚糖积累,而生育前后期FEH活性相对较高,有利于茎鞘中果聚糖的分解及其向籽粒的运转。本研究进一步发现,在A1处理(施氮量255 kg·hm-2)下,乳熟期其余节果聚糖含量最高,其积累量比倒二节高8.77%,茎鞘中果聚糖花后转运率以及对籽粒的贡献率分别比花前提高36.93%、102.15%,而赵万春等[22]发现,施氮量为200 kg·hm-2时,小麦茎鞘的果聚糖含量在成熟期时相对较高,但较其他时期提升不显著。研究结果差异可能是试验环境不同所致。本研究中,乳熟期‘新春37号’(XC37)其余节果聚糖含量比‘新春6号’(XC6)高17.01%,且XC37其余节果聚糖花前、花后转运率及两者对籽粒的贡献率均高于XC6,分别增加47.41%、1.89%、107.67%、45.12%,表明强筋型小麦(XC37)更有利于茎鞘中贮藏物质向籽粒转运,促进籽粒灌浆。施氮量与品种互作对滴灌春小麦果聚糖花前转运率以及其对籽粒的贡献率具有显著影响。总之,适宜的氮肥供给主要通过增强小麦茎鞘中FEH和SST活性促进其储存的果聚糖向籽粒转运。
3.2 施氮量对滴灌春小麦茎鞘中NSC含量及其积累与转运的影响
促进小麦茎鞘中NSC向籽粒转运,可有效提高粒重和产量[23]。本研究发现A1处理有利于茎鞘中NSC积累,而CK2和 B1处理NSC积累较缓,这与Li等[24]的研究结论一致。说明合理施氮有利于植株茎鞘中NSC积累,施氮过多或过少将导致其积累受抑,不利于产量形成。当施氮量为255 kg·hm-2时,各茎节NSC含量及其对籽粒的贡献率最大,其中,乳熟期其余节NSC含量较倒二节提高13.91%,其余节NSC花前、花后对籽粒的贡献率较倒二节分别提高8.27%和19.47%。本研究还发现,在不同施氮量下,两个品种各节位花前NSC对籽粒的贡献率为9.91%~22.33%,花后NSC贡献率达到9.44%~45.65%,基本表现为花后NSC对产量的贡献率较大。这与马尚宇等[5]研究结果略有不同,其研究发现小麦茎鞘中NSC花前转运率对产量的贡献率较大,规律不同可能是各研究的施肥和灌溉方式不同所致[25]。本研究还表明,在A1处理下,乳熟期XC37其余节NSC含量比XC6增加14.23%,花前和花后转运率分别提高20.70%和5.21%,花前和花后对籽粒的贡献率分别提高51.18%和35.26%。施氮量与品种之间存在显著的互作效应,说明适宜的施氮量有利于提高强筋型小麦XC37茎鞘NSC花前转运率以及对籽粒的贡献率,提高产量。但在滴灌条件下,水氮互作对不同基因型春小麦茎鞘NSC积累转运调控产量形成的规律还有待进一步研究。
3.3 施氮量对滴灌春小麦茎鞘干物质量和产量的影响
茎鞘是小麦花前贮存物质的主要营养器官,一般其贮藏物质转运对小麦产量的贡献率为10%~20%[26]。研究表明,适当增施氮肥可促进小麦茎鞘干物质积累与转运[27],施氮量过多则会抑制茎鞘干物质转运并降低其对籽粒的贡献率[28];在施氮量0~360 kg·hm-2范围内增施氮肥,黄淮海麦区小麦花后茎鞘干物质积累及其对籽粒产量的贡献率呈先升后降的趋势,并在施氮量180 kg·hm-2时达到最大[29]。本试验中,施氮量为255 kg·hm-2时成熟期各节位茎鞘干物质量对籽粒的贡献率最大,其中其余节对产量的贡献率为26.63%,倒二节为14.78%,穗下节为8.17%。牛巧龙等[30]研究发现,施氮量为0~300 kg·hm-2时,强筋小麦和弱筋小麦茎鞘干物质转运率分别为51.1%~56.4%、35.2%~48.3%,对籽粒贡献率分别为32.6%~44.6%、15.9%~29.3%,强筋小麦茎鞘干物质转运及其对籽粒的贡献率高于弱筋小麦;这与本试验研究结果一致,即A1处理下成熟期XC37(强筋型)茎鞘干物质转运及其对产量贡献率均高于XC6(中筋型),两个品种茎鞘干物质转运率分别为18.96%~33.83%、16.93%~32.76%,对产量贡献率分别为8.71%~27.44%、7.62%~25.81%,XC37干物质转运量、转运率以及对籽粒的贡献率分别比XC6高9.78%、3.27%、6.32%,说明氮素对小麦干物质转运量及其对籽粒贡献的影响因品种不同而存在差异[31-33]。
4 结 论
不同施氮条件下,两个春小麦品种各节位茎鞘SST活性、果聚糖含量、NSC含量以及茎鞘干物质量随生育期推进均呈先增后降的变化规律,而FEH活性则表现为先降后增再降。不同节位中,其余节各指标高于穗下节、倒二节。与对照(施氮量0 kg·hm-2)相比,A1处理(施氮量255 kg·hm-2)显著提高了其余节茎鞘果聚糖含量、果聚糖代谢酶活性、NSC含量以及茎鞘干物质量,有利于促进茎鞘果聚糖、NSC以及干物质向籽粒转运,对产量的贡献率分别达到23.98%、29.13%、29.96%;XC37各指标以及产量的提升幅度高于XC6。因此,255 kg·hm-2可作为新疆滴灌春小麦(特别是强筋春小麦)较为适宜的氮肥施用水平进行推广应用。
