纤维素酶的研究与应用进展
2023-10-09门中华
赵 鑫,张 红,门中华,马 勇,温 彤
(内蒙古科技大学包头师范学院,内蒙古 包头 014030)
纤维素作为环境中含量极为丰富的可再生有机物,是构成植物细胞壁的主要物质[1]。有数据显示,全球植物每年生成的干物质高达1 500亿t,其中纤维素和半纤维素的总量约850亿t[2]。近年来,随着环境问题的日益凸显及化石燃料存储量的逐渐下降,人们对新能源的需求不断增加,可再生资源受到越来越多的关注。作为自然界中最丰富的可再生有机物,纤维素的转化利用对于解决环境污染、能源危机等具有十分重要的意义。纤维素酶作为一种高活性的生物催化剂,具有纤维素降解能力,且降解条件温和、高效,在纤维素的转化利用中具有广泛的应用价值。目前,研究人员主要通过选育自然环境中纤维素酶高产菌、优化发酵条件、对微生物和纤维素酶进行生物学改造等手段提高纤维素的利用率。在此,作者在介绍纤维素及纤维素酶的基础上,对纤维素酶的研究进展及其在纺织、造纸、饲料、食品及能源等行业的应用进行综述,以期为纤维素酶的进一步研究提供理论基础。
1 纤维素概述
1.1 纤维素的结构与功能
1.1.1 纤维素的结构特点
纤维素是一类由葡萄糖单元通过β-1,4糖苷键连接成的线性聚合物。天然纤维素主要由植物体合成,是自然界中分布最广、含量最多的可再生有机物,其含有的生物炭占自然界总量的一半以上[3]。在植物细胞壁中,纤维素分子之间通过共价键和非共价键紧紧连接,形成较为致密的网状结构。纤维素、半纤维素及木质素相互结合形成了复杂的超分子化合物,进而形成了极为坚固的植物细胞壁结构[4]。在对这类结构的研究中,二相体理论[5]得到了目前为止最为广泛的认可。该理论认为,细胞壁中纤维素主要以两种形式排列:一种是由直链的大分子纤维素整齐排列、折叠而成的结构致密的结晶区;另一种则是由纤维素分子通过不规则排列形成的无定形区[6-8]。
1.1.2 纤维素的生物学功能
纤维素作为陆生植物的骨架材料,经过长达数十亿年的进化,具有极强的维持细胞形态和保护细胞的功能,其生物学功能主要体现在能够提高细胞的机械强度,从而保护细胞免受机械损伤以及渗透压等外力的破坏;另外,纤维素作为细胞壁的主要骨架,承受着内部由于液泡吸水膨胀而产生的扩张力,从而使植物细胞及器官具有固定的形态。此外,由于纤维素的单体结构为葡萄糖,其也是重要的生物碳源和能量来源;但由于其结构致密稳定,因此在自然界中难以被降解和广泛利用[9]。
1.2 纤维素的降解利用
在自然界中,能够降解并利用纤维素作为碳源和能源的生物主要包括部分细菌、真菌及原生动物等,其利用方式多为:纤维素分子经不同类别的葡聚糖水解酶协同作用,降解为单糖或小分子寡糖,随后被进一步分解利用。另外,随着农业生产水平的不断提高,以秸秆、谷壳等为代表的富含纤维素的残余物数量也急剧增加。为了有效处理和利用这类农产品残余物,纤维素高效降解技术的开发和应用逐渐成为研究热点。目前,纤维素的降解方法主要有物理降解法、化学降解法及生物降解法[10]。
1.2.1 物理降解法
物理降解法主要是通过对纤维素类物质进行破碎处理,大幅降低细胞壁中纤维素的结晶度,同时增强破碎后小分子组分的溶解能力,从而改变细胞壁特有的紧密稳定结构,达到降解纤维素的目的[11]。目前,常用的纤维素物理降解法包括微波处理法、冷冻粉碎法、机械破碎微粒法等,普遍存在操作复杂、能耗大、成本高等缺点。纤维素进行物理降解前通常需进行酸碱浸泡等预处理,不仅延长了降解周期,且易造成二次污染。因此,纤维素的物理降解法在实际应用时受到较大限制[12]。
1.2.2 化学降解法
化学降解法是通过化学试剂将半纤维素、纤维素及木质素等大分子溶解,在降低结晶度的同时提高其溶解度,从而达到降解的目的。目前,常用的纤维素化学降解法包括碱水解法、酸水解法、有机溶剂法等[13]。与物理降解法相比,化学降解法操作简单,但存在相似的弊端,即降解产物及设备均需进行复杂的后处理以避免酸碱污染。
1.2.3 生物降解法
生物降解法是指在特定条件下,纤维素被微生物完全分解为小分子化合物的过程。与物理降解法及化学降解法相比,生物降解法具有成本低、无污染、条件温和、操作简单、对仪器设备要求低等特点,且生物降解法可破坏纤维素、半纤维素、木质素这3种分子之间的紧密结构,实现彻底降解。基于以上优势,纤维素的生物降解法逐步占据主导地位[14]。
微生物对纤维素的降解主要通过不同葡聚糖水解酶的协同作用完成。目前,研究人员针对纤维素在机体中的降解过程开展了大量研究,提出了众多假说,其中C1-Cx假说、顺序假说、协同作用假说、短纤维形成假说具有广泛认可度和较高权威性。
1950年,Reese[15]首次提出了C1-Cx假说,该假说认为外切葡聚糖酶(C1)、内切葡聚糖酶(Cx)及β-葡萄糖苷酶(BG)是按照固定步骤对纤维素进行降解,即首先纤维素的结晶区被C1酶结合并断开,然后由Cx酶水解大分子多糖生成纤维二糖及其它小分子,最后由BG酶作用于纤维二糖的糖苷键,生成小分子葡萄糖。
1987年,Enari等[16]提出的顺序假说与C1-Cx假说相反,该假说认为首先Cx酶作用于纤维素的结晶区,再由C1酶进攻大分子多糖生成纤维二糖或其它小分子,最后BG酶降解纤维二糖生成小分子葡萄糖。
协同作用假说是目前大多认可的纤维素降解理论,该假说认为纤维素酶组分之间存在类似于C1-Cx假说、顺序假说的协同作用,且C1酶、Cx酶及BG酶的组成比例对纤维素的降解具有决定性作用[17-18]。
短纤维形成假说认为,首先解链因子或解氢键酶作用于天然纤维素上,破坏纤维素链的氢键,纤维素长链变成短链,再通过C1酶、 Cx酶及BG酶的协同作用生成小分子葡萄糖。
2 纤维素酶概述
2.1 自然界中纤维素酶的微生物来源
2.1.1 原核纤维素降解微生物
原核纤维素降解微生物主要来源于细菌及放线菌。其中细菌的主要来源菌属为纤维粘菌属、芽孢杆菌属、热双歧菌属和欧文氏菌属等[19],细菌分泌的是单一酶,通常为内切纤维素酶,应用pH值范围为中性和碱性,酶活低,产量低,提纯难度高,且多为胞内酶[20]。放线菌的主要来源菌属为纤维放线菌属、诺卡氏菌属和链霉菌属,其分泌酶的应用pH值范围较广,且在高温环境中也具有较高的活性,编码酶的基因较为简单,更容易进行基因重组与改造,但其菌株生长速度较慢且酶产量较低,因此,针对放线菌的研究较少[21]。
2.1.2 真核纤维素降解微生物
真核纤维素降解微生物主要来源菌属为木霉属、曲霉属和青霉属等,其分泌的是复合酶,相比于原核纤维素降解微生物,酶活较高,产量较高,且为胞外酶,易于提取,但其应用pH值范围通常为酸性,只能应用于酸性条件下的工业生产[22-24]。
原生生物通常会出现在宿主后肠中,与其它微生物共同作用降解宿主体内的纤维素,这些宿主包括线虫、鳌虾、福寿螺、白蚁、天牛等[25],单一物种的原生生物产生的纤维素酶通常为单功能酶。因此,虽然不同种类原生生物对纤维素的降解机制不尽相同,但其对纤维素的降解均需要多种单功能酶的协同作用。
2.2 纤维素酶的类别与功能
纤维素酶是由内切葡聚糖酶、外切葡聚糖酶、β-葡萄糖苷酶等具有单一功能的酶组成的复合酶。纤维素在这3种酶的协同作用下,被降解成小分子葡萄糖,其过程如图1a所示[26]。
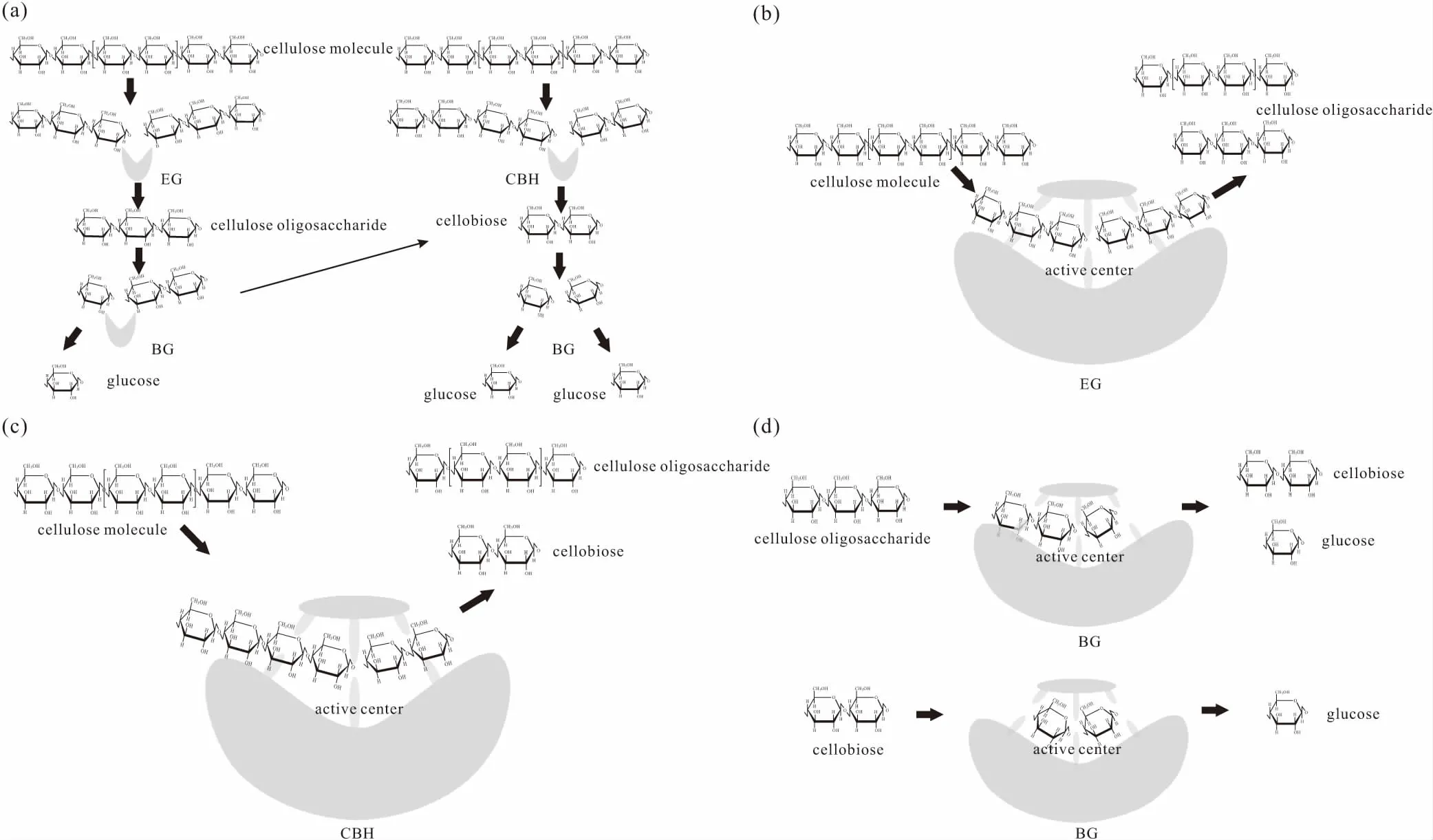
图1 纤维素酶组分协同降解纤维素(a)及内切葡聚糖酶(b)、外切葡聚糖酶(c)、β-葡萄糖苷酶(d)的作用机制Fig.1 Action mechanism of synergistic degradation of cellulose by cellulase components(a),endoglucanase(b),exoglucanase(c),and β-glucosidase(d)
2.2.1 内切葡聚糖酶
内切葡聚糖酶或内切-β-1,4-葡聚糖酶(EC 3.2.1.4),也称作CMC酶、Cx酶,简称EG(真菌)和Cen(细菌)。内切葡聚糖酶随机地切割纤维素多糖链中无定形区,生成长度不一的寡糖,包括纤维二糖、纤维三糖及纤维糊精等,这些寡糖成为新的链端,为下一步降解作好铺垫[17](图1b)。
2.2.2 外切葡聚糖酶
外切葡聚糖酶分为2种酶:一种是β-1,4-D-葡聚糖-纤维二糖水解酶;另一种是β-1,4-D-葡聚糖-葡萄糖水解酶(EC 3.2.1.74),简称CBH(真菌)和Cex(细菌)。外切葡聚糖酶主要切割位点为纤维素长链的还原末端或非还原末端,产生不同的寡糖,包括纤维二糖及葡萄糖等小分子(图1c)。
根据识别位点,外切葡聚糖酶又可分为CBH Ⅰ和CBH Ⅱ两类[27],其中前者的作用位置是纤维素的还原末端,而后者的作用位置是非还原末端。如里氏木霉(Trichodermareesei)可以表达还原性外切葡聚糖酶TrCel7A[28],还可以表达非还原性内切葡聚糖酶TrCel6A[29];Phanerochaetechrysosporium可以表达还原性内切葡聚糖酶PcCel7D[30]。
2.2.3 β-葡萄糖苷酶
β-葡萄糖苷酶或β-1,4-葡萄糖苷酶(EC 3.2.1.21),简称BG。β-葡萄糖苷酶将纤维二糖(其它短链的纤维寡糖)或纤维糊精水解成小分子葡萄糖[19,31](图1d)。
2.3 纤维素酶的结构
大量研究表明,细菌与真菌体内均含有纤维素酶且结构相似[32]。较为典型的纤维素酶通常是由结合结构区域(cellulose binding domain,CBD)、催化结构区域(catalytic domain,CD)和连接结构区域(linker)等3部分组成[33],另外部分纤维素酶分子结构中含有其它特殊催化结构。
2.3.1 结合结构区域
结合结构区域呈楔形的尾巴结构,在纤维素酶催化纤维素降解过程中行使识别催化位点的作用,并将纤维素酶分子与纤维素特异性连接到一起。在纤维素酶分子催化降解纤维素的整个过程中,结合结构区域虽然不起催化作用,却是整个催化过程开始的关键一步。研究[34]表明,结合结构区域不会影响纤维素酶的活性,各类纤维素酶的结合结构区域在纤维素催化降解中的作用机制类似。
2.3.2 催化结构区域
催化结构区域呈球形结构,在纤维素降解过程中主要负责催化β-1,4糖苷键断裂。外切葡聚糖酶与内切葡聚糖酶的催化结构区域的活性位点不同、催化机理也有所不同[35]。其中,前者活性位点在长圆形管道中,可水解不溶于水的纤维素结构,解离长链分子,在与还原末端作用时能达到更好的催化作用;后者活性位点在向外开口处,该开口可与非结晶纤维相结合[36]。
2.3.3 连接结构区域
连接结构区域是一段高度糖基化多肽链,用于连接结合结构区域与催化结构区域,其中高度糖基化可以抵御其它酶的水解,从而确保该结构更加稳定,保证结合结构区域与催化结构区域的距离,使不同酶形成相对稳定的聚集物。
3 纤维素酶的研究进展
3.1 纤维素酶的筛选
有数据显示,自然界中微生物的物种数量有一万亿种,其中可以实现实验室培养的物种不足1%[37]。为了获得性能优异的纤维素酶,筛选优良菌株是最直接、最有效的方法。筛选优良菌株的途径很多,例如土壤、水体及动物肠道、粪便等[38]。目前,从猪粪便[39]、鱼类肠道[40]、白蚁肠道[41]以及土壤[42]中均成功分离获得了纤维素酶,Rathore等[43]在水葫芦湖区的水源中筛选出产纤维素酶菌。此外,在部分极端环境中也存在产纤维素酶菌,且具有独特的性质。Priya等[44]从喜马拉雅山西北部的塔塔帕尼温泉的沉积物中分离出 8 株产纤维素酶的嗜热菌。
虽然目前筛选分离出大量具有产纤维素酶能力的微生物,但所产纤维素酶依然存在降解效率低、酶活性低及受水解底物抑制等问题。因此,纤维素酶高产菌株的选育与发酵条件的优化是有效提高纤维素降解效率的重要途径。
3.2 纤维素酶高产菌株的选育
3.2.1 纤维素酶高产菌株的诱变育种
目前,最直接、最有效的选育纤维素酶高产菌株的方法为诱变育种[45-46]。
3.2.1.1 物理诱变育种
物理诱变育种主要包括紫外线、X-射线、微波、快中子、γ-射线及重离子束辐照等。Raghuwanshi等[47]通过紫外线对棘孢木霉RCK2011进行诱变处理,筛选获得的突变菌株 SR1-7的滤纸酶活最高达到2.2 U·g-1;突变菌株产生的纤维素酶培养液对麦草、玉米芯和甘蔗渣等生物质的降解效率显著提高。Li等[48]通过紫外线与微波联合诱变处理1株绿色木霉,获得了 M-B1、M-B2、M-B3、M-B4、M-B5、M-B6、M-B7 等7 株优良突变菌株,其酶活比诱变前显著提高。
重离子束辐照是一种较为先进的物理诱变育种法,因具有DNA 损伤后难以正确修复、传能线密度高、相对生物学效应高及突变谱广等特性,在菌种选育中具有独特的优势,受到越来越多研究者的关注[49-50]。Dong等[51]利用重离子束辐照对野生型烟曲霉菌株进行诱变处理,筛选获得了酶活较高的突变株MS160.53,其FPA和CMC酶活分别达到1.81 U·mL-1和1.11 U·mL-1,其对碱预处理秸秆的还原糖含量达到789.75 mg·(g秸秆)-1。Wang等[52]利用重离子束辐照诱变处理绿色木霉,获得1株高产菌株,其液体发酵液中滤纸酶活最高达到688.99 U·mL-1,较诱变前提高了约71.29%。
3.2.1.2 化学诱变育种
化学诱变育种主要是通过使用化学诱变剂(如亚硝基胍、甲基磺酸乙酯、亚硝酸盐等)来改变微生物的遗传物质,从而达到诱变育种的目的。Gadgil等[53]利用紫外线与亚硝酸钠对里氏木霉QM9414进行复合诱变处理,筛选获得了1株纤维素酶高产突变菌株,其发酵液中滤纸酶活最高达到0.54 U·mL-1;同时,其对纤维素底物的糖化能力是诱变前的1.2倍。Jiang等[54]通过甲基磺酸乙酯化学诱变与紫外线诱变相结合的方式对绿色木霉进行诱变处理,筛选获得了1株β-葡萄糖苷酶高产突变菌株EU2-77,所产纤维素酶对底物的水解能力远远高于诱变前。
3.2.2 纤维素酶的异源表达
纤维素酶的异源表达主要是通过人为定向改造纤维素酶合成结构基因或调控基因实现纤维素酶基因在宿主中高效表达与分泌。异源表达的一般步骤为:目的基因的克隆、宿主载体的选取、重组菌株的建立、目的基因的诱导表达。自20世纪70年代,纤维素酶的编码基因开始逐步被克隆并实现了纤维素酶基因的异源表达。Fang等[55]将来自外源基因的cbh1强启动子克隆于里氏木霉菌bgl1 基因的上游以提高该基因的表达水平,结果发现,改造后菌株的发酵液中β-葡萄糖苷酶活性显著提高,同时其纤维素酶蛋白的表达量显著增加。Ma等[56]在里氏木霉菌株中通过采用外源基因的cbh1强启动子表达了斜卧青霉来源的β-葡萄糖苷酶基因,结果发现,改造后菌株的发酵液中β-葡萄糖苷酶活性提高了6~8倍,滤纸酶活提高了约30%;此外,使用改造菌株的发酶液进行玉米秸秆的糖化实验时发现,其糖化效率也显著提高。Wang等[57]在里氏木霉中对来源于黑曲霉的β-葡萄糖苷酶基因进行异源表达,结果发现,改造后菌株的β-葡萄糖苷酶活性显著提高,与改造前相比提高了106倍,同时其滤纸酶活也提高了44%。
3.3 纤维素酶的发酵工艺优化
对于产纤维素酶微生物来说,碳源是影响其酶产量最重要的因素;纤维素酶作为一种诱导酶[58],其产生过程需要多糖类物质的诱导。因此,同时作为碳源和诱导物的多糖类物质通常是纤维素酶发酵工艺优化的重点。研究发现,废纸[59]、甘蔗渣[60]和生活污水[61]等可用作产纤维素酶微生物的诱导底物。此外,培养条件如发酵温度、搅拌转速、发酵液pH值等对酶产量同样重要。其中,发酵温度超过一定范围就会导致微生物丧失细胞功能甚至死亡[62];搅拌转速对微生物的影响主要体现在供氧和剪切力上,搅拌转速与供氧、剪切力成正比;发酵液 pH 值通过改变细胞膜电荷而影响微生物活性[63]。
3.4 纤维素酶的改性
选育纤维素酶高产菌株及优化纤维素酶发酵工艺对提高纤维素酶产量是有限的,无法完全满足工业化应用。因此,为进一步提高纤维素酶活性、稳定性以及其它有利于大规模应用的特性,需采用分子修饰对纤维素酶进行改造。目前,常用的分子修饰策略包括化学修饰、定点突变等,其中化学修饰最常见的方法有接枝可溶性大分子或小分子修饰、交联修饰等[64]。
3.4.1 接枝可溶性大分子或小分子修饰
酶的接枝可溶性大分子修饰是将可溶性的大分子修饰剂(如聚乙二醇、葡聚糖等)与酶分子通过共价键连接在一起,从而提高酶的活性及稳定性,降低酶的抗原性[65]。Park等[66]通过在纤维素酶中引入两亲性共聚物共价偶联,改变了酶的吸附参数且提高了酶活。张裕卿等[67]利用聚氧乙烯-马来酸酐聚合物改性纤维素酶,改性后纤维素酶的高温耐受性显著提高,且可以有效促进盾叶薯蓣薯蓣皂苷向薯蓣皂苷元的转化。
由于纤维素酶分子表面有近一半为非极性基团,从而导致纤维素酶的稳定性较差,为了提高纤维素酶的稳定性和活性,可以利用小分子对非极性基团进行修饰。Bund等[68]在2~4 ℃下将酸性纤维素酶与 0.5 mol·L-1马来酸酐连接在一起,修饰后的纤维素酶对碱性环境的耐久度显著提升。
3.4.2 交联修饰
酶的交联修饰是将蛋白质交联剂作用到酶分子内或酶分子间使酶分子发生交联。Zheng等[69]采用超细研磨结合磷酸盐交联纤维素酶,改善了谷子麸皮膳食纤维的体外降血糖和降血脂性能。Li等[70]通过纤维素酶与硫酸铵沉淀,然后与戊二醛交联形成交联纤维素酶聚集体,其具有更高的催化活性、更强的适应性,且重复使用次数大幅增加。
3.4.3 定点突变
酶的定点突变是将氨基酸残基通过定点突变技术连接到酶分子上的靶向位置,然后对该氨基酸残基进行一系列修饰,最终得到突变酶。Yang等[71]采用随机突变和定点突变相结合的定向进化方法提高CtLac的漆酶活性,筛选得到的菌株V243D在最适条件(70 ℃,pH值8.0)下对木质素的氧化降解能力明显增强,且高值醛产量提高了50%~95%。Zhou等[72]对内切葡聚糖酶(CgEndo)进行异源表达和定点突变,结果显示,突变菌株Y63S和N20D/S113T的酶活分别提高了62.31%和57.14%。
4 纤维素酶的应用
纤维素在自然界中分布极为广泛且含量丰富,纤维素酶可将富含纤维素的工业废料和农副产品等有效转化,对于提高原料利用率、综合利用资源等具有重要意义。纤维素酶已广泛应用于纺织、造纸、饲料、食品及能源等行业。
4.1 纺织行业
为了解决纺织行业所存在的环境污染问题,纤维素酶的应用逐渐广泛。纤维素酶可以分解棉织物上的小纤维,防止绒毛起球。纤维素酶在牛仔纺织物的生物去石、纺织物及服饰软化、纺织物纤维的生物抛光及纺织物表面染料的去除等方面[73-74]应用广泛。
4.2 造纸行业
纸浆和纸张作为一种可循环利用的再生资源,其回收及利用对于节约能源、减轻环境污染具有重要意义[75]。脱墨是纸浆处理的重要环节,利用纤维素酶脱墨不但可以防止纸张变黄,而且避免了碱液的使用。与碱性纤维素酶和中性纤维素酶相比,酸性纤维素酶可以显著提高纸张的亮度和清洁度,处理效果更好[76-77]。另外,纤维素酶参与纸浆处理,一方面可以有效提高打浆性能,另一方面可以在不改变纸浆强度的前提下显著改善滤水性能,从而得到较为柔软的薄纸张[78]。
4.3 饲料行业
饲料品质对于禽畜产品品质具有决定性作用。由于反刍动物的反刍胃中含有能够分泌纤维素酶的微生物且酶活性很高,因此,反刍动物对于富含纤维素的饲料具有一定的消化吸收能力,但其对饲料中营养物质的利用效率有限[79]。内切葡聚糖酶作为一种饲料添加剂,其主要通过作用纤维素类饲料的细胞壁,使纤维素结构发生改变,将难以降解的纤维素分子降解为短链寡糖,从而使其营养物质更易被动物消化系统分解吸收,提高饲料的营养价值[80];研究证实,纤维素经内切葡聚糖酶降解后形成的部分寡糖具有类似益生元的功能,即这类物质可以被肠道的有益菌选择性地降解并加以利用,而不能被动物后肠的有害菌降解利用,因而具有增强机体免疫力及提高生育能力等作用[81-82]。
4.4 食品行业
纤维素酶在食品行业的应用极其广泛。例如在啤酒生产中,纤维素酶可以增强过滤性能,提高啤酒产率及品质[83];在白酒生产中,纤维素酶可以降解原料中的大量纤维素,最终形成小分子葡萄糖,为酵母对原料的分解利用提供了便利,进而提升原料中纤维素的利用率及出酒率;在葡萄酒酿造过程中,添加纤维素酶可以提升葡萄酒香味,从而改善葡萄酒品质[84];在酱油酿造过程中,添加纤维素酶不但可以加快发酵速度,而且可以改善酱油风味和品质;在茶叶加工过程中,添加纤维素酶可加快茶多糖的释放,从而提高茶多酚含量。
4.5 能源行业
我国作为农业大国,农业生产中产生的大量富含纤维素的农业废弃物处置不当不仅加剧环境污染,而且造成资源浪费。纤维素酶可以将农业废弃物中的纤维素转化为寡糖甚至是葡萄糖,这些寡糖可经过发酵生产乙醇类产品[85]。另外,在石油开采中,由纤维素酶制成的破乳剂具有零污染、专一性强、没有任何副作用等优点[86-87]。
5 展望
目前,利用纤维素酶对纤维素或富含纤维素原料进行降解或改性已广泛应用于纺织、造纸、饲料、食品及能源等行业。但作为蛋白质,纤维素酶存在稳定性低、耐极端温度和pH值能力差、生产成本高等问题,限制了其应用范围。因此,通过天然纤维素酶筛选、产纤维素菌种改造、发酵条件优化和酶分子改性等手段提高纤维素酶的性能和产量,降低其生产成本一直是研究热点。
