低温胁迫对4 种蔷薇属植物的生理指标及HSP90 基因表达的影响
2023-10-04王翔赵琛闫舟张琛浩曹睿
王翔 赵琛 闫舟 张琛浩 曹睿


摘要:为研究4 种蔷薇属植物(毛萼蔷薇、大花香水月季、维西蔷薇、丽江蔷薇)的抗寒性差异,以筛选出耐寒性强的品种,利用低温培养箱进行不同梯度低温(0、-5、-10、-15 ℃)胁迫处理,测定4 种蔷薇属植物可溶性糖含量、可溶性蛋白含量、丙二醛含量、脯氨酸(Pro)含量、SOD 活性、POD 活性及耐寒相关调控基因HSP90 的表达差异。结果表明,在低温胁迫下,4 种蔷薇属植物叶片可溶性蛋白含量、可溶性糖含量、游离脯氨酸含量、丙二醛含量、SOD 活性和POD 活性均随低温胁迫加剧呈现逐渐升高趋势。对4 种蔷薇属植物HSP90 基因的差异表达分析表明,该基因在4 种植物中的相对表达量随低温胁迫加剧呈现逐渐升高趋势,其中,毛萼蔷薇在0、-5、-10、-15 ℃低温胁迫下HSP90 基因表达量均最高。表明4 种蔷薇属植物的抗寒性存在差异,其中,毛萼蔷薇的抗寒性较好。
关键词:蔷薇属;生理变化;低温胁迫;基因差异表达
中图分类号:S685.12 文献标识码:A 文章编号:1002?2481(2023)03?0241?08
月季(Rosa hybrida) 是蔷薇属(Rosa) 多年生木本花卉,素有“花中皇后”之美誉,因其花色丰富、品种繁多而深受人们喜爱。月季具有花期相对较长、适应性广的特征,让其名列我国十大名花,在盆栽以及地栽等场景广泛栽种,有着巨大的市场潜力。近些年,随着现代月季品种性状改良的不断发展,低温环境对于月季品质所产生的影响不断增加,诸如低温会造成枝条发育受到限制、花瓣出现畸形等问题,给驯化和培育种植带来了更多挑战。当前,冬季环境下显著偏低的温度极大影响到花卉的品质,因此,选择和培育有耐寒特征的月季品种,是加强月季推广的重要方式[1-6]。低温会极大影响到植株的生长与分布,对于植物发育所产生的影响会显著大于其他环境因素[7-9]。低温胁迫一方面会显著影响到植株生物量等关键因素,另一方面还会造成代谢紊乱,在超低温的环境下还可能导致植株出现死亡[10]。有研究结果表明,植物在识别到外部环境温度过低时,会自发启动防御,随之而促成细胞膜透性的提升,因此,产生活性氧,保护酶也较为快速地增多,渗透调控物质SP、SS 与Pro[11]以及MDA含量增加[12],而与代谢有密切联系的生理活动会遭遇显著抑制[13]。在植物抗寒性的分析中, LT50与相关生理指标通常被视为抗寒性的重要指标[14-15]。植物在低温环境中的生理与生化表现可能会因各种基因的调控机制产生转变,并再度诱导相关基因的表达,进一步避免逆境损害的出现[16]。HSPs(热激蛋白)对外界环境变化敏感,在逆境胁迫下高表达的HSPs 对细胞可能起到一定保护作用[17],提高机体对逆境的适应能力,从而增强抗逆性。也有学者研究发现,HSP 热激蛋白会在相关胁迫中产生显著的转录调节效果[18]。目前特定基因的表达研究在低温响应机制、重要基因挖掘等领域取得了很多成果,广泛运用到拟南芥[19]、小麦[20]等各类作物的分析中。当前科学家寻找到数百个基因对于应答反应会产生巨大的影响,XU 等[21]通过对野生山葡萄开展深度分析,得出其依靠管控相关基因的差异表达,进而应对外界的寒冷环境。中外学者围绕抗寒性的分析,多数是关于生理指标的对比性分析[22-23],而围绕抗寒机制开展分子水平研究显著偏少。
为更加深入研究月季抗寒原理,本研究选择4 种蔷薇属植物,分析低温胁迫中HSP90 基因表达变化和相关生理指标差异,为探究分析月季抗寒机制与品种选育提供一定的数据支撑,以期为月季栽培推广提供支持。
1材料和方法
1.1 试验材料
试验选用毛萼蔷薇(R. lasiosepala)(编号S1)、大花香水月季(R. odorata. var. gigantea)(编号S2)、维西蔷薇(R. weisiensis)(编号S3)和丽江蔷薇(R. lichiangensis)(编号S4)4 种蔷薇属植物的盆栽苗为研究对象,均来自云南省农业科学院花卉研究所蔷薇属种质资源圃,为3 年生嫁接植株,植株生长健壮。
1.2 試验方法
2020 年12 月,选定发育无异常、长势较为接近的扦插苗54 株分别栽入到花盆,放在温室(温度设定为25 ℃,湿度设定为65%,光强为200 lx,光周期为14 h 光照/10 h 黑暗)中持续培育14 d。每个处理选用3 株,每个处理3 次重复。分别在人工低温气候箱进行0、-5、-10、-15 ℃低温下胁迫2 h,从各处理植株选取成熟叶片,用于半致死温度和各相关生理指标的测定。同时迅速取各植株顶端幼嫩叶片装入到离心管中,立即投入到液氮中降温速冻后,置于-80 ℃ 冰箱中保存,用于RNA 的提取和qPCR 试验。以上试验重复3 次。
1.3 测定项目及方法
1.3.1 生理生化指标的测定 参考陈建勋等[24]方法测定丙二醛(MDA)含量、游离脯氨酸(Pro)含量、SOD 活性、POD 活性与可溶性糖(SS)含量、可溶性蛋白(SP)含量。
1.3.2 总RNA 提取与cDNA 合成 使用RNA ExtractionKit(大连Takara)提取得到各植株叶片的总RNA。使用NanoDrop 2000 测定获得具体的RNA纯度、完整性以及RIN 值。然后使用qPCR 逆转录盒PrimeScript TMRT reagent Kit(大连Takara)合成cDNA,具体流程详见相应说明书。
1.3.3 qPCR 候选基因与引物设计 参考李瑞雪等[25]所提出的方法,完成HSP90 基因差异表达研究,应用qPCR 分析使用TB Green Premix Ex Taq Ⅱ(大连Takara),结合说明书的流程稳定推进。参照文献[26]的方法计算基因相对表达量。
1.4 数据分析
试验采用Excel 2016、SPSS 19.0 软件进行数据分析。
2结果与分析
2.1 低温胁迫对蔷薇属植物LT50的影响
不同蔷薇属植物的LT50比较结果如图1 所示。
由图1 可知,4 种蔷薇属植物LT50 为-18.87~-10.52 ℃,而S1 與S2 的半致死温度均低于-15 ℃,S3 与S4 的半致死温度低于-10 ℃,但高于-15 ℃。可见4 种蔷薇属植物的LT50 指标从小到大依次为S1
2.2 低温胁迫对蔷薇属植物可溶性蛋白含量的影响
由图2 可知,在0 ℃环境下开展低温胁迫,4 种蔷薇属植物叶片的SP 指标差异显著,可溶性蛋白含量随着温度降低呈现上升趋势。S1、S2 与S3 的可溶性蛋白含量在-15 ℃胁迫下达到了峰值,S4 则是-10 ℃胁迫下达到峰值。在峰值时,SP 含量由高到低依次为S1>S2>S3>S4,与0 ℃ 低温胁迫时相比,4 种植株增长最低为127%,最高为142%。可以看出,LT50更低的蔷薇属植物,其SP 指标的上升幅度更高。
2.3 低温胁迫对蔷薇属植物可溶性糖含量的影响
低温下4 种蔷薇属植物可溶性糖含量的变化如图3 所示。
由图3 可知,4 种蔷薇属植物叶片的SS 指标随着胁迫温度降低而逐渐升高。在0 ℃低温胁迫时,各品种间的可溶性糖含量差异显著(P<0.05),其中,S1 的SS 含量显著高于其他蔷薇属植物。在-15 ℃胁迫下,4 种蔷薇属植物可溶性糖含量表现为S1>S2>S3>S4,与0 ℃低温胁迫时相比,4 种蔷薇属植物最大增长47%,最小增长19%,可见,LT50更低的植株在低温状况下积累的SS 更多。
2.4 低温胁迫对蔷薇属植物游离脯氨酸含量的影响
在低温胁迫的影响下,4 种植株的Pro 含量呈现持续上升的趋势(图4)。在-15 ℃时,Pro 含量均达到峰值,具体表现为S1>S2>S3>S4,和0 ℃低温胁迫相比,4 种植株样本最高增加了382%,最低增加了299%,可见不同的样本在低温胁迫下Pro含量有显著差异(P<0.05)。
2.5 低温胁迫对蔷薇属植物丙二醛含量的影响
低温下4 种蔷薇属植物丙二醛含量的变化如图5 所示。
由图5 可知,4 种蔷薇属植物丙二醛含量表现出不断上升的趋势。低温胁迫中,LT50 相对较低的植株MDA 含量更低。4 种蔷薇属植物在0 ℃低温胁迫下的MDA 含量差异显著(P<0.05)。并且随着胁迫温度的降低,MDA 含量也不断增加,在-15 ℃ 胁迫下达到峰值;LT50 相对较高的植物在低温胁迫下会有着更高的MDA 积累。在不同低温胁迫下,MDA 含量均表现为S2
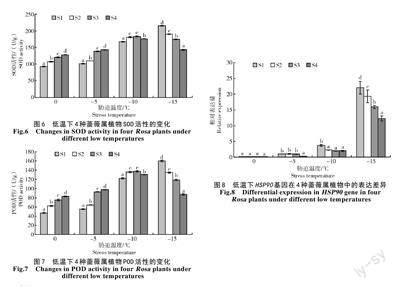
2.6 低温胁迫对SOD 和POD 活性的影响
由图6、7 可知,在低温胁迫下,4 种植物SOD 与POD 活性的变化趋势较为相似,表现为S1、S2 呈现持续上升,S3、S4 则是先上升后下降的变化趋势,同时4 种植株的SOD 与POD 活性在接近LT50温度会有显著下降(P<0.05)。0 ℃低温胁迫时,S1 的SOD与POD 活性显著降低(P<0.05),并且4 种植物的差异显著;在-5 ℃低温胁迫下,仍表现为类似的变化趋势,-10 ℃低温胁迫下S2 与S3 的SOD 活性非常接近,相对于0 ℃ 下增幅分别为62% 与47%,而POD 活性相较于0 ℃时增幅分别为106% 与75%,S1、S2 的SOD、POD 活性在-15 ℃低温胁迫下活性较高,S3、S4 的SOD 与POD 活性则出现了一定程度的下降,并且4 种植物的差异显著(P<0.05)。
2.7 低温胁迫对HSP90 基因表达差异的影响
从图8 可以看出,低温胁迫下HSP90 在4 种蔷薇属植物叶片中的具体表达会有持续提升的变化。0 ℃胁迫情况下,HSP90 基因的相对表达量在4 种植物间差距较小;在-5 ℃ 胁迫下,4 种植物为更好地减少低温负面影响,会开启内部的耐寒基因;接近-10 ℃胁迫下,4 种植物的实际表达会再度产生激活;而-15 ℃胁迫下,基因产生了深度的激活。相关低温影响下,HSP90 的表达表现为S1>S2>S3>S4,可见4 种蔷薇属植物通过HSP90 的表达增强,进而更加有效地应对低温环境,而且该基因表达更高的植物会表现出越强的耐寒性。
3结论与讨论
3.1 叶片低温半致死温度与抗寒性
低温胁迫的影响下,植物会产生相关的生理生化变动,进一步更好地应对外部的低温环境[27-28]。LT50能够较为精准地表现抗寒性能,该指标更低则意味着抗寒性更优秀[29]。温度低于LT50的情况下,植物细胞会产生较为迅速的结冰,生理代谢机制会产生转变,会有相关的冻害现象,使得植株遭遇的伤害也难以恢复[30]。因此,LT50 能够充当作物遭遇无法逆转损害的阈值。本研究表明,4 种蔷薇属植物LT50 在-18.87~ -10.52 ℃ ,从低至高的依次是S1
3.2 叶片渗透调节物质含量与抗寒性
可溶性蛋白(SP)是植物必不可少的一种渗透调节物质,和植物耐寒性有着十分紧密的联系[31]。有研究显示,Hsp 家族热激蛋白等的引导使得可溶性蛋白会出现一定的改变,在植物响应低温胁迫信号过程中具有极为关键的作用 [32]。而可溶性蛋白含量的提升可促使下游物质合成进而为植物响应低温胁迫给予相应的物质以及能量,并有效地提升在低温下的生存率[33]。在低温环境下,糖类代谢往往是用于调控生物合成以及蛋白质分解间的平衡,可溶性蛋白的水解要有非常多的蛋白水解酶如Clp蛋白酶和以及ATP 蛋白酶,从而让多肽不可逆转地去生成可溶性糖[34]。本研究表明,4 种蔷薇属植物可溶性蛋白含量在低温胁迫中表现出不断增加的态势,抗寒性越强的材料在接收到外界环境的低温信号下,对应的蛋白质增加幅度以及分解糖的效率会更高,这就意味着抗寒性越高的材料在接收到冷信号后可以迅速地提高细胞液浓度,并不断把蛋白质转变成糖,经过可溶性蛋白去带动相应的生理反应,可溶性蛋白的转移对其余物质的合成有着十分关键的作用,使植物可以快速进行冷应激反应,并有效地适应低温环境,这和以往杜鹃[35]、鸢尾[36]、木兰[25]等植物的研究结果是一致的。
植物在低温环境中可以经过降低游离Pro 含量,有效地提升自身抗寒性能[37]。在低温胁迫之后,游离Pro 经过对蛋白质的水合作用, 能够有效地确保其细胞空间结构不会受到不利影响,从而完成对植物的有效保护[38]。XIN 等[39]对拟南芥的研究表明,高浓度脯氨酸是抗冻性提升的一个核心因素。在本研究中,4 种蔷薇属植物的Pro 含量伴随温度的下降表现出不断增加的态势,LT50 较低的S1和S2 其Pro 峰值位于-20 ℃左右,同时在低温时期其游离脯氨酸含量要明显的超过其余蔷薇属植株,这和李瑞雪等[25]所研究的低温环境下植物生理行为变化的结论一致,LT50 产生时间越晚,植物所累积的Pro 就越多,耐寒性能就越高,但随着低温胁迫的不断加大,酶系统遭到破坏,叶片Pro 合成速率下降或者其降解速率提高,进而会使得Pro 含量出现降低的变化。
植物在低温情况下,经过生成可溶性糖含量来提升细胞组织的渗透势,从而让结冰点降低,避免过度脱水,免受低温影响[40]。本研究表明,伴随温度的不断下降,4 种蔷薇属植物的SS 含量表现为不断提高,这与在玉米、番茄[41]和甘蔗[42]上研究结论是相同的。LT50 最低的S1 对应的SS 含量要明显超过其余蔷薇属植物,抗寒性最高;第二为S2,而S4 的SS 最小,这意味着LT50 越低的品种低温所生成的SS 越多,抗寒性能越高,这和李瑞雪等[25]在木兰科植物上得到的研究结论相同。
3.3 叶片丙二醛和抗氧化酶活性与抗寒性
低温胁迫能够让胞内活性氧(ROS)迅速生成,从而诱导膜脂过氧化,并进一步导致丙二醛(MDA)不断积累[43]。CAMPOS 等[44]研究表明,可将MDA 含量当做判断植物受到低温伤害程度的一个关键指标[45]。在本研究中,4 种蔷薇属植物叶片MDA 含量伴随温度的下降表现出持续提高的趋势,可见植物膜质过氧化程度逐渐加重,LT50 越低的植物对应叶片MDA 含量提升的迅速越慢。LT50最低的S1 叶片在低温阶段MDA 一直保持在一个比较低的水平,这表明S1 在低温胁迫下膜脂过氧化水平比较弱,也就是其抗寒性能较强。并且MDA增幅的程度和4 种植物抗寒性能具有负相关关系,这和邓仁菊等[46]、WANG 等[47]和朱晓非等[22]得出的结论是相似的。SOD 以及POD 能够有效去除氧自由基,从而维护植株的正常生理功能,并提高它们的抗寒能力[48]。此外,4 种蔷薇属植物整体表现出提升的变化趋势,SOD 以及POD 活性的提升能够有效地降低低温环境对生物膜的损害,这意味着植株对低温会出现相应的响应。不过MDA 含量的提升表明伴随温度的持续下降,叶片膜脂过氧化水平也越来越高。在-5~-10 ℃情况下,LT50 低的S1以及S2 叶片SOD、POD 活性较其余2 种蔷薇属植物更小,可能由于植株自身具有抵抗低温的功能,并不需要凭借保护酶去提高对植株的保护。伴随温度的不断下降,S1 以及S2 叶片中SOD、POD 活性被激活从而防御低温伤害。这表明重度低温胁迫会阻碍植株的抗氧化酶活性,进而让细胞遭受损伤,在LT50 范围内SOD 以及POD 活性的提升能够有效地防止低温对细胞带来的损害。
3.4 抗寒相关调节基因HSP90 与抗寒性
低温是干扰植物生长发育的一个重要因素,植物具备不同的防御机制去抵御寒冷的伤害[47,49-50]。通过转录因子介导的信号传导是最高效的防御应答反应之一,组成了植物复杂调控网络的关键部分[51]。很多研究显示,各种热休克蛋白HSPs 转录因子在温度感知以及信号处理中有着不可替代的关键作用,热激转录因子HSP 在逆境胁迫下和热激元件(HSE)识别并进行组合,进而带动下游基因HSP 的转录以及表达[52],对植物抵御逆境伤害以及其余生理行为有着重要意义。HSP90 为HSPs 的一部分,是类固醇受体的重要部分,可以促使目标蛋白的积累以及激酶的激活,整合质体信号通路,调节CBF 和COR 表达[53]。有研究显示,低温环境可以有效地促使HSP90 基因表达,从而调整拟南芥幼苗的生长发育[47]。在低温环境中,经过促使HSP 基因过量表达去去除多余的活性氧,进而防止氧化胁迫带来的影响[54]。本研究中,HSP90 基因相对表达量随着低温胁迫温度的降低,整体表现出不断增加的态势,这种表达方式和SP、Pro、SOD、POD 在 4 种蔷薇属植株中的变化基本一致,意味着HSP90 会参加到目标蛋白的积累过程中。在0 ℃下,HSP90 基因在4 种植株间的表达量差异不太明显,伴随低温胁迫的不断加大,该基因在叶片中的表达也不断提高。低温时,耐寒种质S1 以及S2 中HSP90 的表达量一直都位于最高。这意味着4 种蔷薇属植物抗寒性强度不同对低温的应答机制也就有所不同,耐寒性越高的植株为防范低温带来的损伤,会更加迅速地开启低温应答机制,激活HSP90 进行表达,进一步让下游和耐寒有关的基因完成表达,提高ROS 清除力度,降低氧化胁迫带来的影响[55],这种随低温胁迫时间不断增加,转录因子表达越来越强的状况在短枝木麻黄[26]和木兰科植物[25]等研究中也有类似的结论。
参考文献:
[1] 许凤,张颢,杨春梅,等. 利用隶属函数值法评价月季耐寒性
[J]. 西南农业学报,2012,25(5):1870-1873.
XU F,ZHANG H,YANG C M,et al. Evaluation of coldresistance
of rose species using membership function[J]. Southwest
China Journal of Agricultural Sciences,2012,25(5):1870-
1873.
[2] 文建雷,薛智德,胡景江. 玫瑰的抗寒性與质膜透性[J]. 西北林
学院学报,2000,15(4):16-20.
WEN J L,XUE Z D,HU J J. Cold resistance and permeability
of roses[J]. Journal of Northwest Forestry College,2000,15(4):
16-20.
[3] 张涛,段大娟,王振一,等. 5 种藤本月季抗寒性比较研究[J]. 西
北林学院学报,2006,21(5):81-83,138.
ZHANG T,DUAN D J,WANG Z Y,et al. Comparison of
freezing resistance among five climbing rose varieties[J]. Journal
of Northwest Forestry University,2006,21(5):81-83,138.
[4] 周家杏,曾丽,陶懿伟,等. 微型月季花粉生活力测定方法的研
究[J]. 上海交通大学学报(农业科学版),2007,25(6):574-577.
ZHOU J X,ZENG L,TAO Y W,et al. Studies on measurement
methods of pollen viability of Rosa chinensis var.minima?s
[J]. Journal of Shanghai Jiao Tong University(Agricultural Science),
2007,25(6):574-577.
[5] 孫佳,曾丽,刘正宇,等. 微型月季品种分类的花粉形态学[J].
中国农业科学,2009,42(5):1867-1874.
SUN J,ZENG L,LIU Z Y,et al. Classification of Rosa hybrida
var.minima cultivars by pollen morphology[J]. Scientia Agricultura
Sinica,2009,42(5):1867-1874.
[6] 何利刚,蒋迎春,吴黎明,等. 几种柑橘资源的抗寒性测定及初
步评价[J]. 农业科技通讯,2015(5):176-178.
HE L G,JIANG Y C,WU L M,et al. Determination and preliminary
evaluation of cold resistance of several citrus resources
[J]. Bulletin of Agricultural Science and Technology,2015(5):
176-178.
[7] YANG Y,YAO N,JIA Z K,et al. Effect of exogenous abscisic
acid on cold acclimation in two Magnolia species[J]. Biologia
Plantarum,2016,60(3):555-562.
[8] PEARCE R S. Plant freezing and damage[J]. Annals of Botany,
2001,87(4):417-424.
[9] WISNIEWSKI M,BASSETT C,ARORA R. Distribution and
partial characterization of seasonally expressed proteins in different
aged shoots and roots of‘ Loring peach(Prunus persica)[J].
Tree Physiology,2004,24(3):339-345.
[10] LI W H,LI L F,TING M F,et al. Intensification of Northern
Hemisphere subtropical highs in a warming climate[J]. Nature
Geoscience,2012,5(11):830-834.
[11] MOELLERING E R,MUTHAN B,BENNING C. Freezing
tolerance in plants requires lipid remodeling at the outer chloroplast
membrane[J]. Science,2010,330:226-228.
[12] JENA K K,KIM S M,SUH J P,et al. Identification of coldtolerant
breeding lines by quantitative trait loci associated with
cold tolerance in rice[J]. Crop Science,2012,52(2):517-523.
[13] WINFIELD M O,LU C G,WILSON I D,et al. Plant responses
to cold:Transcriptome analysis of wheat[J]. Plant Biotechnology
Journal,2010,8(7):749-771.
[14] 刘杜玲,张博勇,孙红梅,等. 早实核桃不同品种抗寒性综合
评价[J]. 园艺学报,2015,42(3):545-553.
LIU D L,ZHANG B Y,SUN H M,et al. Comprehensive
evaluation on cold resistance of early fruiting walnut cultivars
[J]. Acta Horticulturae Sinica,2015,42(3):545-553.
[15] SKINNER D Z,GARLAND-CAMPBELL K A. The relationship
of LT50 to prolonged freezing survival in winter wheat[J].
Canadian Journal of Plant Science,2008,88(5):885-889.
[16] BAXTER A,MITTLER R,SUZUKI N. ROS as key players
in plant stress signalling[J]. Journal of Experimental Botany,
2014,65(5):1229-1240.
[17] CHEN W,LI D S,ZHANG M,et al. Cloning and differential
expression of five heat shock protein genes associated with thermal
stress and development in the polyphagous predatory mite
Neoseiulus cucumeris(Acari:Phytoseiidae)[J]. Experimental
and Applied Acarology,2015,67(1):65-85.
[18] WU Y K,ZOU C,FU D M,et al. Molecular characterization
of three Hsp90 from Pieris and expression patterns in response
to cold and thermal stress in summer and winter diapause of
Pieris melete[J]. Insect Science,2018,25(2):273-283.
[19] FOWLER S,THOMASHOW M F. Arabidopsis transcriptome
profiling indicates that multiple regulatory pathways are
activated during cold acclimation in addition to the CBF cold response
pathway[J]. The Plant Cell,2002,14(8):1675-1690.
[20] XIONG H C,GUO H J,XIE Y D,et al. RNAseq analysis reveals
pathways and candidate genes associated with salinity tolerance
in a spaceflight-induced wheat mutant[J]. Scientific Reports,
2017,7:2731.
[21] XU W R,LI R M,ZHANG N B,et al. Transcriptome profil‐
ing of Vitis amurensis,an extremely cold-tolerant Chinese wild
Vitis species,reveals candidate genes and events that potentially
connected to cold stress[J]. Plant Molecular Biology,
2014,86(4):527-541.
[22] 朱曉非,曾丽,刘洋,等. 微型月季低温处理后生理指标变化
及其耐寒性评价[J]. 上海交通大学学报(农业科学版),2017,
35(4):21-25,30.
ZHU X F,ZENG L,LIU Y,et al. Changes of physiological indexes
and evaluation of cold tolerance of miniature rose after
low temperature treatment[J]. Journal of Shanghai Jiao Tong
University(Agricultural Science),2017,35(4):21-25,30.
[23] 刘峰,李豪,王安,等. 14 种切花月季抗寒性比较与评价[J]. 湖
北农业科学,2018,57(7):75-79.
LIU F,LI H,WANG A,et al. Comparison and evaluation on
cold-tolerance of fourteen cut rose cultivars[J]. Hubei Agricultural
Sciences,2018,57(7):75-79.
[24] 陈建勋,王晓峰. 植物生理学实验指导[M]. 2 版. 广州:华南
理工大学出版社,2006.
CHEN J X,WANG X F. Experimental guidance of plant
physiology[M]. 2nd Edition. Guangzhou:South China University
of Technology Press,2006.
[25] 李瑞雪,金晓玲,胡希军,等. 低温胁迫下6 种木兰科植物的生
理响应及抗寒相关基因差异表达[J]. 生态学报,2019,39(8):
2883-2898.
LI R X,JIN X L,HU X J,et al. Physiological responses and
differential expression of cold resistance-related genes of six varieties
of Magnoliaceae under low temperature stress[J]. Acta
Ecologica Sinica,2019,39(8):2883-2898.
[26] 李楠,郑勇奇,丁红梅,等. 低温胁迫下短枝木麻黄耐寒相关
基因的差异表达分析[J]. 林业科学,2017,53(7):62-71.
LI N,ZHENG Y Q,DING H M,et al. Analysis on differential
expression of cold resistance related genes of Casuarina equi?
setifolia under low temperature stress[J]. Scientia Silvae Sinicae,
2017,53(7):62-71.
[27] 刘艳阳,李俊周,陈磊,等. 低温胁迫对小麦叶片细胞膜脂质
过氧化产物及相关酶活性的影响[J]. 麦类作物学报,2006,26
(4):70-73.
LIU Y Y,LI J Z,CHEN L,et al. Effect of low temperature
stress on peroxidation product of membrane lipids and activity
of related enzymes in wheat seedling leaves[J]. Journal of Triticeae
Crops,2006,26(4):70-73.
[28] NASEF I N. Short hot water as safe treatment induces chilling
tolerance and antioxidant enzymes,prevents decay and maintains
quality of cold-stored cucumbers[J]. Postharvest Biology
and Technology,2018,138:1-10.
[29] 令凡,焦健,李朝周,等. 不同油橄欖品种对低温胁迫的生理
响应及抗寒性综合评价[J]. 西北植物学报,2015,35(3):
508-515.
LING F,JIAO J,LI C Z,et al. Physiological response and
comprehensive evaluation of cold resistance under cold stress
for different varieties of Olea europaea[J]. Acta Botanica
Boreali-Occidentalia Sinica,2015,35(3):508-515.
[30] 任俊杰,赵爽,李美美,等. 不同核桃品种抗霜冻能力的评价
[J]. 林业科学,2014,50(9):67-72.
REN J J,ZHAO S,LI M M,et al. Evaluation of frost resistance
ability of different walnut cultivars[J]. Scientia Silvae
Sinicae,2014,50(9):67-72.
[31] 姚海梅,李永生,张同祯,等. 旱—盐复合胁迫对玉米种子萌
发和生理特性的影响[J]. 应用生态学报,2016,27(7):2301-
2307.
YAO H M,LI Y S,ZHANG T Z,et al. Effects of combined
drought and salinity stress on germination and physiological
characteristics of maize(Zea mays)[J]. Chinese Journal of Applied
Ecology,2016,27(7):2301-2307.
[32] HEIDARVAND L,MILLAR A H,TAYLOR N L. Responses
of the mitochondrial respiratory system to low temperature
in plants[J]. Critical Reviews in Plant Sciences,2017,36
(4):217-240.
[33] 王丹,宣继萍,朱小晨,等. 假俭草抗寒性与体内碳水化合物、
脯氨酸、可溶性蛋白含量的关系[J]. 草地学报,2010,18(6):
816-822.
WANG D,XUAN J P,ZHU X C,et al. Relationships of freezing
tolerance and the contents of carbohydrates,proline,protein
in centipedegrass(Eremochloa ophiuroides(Munro.) hack.)[J].
Acta Agrestia Sinica,2010,18(6):816-822.
[34] WANG J M,YANG Y,LIU X H,et al. Transcriptome profiling
of the cold response and signaling pathways in Lilium lanci?
folium[J]. BMC Genomics,2014,15(1):203.
[35] 刘冰,曹莎,周泓,等. 杜鹃花品种耐寒性比较及其机制研究
[J]. 园艺学报,2016,43(2):295-306.
LIU B,CAO S,ZHOU H,et al. A study on cold tolerance difference
and its mechanisms in Azalea cultivars[J]. Acta Horticulturae
Sinica,2016,43(2):295-306.
[36] 王冠群,李丹青,張佳平,等. 德国鸢尾6 个品种的耐寒性比较
[J]. 园艺学报,2014,41(4):773-780.
WANG G Q,LI D Q,ZHANG J P,et al. Comparison of cold
tolerance within 6 cultivars of Iris germanica[J]. Acta Horticulturae
Sinica,2014,41(4):773-780.
[37] 岳海,李国华,李国伟,等. 澳洲坚果不同品种耐寒特性的研
究[J]. 园艺学报,2010,37(1):31-38.
YUE H,LI G H,LI G W,et al. Studies on cold resistance of
different Macadamia cultivars[J]. Acta Horticulturae Sinica,
2010,37(1):31-38.
[38] 王兆. 低温胁迫对彩叶草的生理效应及抗寒性研究[D]. 福
州:福建农林大学,2014.
WANG Z. Physiological responses to chilling stress in Solenostemon
scutellarioides and its inductive research[D]. Fuzhou:
Fujian Agriculture and Forestry University,2014.
[39] XIN Z,BROWSE J. Eskimol mutants of Arabidopsis are constitutively
freezing-tolerant[J]. Proceedings of the National
Academy of Sciences of the United States of America,1998,95
(13):7799-7804.
[40] POCIECHA E, DZIURKA M. Trichoderma interferes with
cold acclimation by lowering soluble sugars accumulation resulting
in reduced pink snow mould(Microdochium nivale) resistance
of winter rye[J]. Environmental and Experimental
Botany,2015,109:193-200.
[41] 简令成,卢存福,李积宏,等. 适宜低温锻炼提高冷敏感植物
玉米和番茄的抗冷性及其生理基础[J]. 作物学报,2005,31
(8):971-976.
JIAN L C,LU C F,LI J H,et al. Increment of chilling tolerance
and its physiological basis in chilling-sensitive corn
sprouts and tomato seedlings after cold-hardening at optimum
temperatures[J]. Acta Agronomica Sinica,2005,31 (8) :
971-976.
[42] 张保青,杨丽涛,李杨瑞. 自然条件下甘蔗品种抗寒生理生化
特性的比较[J]. 作物学报,2011,37(3):496-505.
ZHANG B Q,YANG L T,LI Y R. Comparison of physiological
and biochemical characteristics related to cold resistance in
sugarcane under field conditions[J]. Acta Agronomica Sinica,
2011,37(3):496-505.
[43] 王志昊,叶冬梅,何炎红,等. 5 种沙生植物丙二醛、脯氨酸和2
种氧化物酶比较[J]. 分子植物育种,2018,16(11):3727-3731.
WANG Z H,YE D M,HE Y H,et al. Comparison of the content
of MDA,proline and activity of two kinds of enzyme in 5
common desert plants[J]. Molecular Plant Breeding,2018,16
(11):3727-3731.
[44] CAMPOS P S, QUARTIN V N, RAMALHO J C, et al.
Electrolyte leakage and lipid degradation account for cold sensitivity
in leaves of Coffea sp.plants[J]. Journal of Plant Physiology,
2003,160(3):283-292.
[45] 叶艳然,王文莉,郑成淑,等. 四种野生苔草属植物的耐寒性
评价[J]. 应用生态学报,2017,28(1):89-95.
YE Y R,WANG W L,ZHENG C S,et al. Evaluation of cold
resistance of four wild Carex speices[J]. Chinese Journal of Applied
Ecology,2017,28(1):89-95.
[46] 邓仁菊,范建新,王永清,等. 低温胁迫下火龙果的半致死温
度及抗寒性分析[J]. 植物生理学报,2014,50(11):1742-1748.
DENG R J,FAN J X,WANG Y Q,et al. Semilethal temperature
of pitaya under low temperature stress and evaluation on
their cold resistance[J]. Plant Physiology Journal,2014,50
(11):1742-1748.
[47] WANG R H,ZHANG Y,KIEFFER M,et al. HSP90 regulates
temperature-dependent seedling growth in Arabidopsis by
stabilizing the auxin co-receptor F-box protein TIR1[J]. Nature
Communications,2016,7:10269.
[48] AM?GLIO T,PIGEON D,ARCHILLA O,et al. Adaptation
to cold temperature and response to freezing in roses[J]. Acta
Horticulturae,2003,618:515-520.
[49] 王明霞,梅超,宋倩娜,等. 低溫胁迫下6 种马铃薯组培苗耐寒
性评价[J]. 山西农业科学,2021,49(12):1502-1506.
WANG M X,MEI C,SONG Q N,et al. Evaluation on cold
tolerance of six potato seedlings in tissue culture under low
temperature stress[J]. Journal of Shanxi Agricultural Sciences,
2021,49(12):1502-1506.
[50] 方宇辉,韩留鹏,华夏,等. 小麦抗寒性研究进展[J]. 河南农业
科学,2022,51(4):1‐10.
FANG Y H,HAN L P,HUA X,et al. Research progress on
cold resistance of wheat[J]. Journal of Henan Agricultural Sciences,
2022,51(4):1‐10.
[51] ASHRAF M. Inducing drought tolerance in plants:recent advances[
J]. Biotechnology Advances,2010,28(1):169-183.
[52] PELHAM H R B. A regulatory upstream promoter element in
the Drosophila Hsp 70 heat-shock gene[J]. Cell,1982,30(2):
517-528.
[53] NOREN L,KINDGREN P,STACHULA P,et al. HSP90,
ZTL, PRR5 and HY5 integrate circadian and plastid signaling
pathways to regulate CBF and COR expression[J]. Plant Physiology,
2016,177(2):1392-1406.
[54] PARK S,LEE C M,DOHERTY C J,et al. Regulation of the
Arabidopsis CBF regulon by a complex low-temperature regulatory
network[J]. The Plant Journal,2015,82(2):193-207.
[55] 楊帆,丁菲,杜天真. 盐胁迫下构树DREB 转录因子基因表达
的实时荧光定量PCR 分析[J]. 林业科学,2010,46(4):
146-150.
YANG F,DING F,DU T Z. DREB gene expression in leaves
of Broussonetia papyrifera seedlings under salt stress detected
by real-time fluorescent quantitative PCR[J]. Scientia Silvae
Sinicae,2010,46(4):146-150.
