低温对不同萌发状态裸燕麦种子生长生理特性的影响
2016-07-25张丽贾志国
张丽++贾志国
摘要:将3种萌发状态(吸胀、萌动、发芽)的裸燕麦种子分别于0 ℃下处理24、48、72 h,测定裸燕麦种子发芽及各项生长、生理指标。结果表明:随低温胁迫时间的延长,发芽率、发芽势、发芽指数及活力指数均显著降低。与对照相比,低温胁迫显著抑制了根长、芽长、鲜质量、干质量及根冠比,其中48 h时间处理下,根长、鲜质量、干质量及根冠比受抑制程度最小。各处理下的相对电导率、丙二醛含量及可溶性蛋白含量表现为不同程度的增加,脯氨酸含量显著降低。综上所述,裸燕麦对24、48 h的低温处理有一定的忍耐能力,3种萌发状态的裸燕麦种子抗寒能力从强到弱依次为发芽种子> 吸胀种子> 萌动种子。
关键词:裸燕麦;低温胁迫;萌发;生理指标;抗寒性
中图分类号: S512.601文献标志码: A文章编号:1002-1302(2016)06-0161-04
收稿日期:2015-04-23
基金项目:河北北方学院自然科学研究计划。
作者简介:张丽(1979—),女,博士,讲师,从事作物与经济林栽培育种工作。E-mail:yingrizl@126.com。裸燕麦(Avena nuda L.)别称莜麦、铃铛麦,禾本科燕麦属一年生草本植物[1]。裸燕麦性喜凉爽,耐干旱、抗盐碱、生长周期短,是我国华北、西北等高寒地区的主要粮饲兼用作物[2],其营养价值高,富含人体必需的8种氨基酸,是北方地区人们喜爱的特色食物,其秸秆也是优质的牧草[3]。近年来研究发现,裸燕麦具有降脂、降糖的功效,被作为一种营养保健功能食品加以开发利用[4],因此裸燕麦是一种多用途、高经济价值的作物。
我国海拔较高的高寒地区为裸燕麦的适宜种植区。裸燕麦是禾本科中抗寒性较强的作物,萌芽期是其生长发育过程中最关键的时期,此时由于其抗寒能力强,能抵抗一定的低温胁迫,但当温度过低,超过其所能忍受的极限低温时便会造成胁迫,影响种子发芽率和发芽势,造成出苗不齐,从而影响其最终产量。前人关于裸燕麦抗旱性研究较多[5-7],对其抗寒机理的研究较少。本试验研究了同一低温不同处理时间胁迫对不同萌发状态裸燕麦种子的影响,从萌发、生长、生理3个方面探讨了裸燕麦对低温胁迫的响应,揭示了裸燕麦抗寒生理机制,为我国北方严寒地区裸燕麦高产栽培及进一步推广提供理论依据。
1材料与方法
1.1试验材料与设计
选择裸燕麦常规品种花早2号为试材。挑选大小一致、籽粒饱满的种子,分成36组,每组100粒,经清水浸种24 h后,在20 ℃进行萌发试验,将种子处理为吸胀、萌动、发芽3种萌发状态,分别进行低温胁迫发芽试验。将不同萌发状态的裸燕麦种子置于0 ℃低温下,分别处理24、48、72 h,3个时间处理低温胁迫结束后,转移至20 ℃继续培养;对照始终保持20 ℃。重复3次。
1.2试验及测定方法
1.2.1发芽试验方法种子低温胁迫萌发试验根据颜启传的方法[8]进行改进。取足量种子放入大烧杯中浸泡24 h,然后放入经过消毒的大瓷盘中,用湿布覆盖,放在阴蔽处培养,供试验选用。首先于15 cm培养皿内铺2层滤纸作为发芽床,选取浸泡后的均匀饱满种子12组,每组100粒,均匀摆放,作为吸胀组试验材料。将培养皿置于0 ℃光照培养箱分别处理24、48、72 h,每天补充适量的蒸馏水,以保持渗透势不变,分别测定各项生理指标。随着试验的进行,再于瓷盘中分别挑选处于萌动和发芽状态的种子进行上述试验,每处理重复3次。
1.2.2发芽指标的测定从上述3种萌发状态的种子中选择萌动处理组为代表,进行标准发芽试验,每天记录发芽数,于试验后4 d计算发芽势,8 d后试验结束,计算发芽率、发芽指数及活力指数。
1.2.3生长指标的测定将“1.2.2”节发芽结束后的各处理在20 ℃下培养15 d,利用常规方法测定裸燕麦幼苗生长初期根长、苗高、根鲜质量、苗鲜质量、干质量,计算根冠比。
1.2.4生理指标的测定相对电导率采用电导率法测定。称取0.5 g样品,加入10 mL水,浸泡24 h,测定其浸出液电导率,煮沸15 min,冷却后再次测定其电导率,将2次电导率代入公式,记算结果;膜伤害指标丙二醛(MDA)含量采用硫代巴比妥酸(TBA)比色法[9]测定;游离脯氨酸含量采用酸性茚三酮显色法[9]测定;可溶性蛋白含量采用考马斯亮蓝 G-250 染色法[10]测定。
1.3数据分析
应用Microsoft Excel软件进行数据整理和绘图,SPSS 160软件进行数据统计分析,用单因素方差分析(One-Way ANOVA)和最小显著差异法(LSD)比较不同处理间的差异。
2结果与分析
2.1低温胁迫对裸燕麦种子发芽的影响
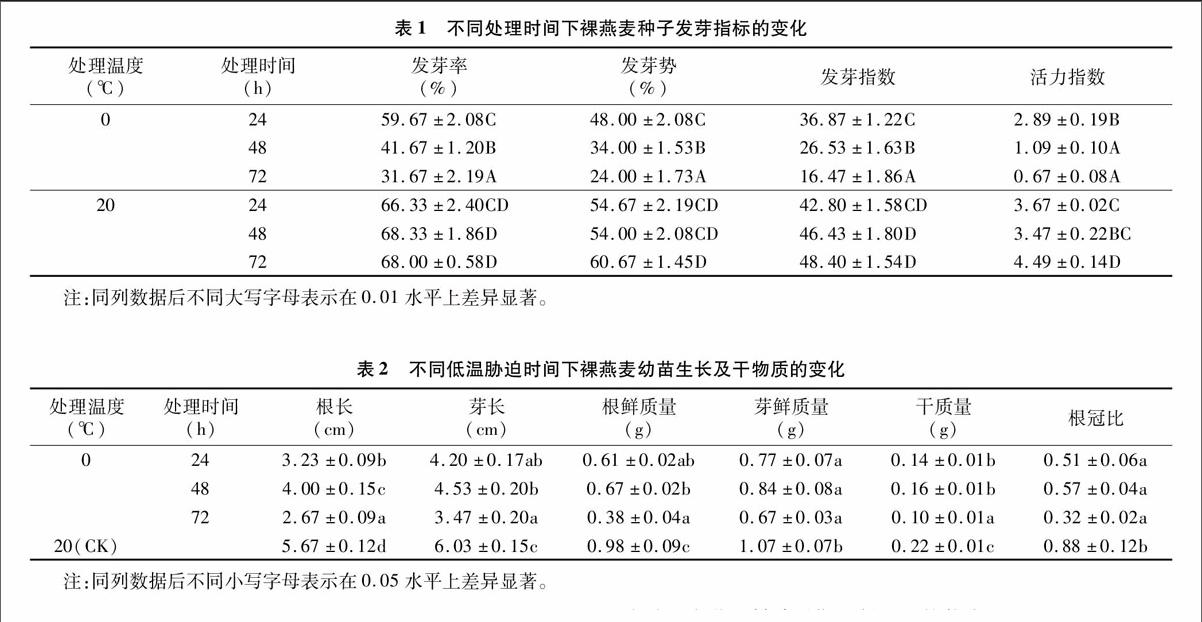
由表1可以看出,随着低温胁迫时间的延长,裸燕麦种子的发芽率、发芽势、发芽指数及活力指数均呈现逐渐降低的趋势,与20 ℃对照相比差异显著。24、48、72 h的低温胁迫后发芽率分别降低为对照的90.0%、61.0%、46.6%,且处理间差异显著。发芽势的变化规律与发芽率基本一致,72 h的低温胁迫导致发芽势降低为对照的39.6%,表明相比发芽率低温胁迫对发芽势的影响更大。24 h的低温处理也显著降低了发芽指数。活力指数则表现为极显著差异(P<0.01),随着胁迫时间的延长,48、72 h胁迫下,发芽指数和活力指数分别降低为对照的57.1%、34.0%和31.4%、14.9%,说明低温胁迫一定程度上抑制了裸燕麦的萌发,活力指数的变化可以看出即使能够萌发的那部分种子,其继续生长的活力也受到了明显的影响。
2.2低温胁迫对裸燕麦幼苗苗高、根长、鲜质量、干物质及根冠比的影响
温度是植物种子萌发出苗的基本条件之一,低温胁迫导致萌动发芽后的种子生长缓慢甚至停止生长[11]。本研究结果(表2)表明,裸燕麦幼苗的根长、芽长均受到了低温抑制,与对照相比均显著降低,随着低温胁迫时间的延长,根长表现为先升后降的趋势,48 h的低温胁迫反而使根长增加,与24 h处理相比,48 h处理下芽长变化不显著,72 h的低温处理对芽长的抑制作用进一步加强。3种低温时间处理下,根鲜质量显著低于对照,24、72 h低温处理后的芽鲜质量也显著降低,相比而言,低温胁迫对根的抑制作用更明显。随低温胁迫时间的增加,48 h处理后的根鲜质量和芽鲜质量均略有增加,72 h处理又继续下降,但处理间差异不显著。裸燕麦幼苗干物质受到低温的显著抑制,0 ℃处理72 h后的种子萌发后的幼苗干质量仅为对照的45.5%。与对照相比,根冠比与干质量的变化规律基本一致,均低于对照,72 h处理后降低为对照的36.4%,3个时间处理之间对比发现,48 h处理后的干质量和根冠比的平均值最大,与对照不显著,表明裸燕麦对低温胁迫有一定的适应抵抗能力。
2.3不同低温胁迫时间对裸燕麦种子生理指标的影响
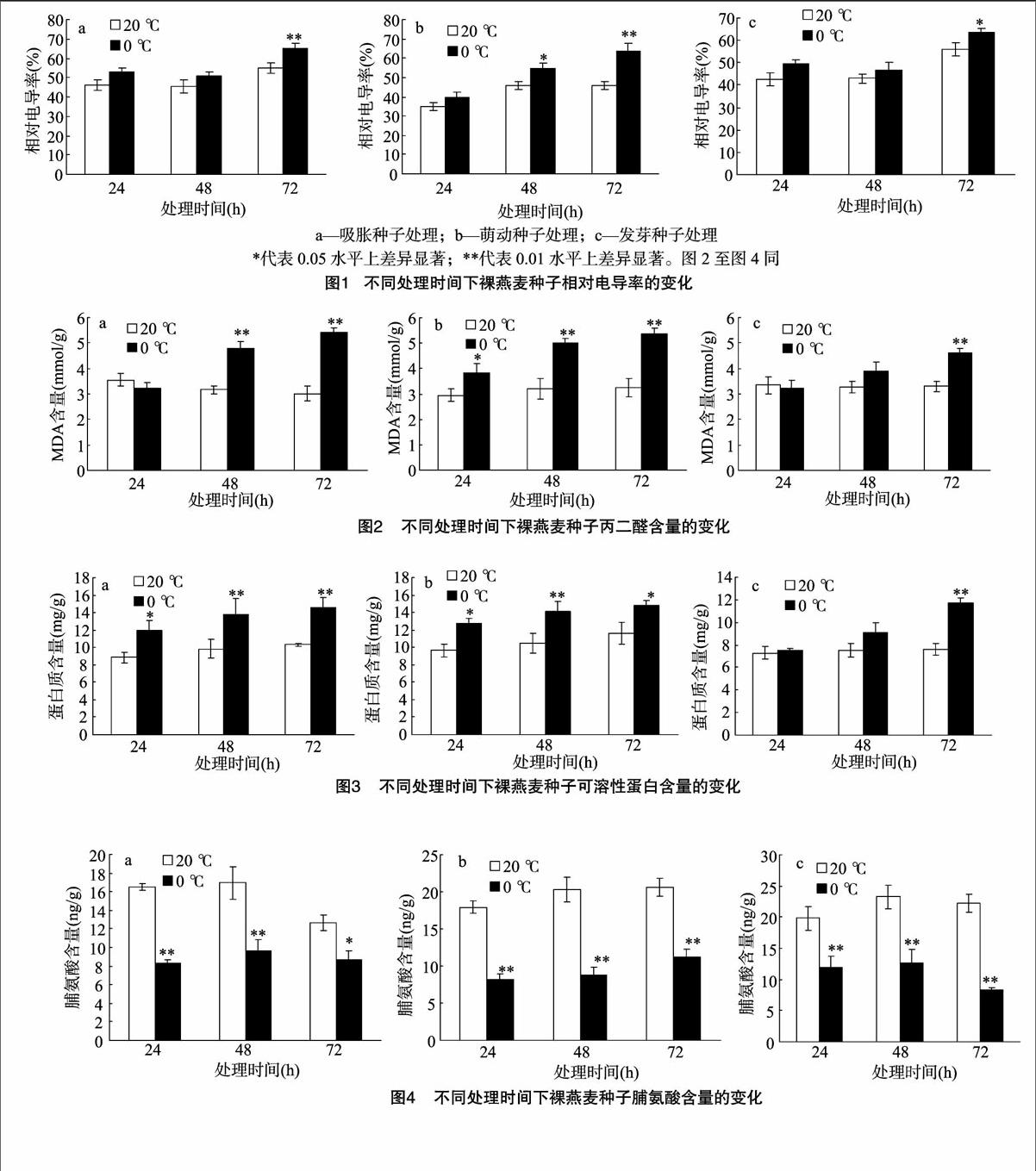
2.3.1不同低温胁迫时间下裸燕麦种子相对电导率的变化由图1可知,与对照相比,低温处理后不同萌发状态的裸燕麦种子相对电导率均有所增加。图1-a所示低温处理72 h的吸胀种子相对电导率极显著增加,而24、48 h处理下增加不明显。随着0 ℃低温胁迫时间的延长,萌动种子相对电导率逐渐增加(图1-b),与各自对照相比,48 h处理达显著差异,72 h处理差异极显著。发芽种子在低温处理72 h后也显著增加(图1-c)。吸胀、萌动、发芽3种萌发状态的裸燕麦种子72 h处理后电导率的相对增加量分别为10.2%、18.1%、7.6%,表明发芽种子抗低温能力较强,而萌动种子作为种子发芽的敏感时期对低温抵抗能力最弱。
2.3.2不同低温胁迫时间下裸燕麦种子丙二醛含量的变化植物在逆境胁迫过程中产生过剩的自由基,引发膜脂过氧化作用,膜脂过氧化的中间产物自由基和最终产物MDA都会造成膜系统的损伤[12]。由图2可以看出,随低温处理时间的延长,不同萌发状态的裸燕麦种子丙二醛含量也会增加。与各自对照相比,48、72 h处理后的吸胀和萌动种子差异极显著(图2-a、图2-b)。图2-c所示,24、48 h的低温处理时间下发芽状态种子的丙二醛含量差异不显著,当胁迫时间延长至72 h时其含量显著增加。24 h低温胁迫后吸胀和发芽种子丙二醛含量无明显变化,说明裸燕麦对短时间的低温胁迫有一定的忍耐能力。
2.3.3不同低温胁迫时间下裸燕麦种子可溶性蛋白含量的变化可溶性蛋白作为渗透调节物质、能量物质和信息传递物质在植物抵御逆境过程中起着重要的作用[13]。由图3可知,经过低温胁迫处理的裸燕麦种子中可溶性蛋白含量均高于对照,说明低温胁迫造成种子中可溶性蛋白含量升高。3种低温处理时间下,处于吸胀和萌动时期的裸燕麦种子可溶性蛋白含量均显著高于对照(图3-a、图3-b)。发芽状态的裸燕麦种子在处理时间为72 h时,可溶性蛋白含量显著高于对照,24、48 h处理比对照略有增加,但差异不显著。72 h 低温胁迫处理下,吸胀、萌动、发芽3种萌发状态可溶性蛋白增加量分别为各自对照的1.41、1.24、1.54倍。萌动时期蛋白增加量最低,表明此时期的裸燕麦种子对低温更敏感,抗寒性较弱。
2.3.4不同低温胁迫时间下裸燕麦种子脯氨酸含量的变化由图4可知,与各自对照相比,3种萌发状态下的裸燕麦种子低温胁迫后的脯氨酸含量均显著降低。不同萌发状态种子处理之间对比发现,24、48 h低温胁迫后,发芽状态下脯氨酸积累最多,其平均值分别为吸胀、萌动种子的1.37、1.46倍,表明发芽状态的裸燕麦种子在一定时间的低温胁迫范围内,脯氨酸可以迅速积累以抵抗逆境条件。
3结论与讨论
各种作物种子对发芽要求一定的温度,过低的温度会抑制种子的萌发[14-15]。本研究中低温胁迫抑制了裸燕麦种子萌发,随着低温处理时间的延长,发芽势逐渐下降,发芽总数也显著减少。不同低温处理时间下裸燕麦种子的发芽率、发芽势、发芽指数及活力指数均低于对照,但活力指数下降的最快,表明低温胁迫在一定程度上抑制了萌发,而仍能萌发的那部分种子活力也大大降低了。裸燕麦芽苗期的生长指标也受到低温的影响,持续低温阻碍了芽和根的伸长,48 h低温处理后的各生长指标的平均值均高于其他2种时间处理,表明恢复正常温度后,裸燕麦仍能正常生长,低温处理48 h后的种子在适宜条件下生长最快,处理裸燕麦对持续低温48 h有一定的忍耐能力,温度回升后尚能恢复,但当低温胁迫时间持续超过72 h,种子发芽生长已比较困难,只能勉强维持生命活动。本试验还发现,24 h低温胁迫下的生长恢复能力低于 48 h 处理,对于抗寒性较强的裸燕麦来说可能存在一个低温锻炼的问题,48 h的持续低温使幼苗体内积累了更多的干物质,从而保持了一定的生长量。前人关于小麦的研究中发现,干旱胁迫虽然导致了不同萌发状态种子的伤害,但并不丧失其自身的修复和恢复生长的能力[16]。
在低温冷害条件下,植物由于细胞膜内大量溶质外渗导致了其浸出液的电导率增加,表明细胞膜遭受了破坏,电导率增加越少,表明抗寒性越强[17]。经过24、48 h低温处理的裸燕麦膜透性增加的程度较小,尤其是吸胀种子和发芽种子其数值虽略有增加,但与对照差异不显著,温度回升后靠自身的修复能力可以恢复正常。经过72 h低温处理的裸燕麦膜透性增加程度较大,细胞内电解质大量外渗,电导率显著增加。分析电导率指标得出,裸燕麦可以忍耐24、48 h的持续低温,且发芽种子的耐寒能力大于吸胀种子和萌动种子。低温处理后,3种不同萌发状态的裸燕麦种子丙二醛、可溶性蛋白含量表现为不同程度的增加,经分析得出与电导率指标基本一致的结果,其抗寒能力从强到弱依次为发芽种子>吸胀种子>萌动种子。萌动状态的种子抵御低温胁迫的能力较差,该时期对外界环境条件非常敏感,各种生理活性物质及代谢反应开始趋于旺盛,持续低温干扰了萌发的进程,其受伤害程度也较大。
脯氨酸含量作为抗寒性指标前人已有大量报道[18-19]。一般认为,脯氨酸积累越多其抗寒性越强,但也有不一致的观点,认为脯氨酸与抗寒性关系不大[20]。本试验发现了与前人研究不一致的结果,各处理组脯氨酸含量均显著低于对照,可能是由于裸燕麦种子蛋白质含量丰富,20 ℃正常萌发时,蛋白质大量分解为可供幼苗利用的游离氨基酸,而0 ℃处理抑制了蛋白水解酶的活性,从而导致对照组脯氨酸含量大于处理组,抵消了低温胁迫后脯氨酸的积累作用。如果忽略对照组,单纯分析低温胁迫处理发现,发芽状态的脯氨酸积累量大于吸胀和萌动的种子,与上述指标分析结果一致。因此,脯氨酸含量可以作为裸燕麦抗寒性的指标。
综上所述,裸燕麦萌发时期抗低温能力较强,且发芽种子>吸胀种子>萌动种子。从吸胀、萌动到发芽不同萌发状态的裸燕麦种子,遇到短时间的低温胁迫后,并不会丧失恢复能力,在适宜的温度条件下仍能重新恢复生长,比延迟播种争得了更宝贵的生长时间,为裸燕麦生产中播种期的确定提供了科学依据。
参考文献:
[1]杨海鹏. 中国燕麦[M]. 北京:农业出版社,1989.
[2]郑克宽,韩冰,于海峰,等. 裸燕麦新品种的经济性状和实用价值的研究[J]. 内蒙古农业大学学报:自然科学版,2002,23(1):61-65.
[3]崔海,王加启,卜登攀,等. 燕麦饲料在动物生产中的应用[J]. 中国畜牧兽医,2010,37(6):214-217.
[4]王雁丽,林凤仙,赵英. 浅论高寒地区裸燕麦的开发及综合利用[J]. 吉林农业科学,2007,32(3):57-59.
[5]赵海超,刘景辉,任永峰,等. 水分胁迫对内农大莜1号裸燕麦品种生长指标的影响[J]. 作物杂志,2010(1):73-76.
[6]李威. 六种裸燕麦品种苗期抗旱性比较[J]. 青海大学学报:自然科学版,2008,26(4):44-48.
[7]牛瑞明,王燕,吴桂丽,等. 裸燕麦种子萌发对模拟干旱胁迫的响应及其耐旱性综合评价[J]. 麦类作物学报,2011,31(4):753-756.
[8]颜启传. 种子学[M]. 北京:中国农业出版社,2001:425-432.
[9]王学奎. 植物生理生化实验原理和技术[M]. 2版.北京:高等教育出版社,2006:278-280.
[10]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000.
[11]韩春梅. 不同温度处理对豇豆种子萌发及抗氧化酶活性的影响[J]. 长江蔬菜,2010(10):29-30.
[12]吕优伟,贺佳圆,白小明,等. 9个野生早熟禾对低温胁迫的生理响应及苗期抗寒性评价[J]. 草地学报,2014,22(2):326-333.
[13]李春燕,陈思思,徐雯,等. 苗期低温胁迫对扬麦16叶片抗氧化酶和渗透调节物质的影响[J]. 作物学报,2011,37(12):2293-2298.
[14]张荣萍,吉琼芳. 低温胁迫对粳型巨胚稻西巨胚一号种子萌发的影响[J]. 西昌学院学报:自然科学版,2014,28(4):5-7.
[15]谷英楠,杨克军,王玉凤,等. 低温对寒地玉米种子萌发及幼苗抗氧化酶活性的影响[J]. 安徽农业科学,2014,42(16):5092-5096.
[16]朱云集,崔金梅,彭文博,等. 不同萌发状态的小麦种子在干旱逆境下的生理特性及恢复能力[J]. 华北农学报,1999,14(2):73-78.
[17]王建强,邓永成. 低温胁迫对银叶菊和矾根抗寒生理指标的比较研究[J]. 中国农学通报,2014,30(7):224-227.
[18]曹慧明,史作民,周晓波,等. 植物对低温环境的响应及其抗寒性研究综述[J]. 中国农业气象,2010,31(2):310-314, 319.
[19]吴惠俐,李雪平,高健,等. 禾本科植物抗寒机理的研究进展[J]. 竹子研究汇刊,2014,33(1):7-11.
[20]宋正熊,朱列书,尹佳,等. 低温胁迫对烟草幼苗生化指标的影响及相关性分析[J]. 江西农业学报,2014,26(2):99-101, 105.
