不同种源麻竹表型多样性及其与环境因子的相关性
2023-09-27李东宝何天友荣俊冬郑郁善陈礼光
李东宝, 吴 敏, 余 蓉, 何天友, 荣俊冬, 郑郁善,, 陈礼光
〔福建农林大学: a. 林学院(碳中和学院), b. 风景园林与艺术学院, 福建 福州 350002〕
植物表型多样性普遍认为是遗传多样性和环境多样性相互作用的结果[1-3],其中遗传多样性是一个物种所涵盖的群体间和群体内个体间的遗传变异的总和,包括表型、细胞学、生化和分子等方面的遗传变异[4-5]。植物表型是遗传多样性的直观体现,利用表型间的差异对遗传多样性进行研究能够直观地揭示种群的遗传结构和变异程度,是进行遗传育种的基础工作。目前,已有学者对箭竹(FargesiaspathaceaFranch.)[6]、绿竹(BambusaoldhamiiMunro)[7]、圣音竹〔Phyllostachysedulisf.tubaeformis(S. Y. Wang) Ohrnb.〕[8]、毛竹〔Phyllostachysedulis(Carriere) J. Houzeau〕[9]、糙花少穗竹〔Oligostachyumscabriflorum(McClure) Z. P. Wang et G. H. Ye〕[10]和粉单竹(BambusachungiiMcClure)[11]等竹种进行了表型研究,为相关竹种的资源保护和遗传改良奠定了基础。
麻竹(DendrocalamuslatiflorusMunro)为禾本科(Poaceae)牡竹属(DendrocalamusBenth.)一种大型合轴丛生竹,为中国南方栽种范围最广的牡竹属竹种[12]。当前,对于麻竹的研究多数集中在笋芽萌发[13]、竹笋离体繁殖[14]、环境胁迫响应机制[15]和竹笋增产[16]等方面,而有关麻竹表型多样性的研究虽有初步报道[17]21-48,但调查的种源地较少,对种源的分布及其遗传丰富度、种源间和种源内的变异、种源的地理变异格局等均不明确,不利于麻竹遗传育种工作的进一步开展。鉴于此,本研究对20个麻竹种源进行调查、收集,系统测定了麻竹17个表型性状,研究不同种源麻竹表型性状的地理变异规律及其与环境因子的相关性,探索麻竹生长对异质性环境的响应模式以及麻竹的地理变异水平和规律,以期为麻竹遗传改良提供理论基础和数据支持。
1 材料和方法
1.1 材料
于2023年3月至4月在麻竹分布区选取具有代表性的种源地[18]7-9,包括福建省(永泰县、漳平市、福安县、永安市和华安县)、四川省(东坡县和富顺县)、重庆市(荣昌县和涪陵区)、云南省(陇川县和绥江县)、广西壮族自治区(兴宁区、永福县和西乡塘区)、广东省(揭东县和仁化县)、贵州省(兴义县和望谟县)、浙江省(苍南县)和江西省(章贡区),共20个种源地,种源地基本情况见表1,经度、纬度和海拔通过手持GPS仪测量,降水量和温度等气象数据来自中国气象数据中心(https:∥www.date.cma.cn/),为当地近30年的平均值。
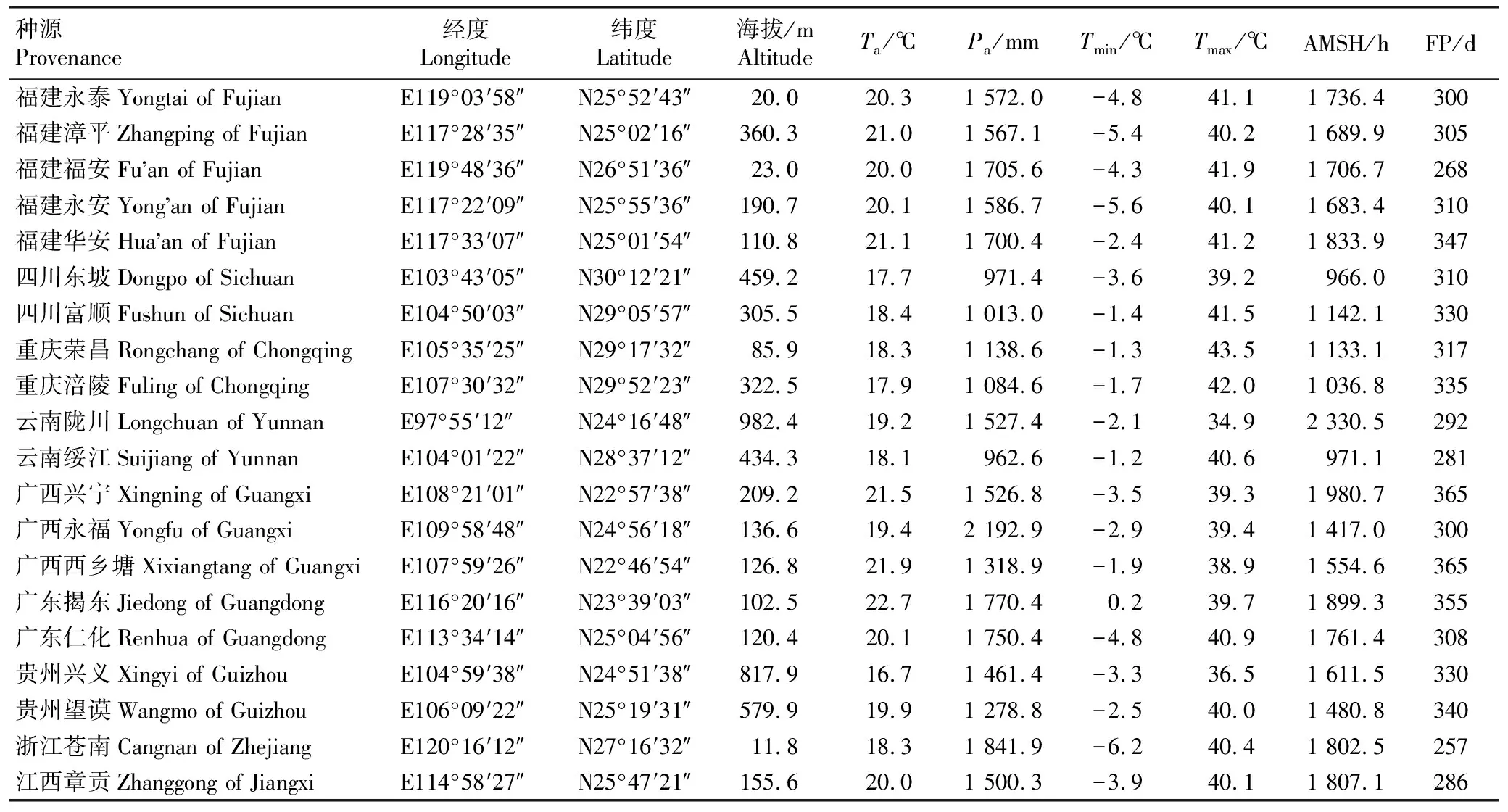
表1 不同麻竹种源地的基本情况1)
由于选取麻竹天然更新林较为困难,在调查前咨询当地相关林业部门和农户,调查的麻竹基本上引种超过20年。每个种源设置1个面积20 m×20 m的样地,参考邱尔发[18]7-8和熊文愈等[19]的方法判别株龄,对样地内株龄1、2和3 a麻竹进行每木检尺,使用围径尺(精确0.1 cm)测量胸径,根据各龄级麻竹平均胸径确定各龄级的样株。每个种源每个龄级5个重复,即取株龄1、2和3 a麻竹各5株,自基部伐倒,调查麻竹的表型性状。
1.2 表型性状的选取和测定
参考张闻博等[9]和邢新婷[17]21-24的方法选取样株的17个测量指标。总节数、枝下节数(第1个分枝以下节数)、分枝数和丛内株数(样竹所在丛内含有的株数)直接观测计数;使用卷尺(精度0.1 cm)分别测量株龄1、2和3 a麻竹的株高、最长节间长、枝下高(第1个分枝以下高度)、最长枝长、冠幅(南北方向的最大宽度)、胸高处节间长、叶长、叶宽,并计算叶长宽比;使用围径尺测量胸径;使用Peacock G型厚度计(日本Peacock公司,精度0.1 mm)测量叶厚(叶片的上、中、下部厚度的均值);使用Epson V700扫描仪(日本Epson公司)扫描叶片,并使用Image J软件计算叶面积;使用卡西洛双臂量角器(河南省邦特工量具有限公司,精度0.1°)测量分枝角度(具有明显侧枝的5个分枝角度的均值)。其中,叶片的选取为随机分别摘取株龄1、2和3 a样株上、中、下部的30枚成熟叶片,结果取平均值。
1.3 数据处理和分析
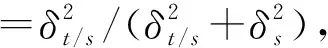
2 结果和分析
2.1 不同种源麻竹表型性状的比较和变异分析
2.1.1 不同种源麻竹表型性状的比较 不同种源麻竹17个表型性状的比较见表2。结果显示:四川富顺种源的株高、胸径和分枝角度最大,分别为19.39 m、11.74 cm和87.23°,总体显著高于其他种源;贵州兴义种源的株高和胸径最小,云南陇川种源的分枝角度最小,分别为6.74 m、4.61 cm和30.73°。云南绥江种源的最长节间长、总节数和胸高处节间长最大,分别为60.33 cm、39.0和41.40 cm,总体显著高于其他种源;重庆涪陵种源的最长节间长和总节数最小,福建永泰种源的胸高处节间长最小,分别为35.91 cm、13.0和29.90 cm。贵州望谟种源的枝下节数、枝下高和丛内株数最大,分别为22.0、7.63 m和34.0,显著高于其他种源;四川东坡、广西永福和浙江苍南种源的枝下节数均最小(3.0),浙江苍南种源的枝下高最小(0.63 m),重庆荣昌种源的丛内株数最小(2.0)。浙江苍南种源的分枝数最大(29.0),总体显著高于其他种源;贵州望谟种源的分枝数最小(4.0)。广东仁化种源的最长枝长和冠幅最大,分别为6.85 m和6.57 m,总体显著高于其他种源;重庆涪陵种源的最长枝长和四川东坡种源的冠幅最小,分别为0.78 m和0.46 m。广西兴宁种源的叶长、叶宽和叶面积最大,分别为43.60 cm、8.58 cm和240.38 cm2,总体显著高于其他种源;福建漳平种源的叶长、四川东坡种源的叶宽和广东仁化种源的叶面积最小,分别为24.46 cm、4.33 cm和80.64 cm2。浙江苍南种源的叶厚最大(0.26 mm),总体显著高于其他种源;福建永泰种源的叶厚最小(0.15 mm)。四川东坡种源的叶长宽比最大(7.36),显著高于其他种源;福建漳平种源的叶长宽比最小(4.23)。
表2 不同种源麻竹17个表型性状的比较
方差分析结果(表3)显示:不同种源麻竹的叶厚在种源间存在极显著差异,其他16个表型性状在种源内和种源间均存在极显著差异。说明麻竹表型存在较为丰富的变异。
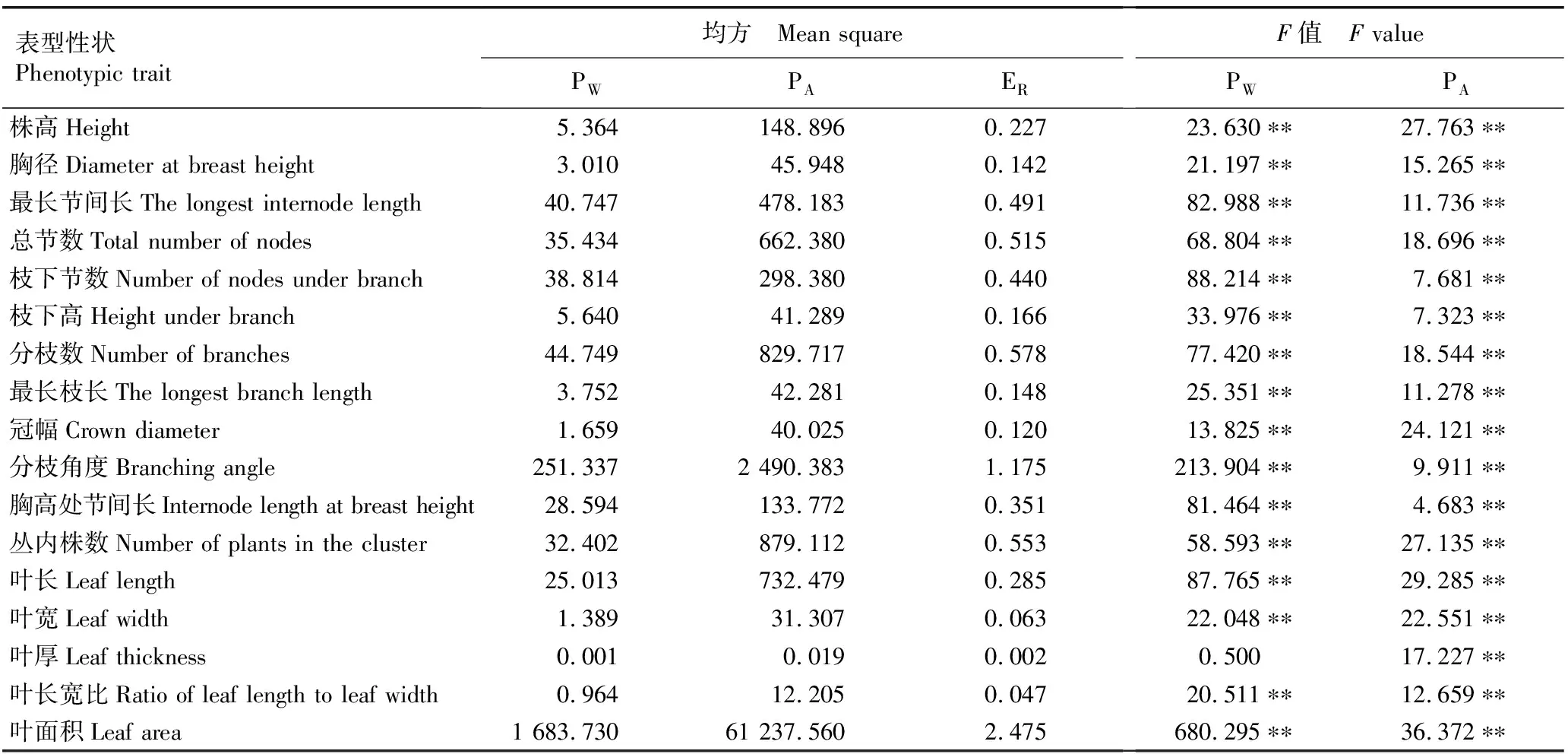
表3 不同种源麻竹17个表型性状的方差分析1)
2.1.2 不同种源麻竹表型性状的变异分析 不同种源麻竹17个表型性状的变异系数见表4。结果显示:麻竹各表型性状的变异系数均值在13.64%~67.91%之间,其中,枝下高的变异系数均值最大,枝下节数(59.36%)次之,最长节间长的变异系数均值最小。从不同种源地来看,不同种源麻竹表型性状的变异系数均值在14.21%~43.93%之间,其中,云南陇川种源的变异系数均值最大,明显高于其他种源,四川东坡种源(43.85%)次之,广东仁化种源的变异系数均值最小。总体上看,20个种源麻竹17个表型性状的变异系数均值为30.84%,其中,云贵川渝地区种源表型性状的变异系数均值较大,东南沿海地区种源居中,广东和广西地区种源表型性状的变异系数均值较小。
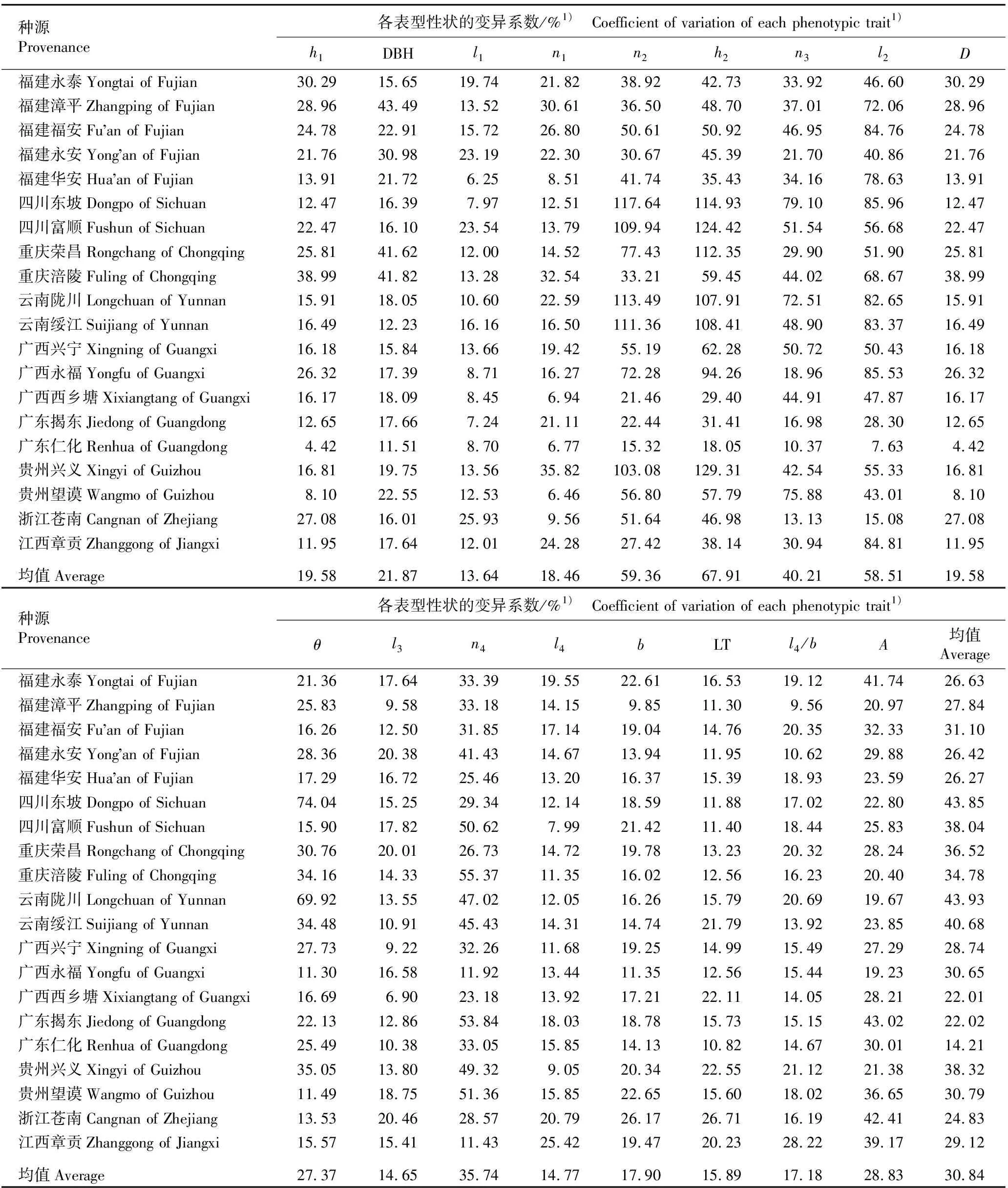
表4 不同种源麻竹17个表型性状的变异系数
2.2 不同种源麻竹表型性状的分化分析
不同种源麻竹17个表型性状的方差分量和表型分化系数见表5。结果显示:麻竹17个表型性状种源间的表型分化系数在6.10%~77.47%之间,均值为41.61%,其中,株高、丛内株数和冠幅的表型分化系数较高,分别为77.47%、76.63%和71.98%;种源内17个表型性状的方差分量百分比在7.14%~74.11%之间,均值为44.48%,而种源间17个表型性状的方差分量百分比在3.57%~59.43%之间,均值为37.25%,说明麻竹表型性状在种源间和种源内均存在不同程度的变异,且种源内变异大于种源间变异。

表5 不同种源麻竹17个表型性状的方差分量和表型分化系数
2.3 不同种源麻竹表型性状的聚类分析
基于麻竹17个表型性状,对20个种源进行聚类分析(图1)。结果显示:在欧氏距离6.8处,20个种源可分为3类:第1类包括福建永泰、四川东坡、重庆荣昌、重庆涪陵、四川富顺、云南陇川、云南绥江、广西永福和贵州兴义的种源;第2类包括福建漳平、江西章贡、广东仁化、浙江苍南、福建福安、福建华安、福建永安、广西兴宁和贵州望谟的种源;第3类包括广东揭东和广西西乡塘的种源。总体上看,20个麻竹种源并没有严格依据地理位置聚类,可分为云贵川渝地区一类、东南沿海地区一类和粤桂地区一类。说明麻竹表型变异存在片段性和不连续性,与麻竹表型性状的区域化变异情况基本一致。
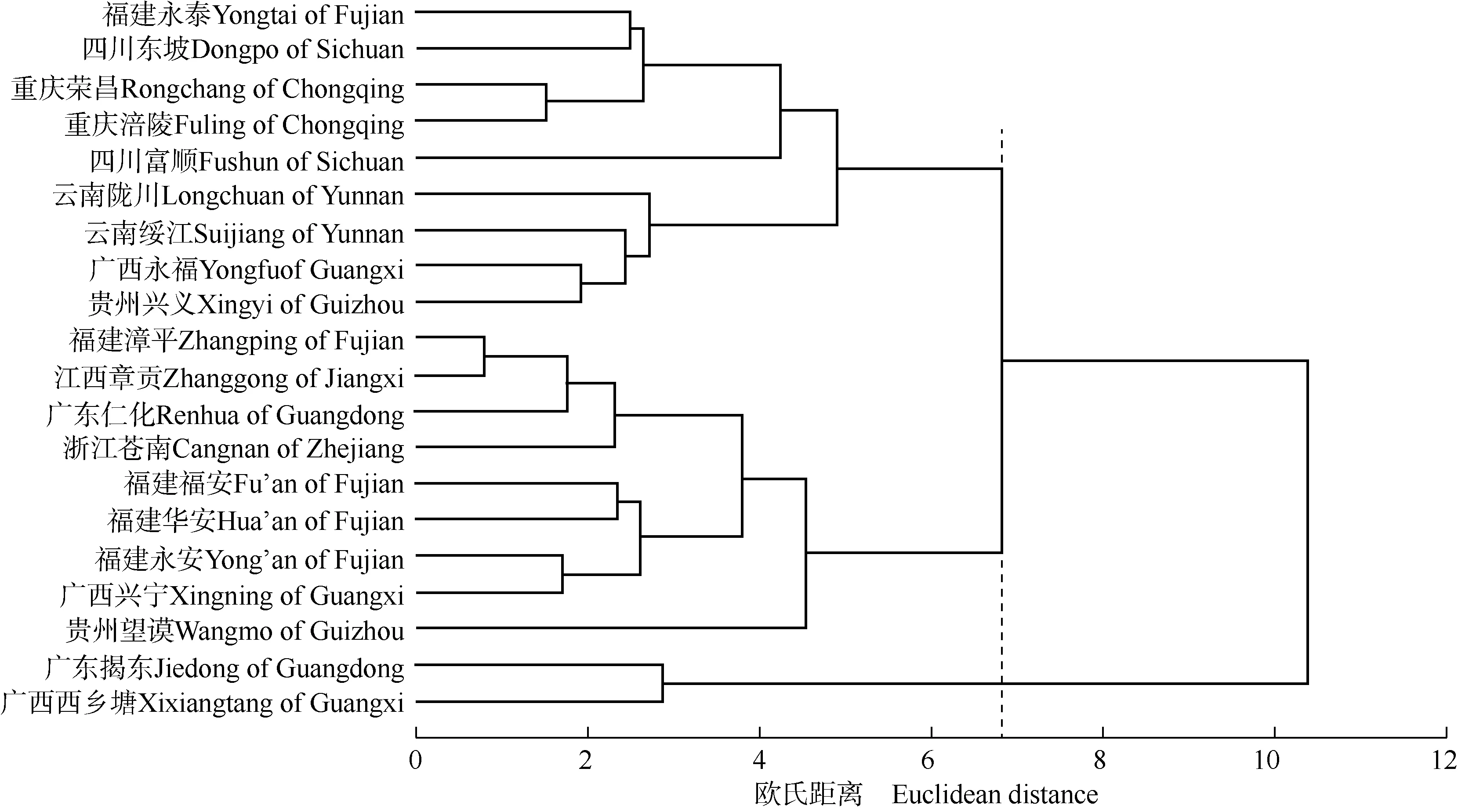
图1 基于17个表型性状的20个麻竹种源的聚类图
2.4 麻竹表型性状与环境因子的相关性分析
将不同种源麻竹17个表型性状与环境因子进行相关性分析,结果见表6。结果显示:麻竹大多数表型性状与除海拔外的环境因子间呈显著或极显著相关;仅枝下节数、枝下高、最长枝长和冠幅受海拔影响较大,与海拔呈显著或极显著相关。株高与经度、纬度和年均降水量呈极显著负相关,与年均温、极端最低温和极端最高温呈极显著正相关;胸径、最长节间长和总节数与经度、纬度、极端最低温和无霜期呈显著或极显著负相关,与年均温和极端最高温呈极显著正相关;枝下节数和枝下高与海拔和极端最高温呈显著或极显著正相关;分枝数与经度、年均降水量和年均日照时数呈极显著正相关,与年均温和无霜期呈极显著负相关;最长枝长和冠幅与除极端最低温和无霜期外的所有环境因子呈极显著相关;胸高处节间长与年均温、年均降水量和无霜期呈极显著负相关,与年均日照时数呈极显著正相关;丛内株数、叶长、叶宽和叶面积与经度和纬度呈显著或极显著负相关,叶长、叶宽、叶厚和叶面积与年均温、年均降水量、极端最低温、极端最高温、年均日照时数和无霜期总体呈显著或极显著相关。总体上看,麻竹表型性状受到多个环境因子的综合影响,其中,经纬度、温度和降水量对麻竹主要生长指标有明显的影响,且相较于极端最低温,极端最高温对麻竹生长的影响更大。
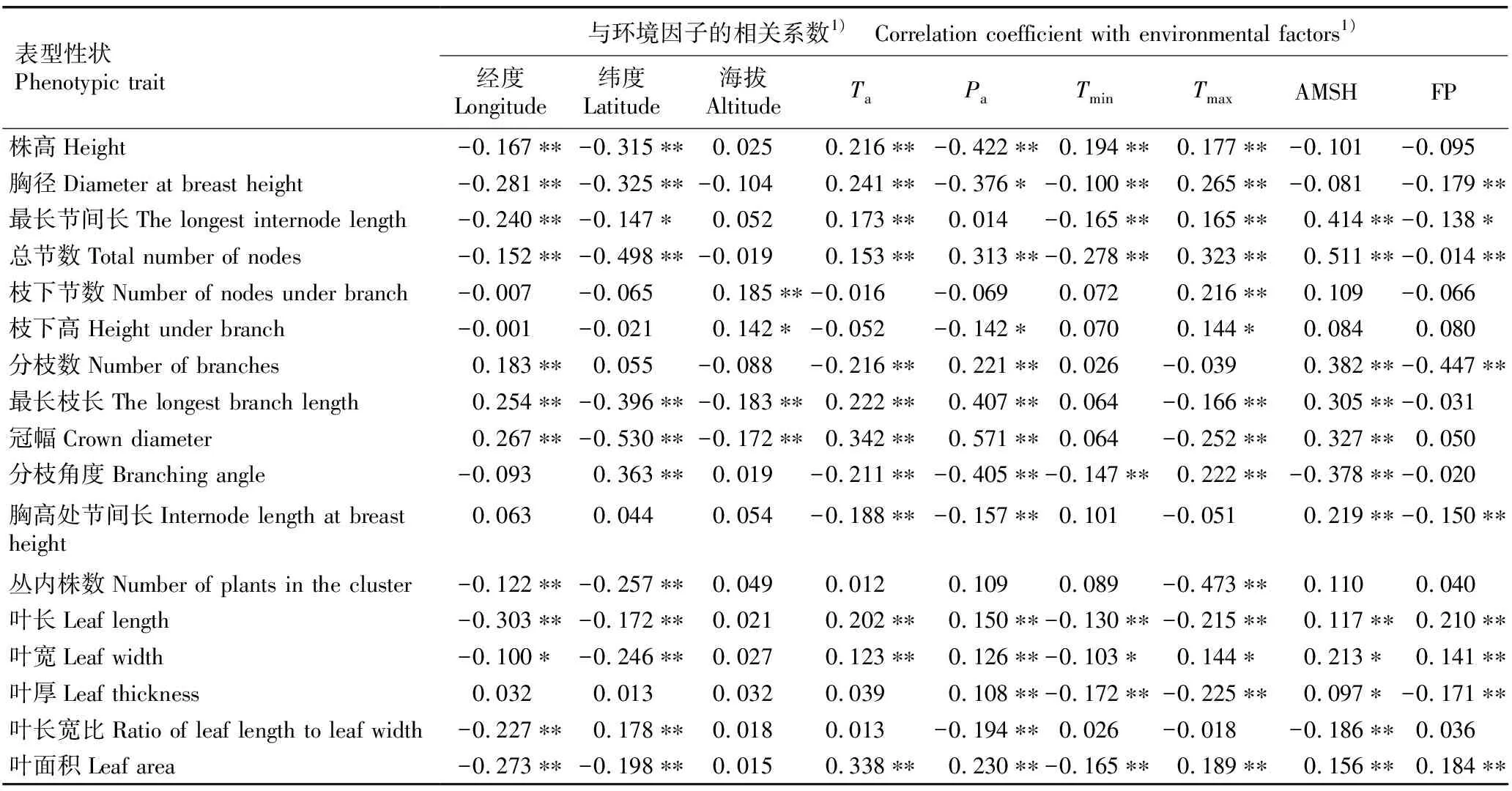
表6 不同种源麻竹17个表型性状与环境因子的相关性分析
3 讨 论
受自身遗传和环境的综合影响,植物在表型形态特征及生理特性等方面会产生一系列变化[23]。表型变异广泛存在于不同植物之间,如油松(PinustabuliformisCarriere)[24]、无患子(SapindussaponariaLinn.)[25]和蒙古栎(QuercusmongolicaFisch. ex Ledeb.)[26]。本研究中,麻竹表型性状在种源内和种源间总体上存在极显著差异,说明麻竹表型性状的变异较为丰富,这与邢新婷[17]25-29在麻竹的遗传育种中得到的结果相似。已有研究结果表明:株高和胸径是竹类经济性状,是反映竹类生物量及生长情况的重要指标[27]。本研究中,四川富顺种源的株高和胸径最大,可以作为材用竹的选育目标。从叶片角度看,叶的大小与叶的能量平衡有关,一般认为植物在资源丰富的生境中,往往具有较大的叶面积[28-29]。本研究中,广西兴宁的年均降水量较多、年均日照时数较长,该地的麻竹叶长、叶宽、叶面积均最大,说明广西兴宁的麻竹生长在资源较为丰富的环境中。此外,也有研究认为生长在水分匮乏生境下的植物叶片一般较厚[30],与本研究中的结果不同,浙江苍南属东南沿海地区,降水较多,但该种源地的麻竹叶较厚,这可能是为了适应风力较大的环境。
表型变异系数能直观反应植物表型变异的程度,变异系数越大,植物多样性越丰富,植物对环境变化的适应力越强[31]。本研究结果显示:20个种源麻竹17个表型性状总体的变异系数均值为30.84%,变异水平高于文冠果(XanthocerassorbifoliumBunge)[32]、珙桐(DavidiainvolucrataBaill.)[33]和槐〔Styphnolobiumjaponicum(Linn.) Schott〕[34],变异系数均值分别为11.72%、16.22%和9.40%,说明麻竹表型多样性丰富,反映出在长时间的地理隔离中,不同种源麻竹之间的基因交流可能受到了严重的阻碍[35]。本文中20个麻竹种源总体表现为云贵川渝地区的种源表型变异较大,东南沿海地区居中,粤桂地区的种源表型变异较小。产生这种变异规律的原因可能有以下几个方面:1)空间地理限制,由于麻竹天然林为零星分散,基本上都是栽培类型,麻竹自然分布区内少见成片的天然林分[17]22,其地理分布区基本上是当地相关林业部门或农户从附近麻竹分布区引种而来,这种引种方式使麻竹基因仅能在一定地理范围内进行交流,从而使得各种源间的变异幅度区域化;2)在遇到严峻的环境胁迫时,植物会发生更大程度的表型变异以适应外部环境的改变,由于云贵川渝地区海拔较高、东南沿海地区风力较大和雨水冲刷较强等原因,这些种源地的麻竹受到的生态环境压力较大,因而导致云贵川渝地区和东南沿海地区的种源表型变异较大;3)由于麻竹属于笋材两用竹,在麻竹自然分布区存在一些人为干预现象,在一定程度上影响了麻竹表型性状的分化方向。相关研究结果表明:环境的改变会造成植物株高和叶片大小等营养性状的改变,但繁殖器官在经历复杂的进化之后,变异程度较低,环境的差异是造成营养器官变异的重要原因之一[36]。本研究结果显示:枝下高、枝下节数和最长枝长等营养性状的变异系数总体在10%以上,而麻竹天然结实量较少,导致种子取样极为困难,因此对麻竹营养性状与繁殖器官的变异程度比较还有待进一步的研究和探索。
本研究选取的20个麻竹种源地基本上涵盖了全部的麻竹地理分布区,能较好地反映麻竹表型性状的变异特征。方差分析结果表明:麻竹种源内各表型性状的方差分量百分比均值(44.48%)大于种源间各表型性状的方差分量百分比均值(37.25%),表型分化系数均值为41.61%,说明麻竹表型性状差异的主要来源是种源内变异,但种源间仍有一定程度的表型分化,这反映出麻竹遗传与环境互作的复杂性,同时也是麻竹对不同环境适应的结果。产生这种结果的原因可能是:一方面,麻竹的繁殖方式多为无性繁殖,种源间基因交流困难,从而有利于种源间的遗传分化,且麻竹为丛生竹,同一丛内不同个体发育程度不同,差异明显,导致种源内的表型多样性高于种源间;另一方面麻竹种源的地理隔离和生境的片段化,进一步促进了麻竹种源间的表型分化[37]。与其他植物相比,麻竹的表型分化水平高于马尾松(PinusmassonianaLamb.)[22]、川西云杉(Picealikiangensisvar.rubescensRehder et E. H. Wilson)[38]和麻栎(QuercusacutissimaCarr.)[39],表型分化系数均值分别为6.64%、36.53%和23.98%;而低于云南黄连(CoptisteetaWall.)[40]、苦楝(MeliaazedarachLinn.)[41]和石斛(DendrobiumnobileLindl.)[42],表型分化系数均值分别为74.41%、54.47%和65.80%,表明麻竹表型分化位于中等水平。此外,本研究发现丛内株数的表型分化系数(76.63%)较大,这是因为丛内株数影响麻竹对养分吸收,当同一丛内有较多株时,可能会出现养分供应不足的情况,进而影响麻竹生长,而丛内株数的分化有利于麻竹对不同环境中养分的适应。野外调查发现,长势较好的麻竹大多同一丛内株数较少,这种丛内株数的高水平分化现象体现了麻竹对环境中养分变化的高度适应性。
不同植物对环境的适应性和敏感性存在差异,进而使植物表现出不同的地理变异格局[38]。聚类分析结果表明:20个麻竹种源的聚类并不完全受地理位置限制,这是因为麻竹多为栽培种,花粉和种子在种源间传播较为困难,即使地理位置较近的种源间也很少有基因交流的机会。这在青海野生中国沙棘(HippophaerhamnoidesLinn.)资源表型性状多样性研究[43]中也得到了类似的结果。此外,柳江群等[44]对长柄扁桃(PrunuspedunculataPall.)天然种群表型变异的研究发现,相似的生境是植物严格按照地理位置聚类的关键,而麻竹所处的环境和局部小气候相对独立,导致供试种源没有严格按照地理位置聚类。影响植物表型变异的因子有很多,其变异规律十分复杂,是遗传和环境共同作用的结果。例如:五角枫〔Acerpictumsubsp.mono(Maxim.) H. Ohashi〕表型性状呈现以经度和纬度并存的变异规律[45],浙江楠(PhoebechekiangensisC. B. Shang)表型性状呈现以海拔为主的变异规律[37],而山苍子〔Litseacubeba(Lour.) Pers.〕表型性状则呈现以年均降水量和海拔为主的变异规律[46]。本研究发现,麻竹的多数表型性状与经度、纬度、年均温、年均降水量、极端最高温和年均日照时数呈显著或极显著相关,这与麻竹生态习性以及其分布范围呈现东西较广、南北狭窄的特征有关[17]32。姬明飞等[24]的研究结果表明:树种分布的范围是影响树木与环境因子相关性的重要因子,一般来说,树种分布区越大、相关性也较大,分布区越小、相关性也较小。20个麻竹种源地分布范围的东西跨度较大,经度在东经97°55′12″~120°16′12″之间,年均温、年均降水量、年均日照时数和极端最高温等环境因子也各不相同,导致表型性状与环境因子相关性较高。同时,麻竹为阳性植物,喜高温、向阳的环境,而纬度越低的地区气温越高,这也是导致麻竹主要生长性状株高、胸径、最长节间长和总节数与极端最高温呈极显著正相关,与纬度呈显著或极显著负相关的主要原因。与海拔的相关性分析结果显示:仅枝下节数、枝下高、最长枝长和冠幅4个表型性状与海拔呈显著或极显著相关,其原因可能是相较于海拔,经度和纬度对麻竹表型性状的影响更大,也可能与没有在同一纬度上调查不同海拔的种源差异有关。另外,受调查因子的限制,本研究并没有考虑种源地的土壤状况、坡度和坡向等因子,因此,本研究仅为不同种源麻竹表型性状与环境因子相关性的初步报道,今后将开展进一步的深入研究。
4 结 论
不同种源麻竹表型性状存在丰富的变异性,除叶厚外,其余16个表型性状在不同种源内和种源间均存在极显著差异,变异系数均值在13.64%~67.91%之间,其中枝下高的变异系数均值最大,最长节间长的变异系数均值最小。麻竹种源内变异大于种源间,表型性状差异的主要来源是种源内变异;麻竹的表型分化位于中等水平,表型分化系数均值为41.61%。总体上看,麻竹种源并没有严格依据地理位置聚类,可分为云贵川渝地区一类、东南沿海地区一类和粤桂地区一类,其中,云贵川渝地区的种源表型变异较大,东南沿海地区居中,粤桂地区的种源表型变异较小。麻竹大多数表型性状与除海拔外的环境因子间呈显著或极显著相关,这种变异规律揭示了麻竹东西分布较广、南北分布狭窄的原因。
