海南清澜港海南海桑群落中物种生态位特征和种间联结性
2023-09-27张孟文钟才荣吕晓波方赞山
张孟文, 钟才荣, 吕晓波, 方赞山, 程 成
〔海南省林业科学研究院(海南省红树林研究院), 海南 海口 571100〕
目前,全球性的植被破坏和生物多样性损失严重,全世界约有39.4%的维管植物物种面临灭绝的威胁[1-2],中国受威胁的高等植物比例为15%~20%[3]。极端气候变化、人类活动干扰以及外来物种入侵等都是濒危植物数量大幅度减少的外在因子[4]。因此,濒危植物的保护已成为全世界关注的一个主要问题。濒危植物生态位特征为揭示其资源现状及发展趋势提供了重要依据,而种间联结关系反映了濒危植物所在群落的结构与功能特征[5-6]。通过开展濒危植物生态位特征和种间联结性研究,深入探讨濒危植物种内、种间关系及其在群落中的地位和作用,对揭示群落中物种共存机制和保护濒危植物具有十分重要的意义。
植物群落由多个相互影响的植物个体组成,通过对生境中的资源共享和竞争,表现出不同的种间关系和生态位特征[7]。目前,学者们常用生态位宽度和生态位重叠等指标描述群落内各物种的生态位特征,从而探讨物种对周边环境资源的竞争力以及分布特征[8]。此外,确定不同物种的空间关联程度对研究物种间的相互作用以及群落的组成和动态具有重要意义[9]。物种间的空间关联程度大体包含正联结、负联结和不联结3种情况及其显著性,一般采用总体联结指数、χ2检验、联结系数和Pearson相关系数等方法和指标来探讨群落物种间的联结性[10]。当前学者对生态位特征和种间联结性的研究对象主要为不同植物生活型、不同植被类型以及不同群落垂直结构的优势种或濒危种[11-16]。
红树林是自然分布在热带与亚热带海岸潮间带,并且物种组成以红树植物为主的植被类型[17]。作为蓝碳生态系统中惟一的木本植物,红树林在人类生命防线、物种基因库、蓝碳和海洋净化器等方面扮演重要角色[18]。但对红树林尤其是濒危红树植物的生态位特征和种间联结性等方面的研究相对不足,主要表现在2个方面:一方面是研究对象仅局限于少量的红树林群落优势种的生态位特征和种间联结性,另一方面是研究内容更侧重于红树林的群落结构和物种组成[19-21]。已有研究结果表明:中国红树植物种类约占全世界的1/3,但有近50%的红树种类处于不同程度的濒危状态,其中海桑属(SonneratiaLinn. f.)的濒危植物种数最多[22]。海桑属植物是典型的以种子繁殖的非胎生植物,树体高大,对潮间带周期性海水淹没环境有较强的适应性,既是红树林群落的重要组成部分,也是红树林保护与恢复的重要树种资源[23-24]。
海南海桑(Sonneratia×hainanensisW. C. Ko, E. Y. Chen et W. Y. Chen)为千屈菜科(Lythraceae)海桑属常绿乔木,生长在立地条件较好、土壤松软、淤泥深厚,且常有潮水浸淹的中潮滩和高潮滩上[25]。海南海桑为中国特有种,仅天然分布于海南清澜红树林省级自然保护区[26]。受人类活动干扰,海南海桑的生存状态非常严峻,个体数量极其稀少,种群处于极度濒危状态,已被国家林业和草原局列为中国极小种群野生植物[27]。近年来,学者对海南海桑的研究主要集中于资源现状[25]、繁殖特征[28]以及适应性[29]25等方面,而关于种间关系对海南海桑种群分布的影响目前还不清晰,研究海南海桑的生态位特征和种间联结性是保护与恢复该种群的重要途径。因此,本文以海南清澜港自然分布的海南海桑群落为研究对象,对其中物种生态位特征和种间联结性进行定量研究,比较不同物种间的相互关系,以期为海桑属濒危植物的保护和恢复提出科学的策略。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于海南清澜红树林省级自然保护区的八门湾片区(东经110°48′35″~110°53′08″、北纬19°37′11″~19°37′58″)。保护区属于热带海洋性季风气候,年均温24.3 ℃,极端最高温39.1 ℃,极端最低温4.7 ℃,年均降水量1 975.2 mm,年均日照时数1 922.5 h,年均蒸发量1 892.4 mm;该区域的潮汐类型为不规则的全日潮,平均潮差0.75 m,最大潮差2.06 m,海水盐度1.8%~2.8%。目前保护区总面积2 914.6 hm2,其中核心区面积884.1 hm2、缓冲区面积966.6 hm2、实验区面积1 063.9 hm2,保护了面积约为1 444.5 hm2的红树林植被[29]13。
1.2 研究方法
1.2.1 样地设置和样方调查 于2021年3月至5月对海南海桑的资源现状进行系统调查,最终确定了该物种的地理分布和数量特征。在此基础上,采用典型取样法[30],在海南海桑集中分布区域设置16个面积10 m×10 m的样地,调查面积共1 600 m2。调查时将每个样地划分为4个面积5 m×5 m的样方,采用胸径尺(精度0.1 cm)和塔尺(精度0.1 cm)测量并记录各样方内所有胸径大于等于1 cm的植株的胸径、树高和枝下高。
1.2.2 重要值计算 参考文献[31]中的方法计算海南海桑群落中各物种重要值。
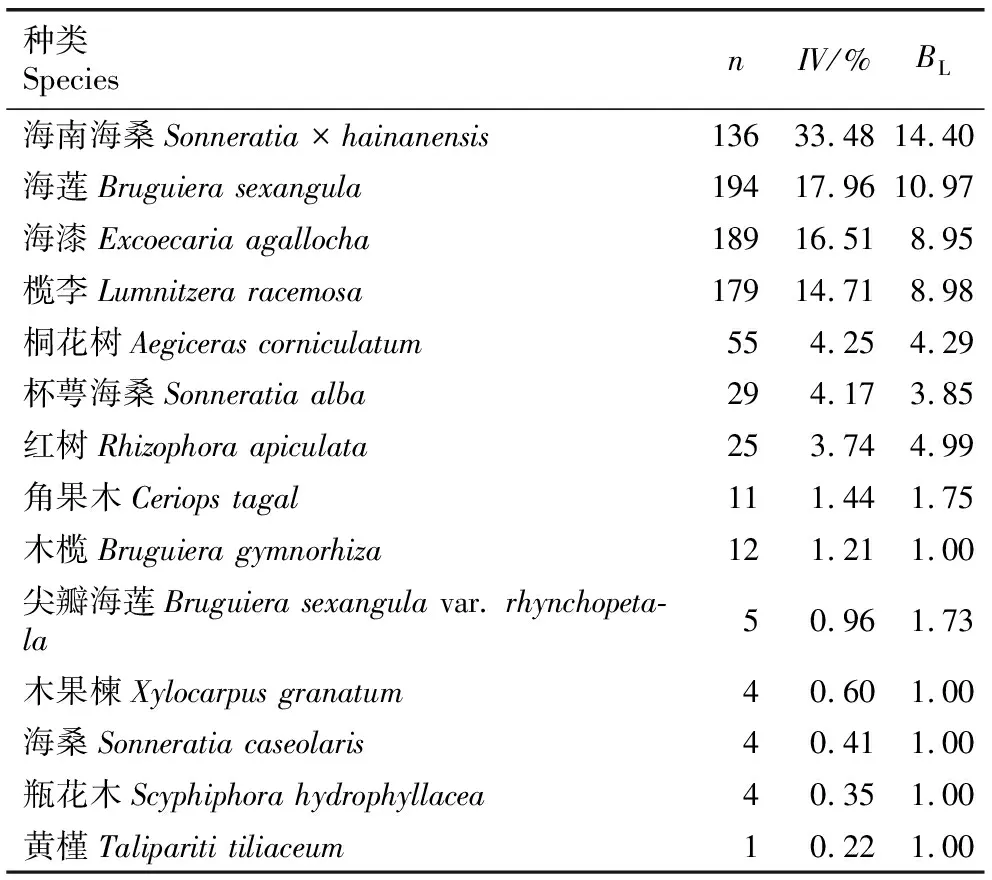
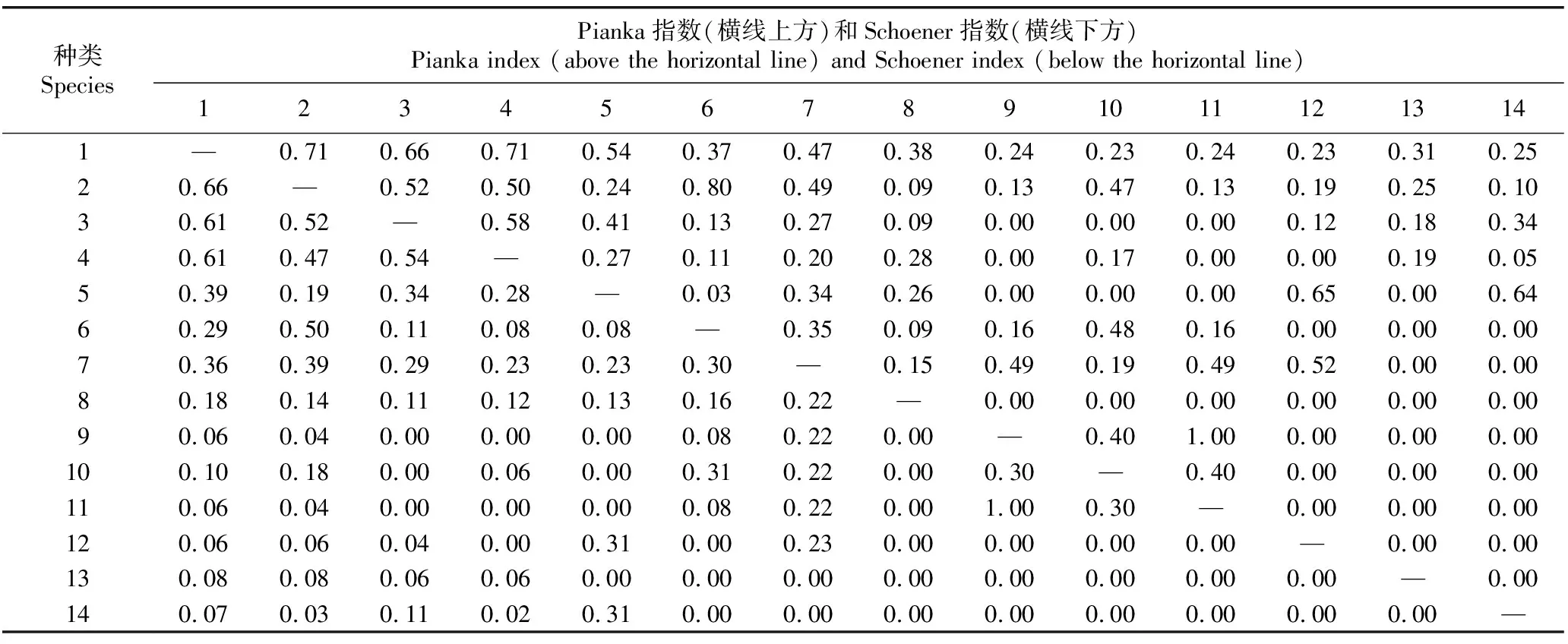
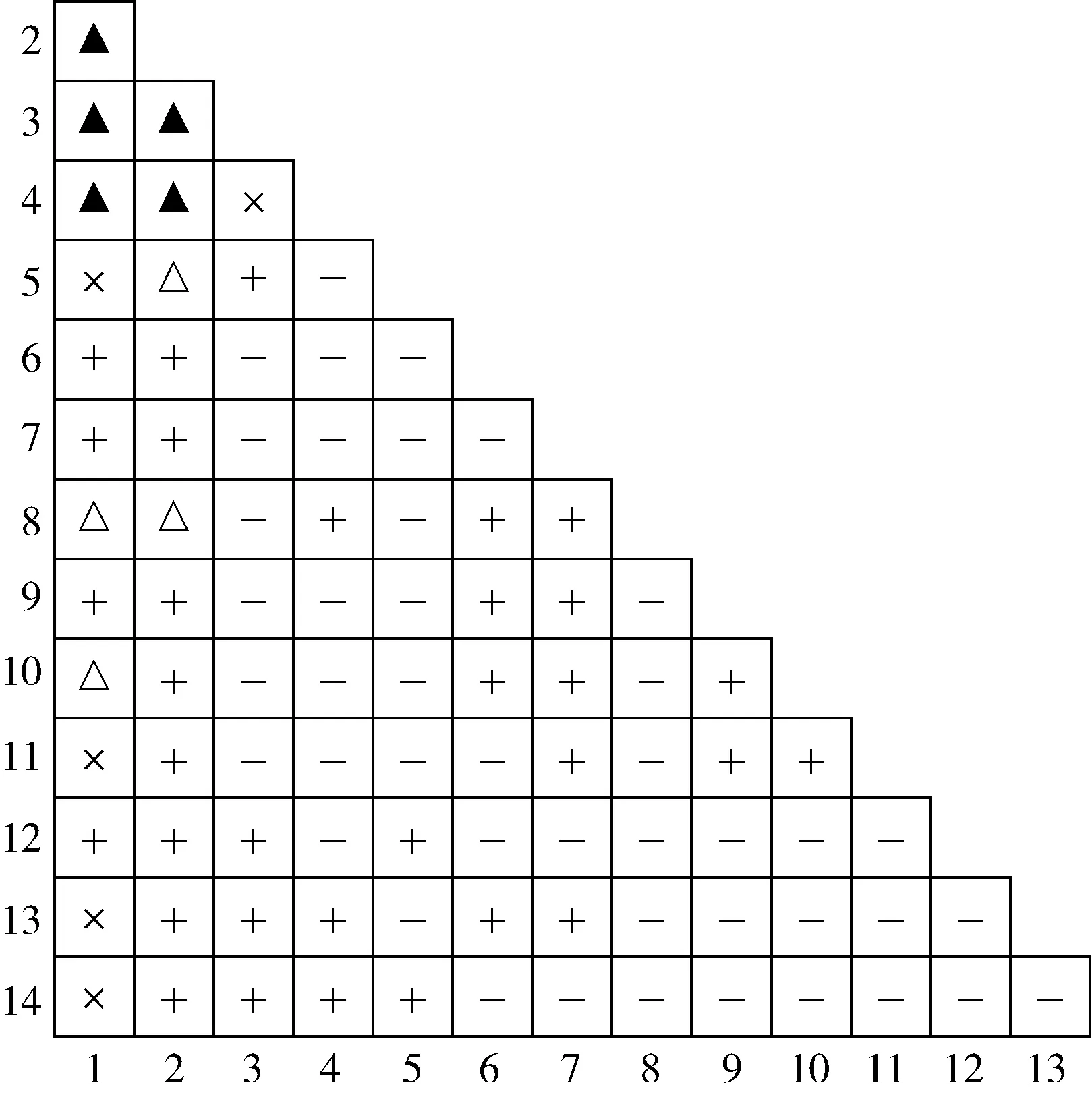
1.2.3 生态位分析 参考文献[32-33]中的方法进行生态位分析。采用Levins指数(BL)表示海南海桑群落中各物种的生态位宽度。采用Pianka指数(Qik)表示各物种间的生态位重叠:Qik=0.00表示生态位完全不重叠;0.00 采用EXCEL 2007软件整理和分析数据,采用Origin 2021软件绘图。 海南清澜港海南海桑群落物种组成及Levins指数见表1。结果显示:海南海桑群落共有14种(变种,下同)植物,隶属于8科10属,其中,海南海桑有136株,个体数排名第4,林下未见幼龄个体。海南海桑的重要值最大(33.48%),其后依次为海莲〔Bruguierasexangula(Lour.) Poir.〕、海漆(ExcoecariaagallochaLinn.)、榄李(LumnitzeraracemosaWilld.),重要值均大于10%;尖瓣海莲(Bruguierasexangulavar.rhynchopetalaKo)、木果楝(XylocarpusgranatumKoenig)、海桑〔Sonneratiacaseolaris(Linn.) Engl.〕、瓶花木(ScyphiphorahydrophyllaceaGaertn. f.)和黄槿〔Taliparititiliaceum(Linn.) Fryxell.〕的重要值均小于1%,其中黄槿的重要值最小(0.22%)。 表1 海南清澜港海南海桑群落物种组成及Levins指数1) 结果(表1)还显示:群落中物种间的生态位宽度存在差异。海南海桑、海莲、海漆和榄李的Levins指数较大,分别为14.40、10.97、8.95和8.98,表明这4种植物的生态位宽度较大,对生境的适应性较强。桐花树〔Aegicerascorniculatum(Linn.) Blanco〕、杯萼海桑(SonneratiaalbaJ. Smith)和红树(RhizophoraapiculataBl.)的Levins指数分别为4.29、3.85和4.99,表明这3种植物的生态位宽度处于中等水平,对生境的适应性一般。角果木〔Ceriopstagal(Perr.) C. B. Rob.〕和尖瓣海莲的Levins指数较小,分别为1.75和1.73;木榄〔Bruguieragymnorhiza(Linn.) Savigny〕、木果楝、海桑、瓶花木和黄槿的Levins指数最小,均为1.00,表明这7种植物的生态位宽度较小,对生境的适应性较弱。 海南清澜港海南海桑群落物种间的Pianka指数(Qik)和Schoener指数(Cik)见表2。结果显示:海南海桑群落中14种植物共组成91个种对,其中,Qik=0.00的种对有33个,占总种对数的36.3%;0.00 表2 海南清澜港海南海桑群落物种间的Pianka指数和Schoener指数1) 结果(表2)还显示:91个种对中,Cik=0.00的种对有33个,占总种对数的36.3%。0.00 2.3.1 总体联结性分析 海南清澜港海南海桑群落物种间的方差比率(VR)为0.59,小于1,说明海南海桑群落物种间总体表现为负联结。进一步采用统计量W检测VR值偏离1的显著性,W值为9.44,在(7.96,26.30)区间内,说明海南海桑群落物种间总体联结性不显著。 2.3.2 种间联结性分析 海南清澜港海南海桑群落物种间的χ2检验半矩阵图见图1。结果显示:海南海桑群落物种组成的91个种对中,正联结种对有38个,负联结种对有48个,无关联种对有5个,分别占总种对数的41.8%、52.7%和5.5%,正负关联比为0.79。在38个正联结种对中,显著正联结种对有5个,分别为海南海桑和海莲、海南海桑和海漆、海南海桑和榄李、海莲和海漆以及海莲和榄李。在48个负联结种对中,显著负联结种对有4个,分别为海南海桑和角果木、海南海桑和尖瓣海莲、海莲和桐花树以及海莲和角果木。海南海桑与其他物种组成的13个种对中,显著正联结种对有3个,不显著正联结种对有4个,显著负联结种对有2个,无关联种对有4个。 ▲: 显著正联结Significant positive association; △: 显著负联结Significant negative association; +: 不显著正联结Non-significant positive association; -: 不显著负联结Non-significant negative association; ×: 无关联No association. 植物的重要值与生态位宽度均可用于评估某个种群在特定群落中的地位和作用,但二者间存在差异[35]。重要值主要侧重植物在群落中的主体地位,而生态位宽度体现在植物与环境的相互关系及植物对周边资源的利用能力上[36]。已有研究结果表明:生态位宽度与植物濒危没有必然的联系[37]。有的濒危植物在群落中生态位宽度较小,与群落中其他物种竞争激烈[30];而有的濒危植物在群落中生态位宽度较大,适应性强[38-39]。本研究中,海南海桑群落中共有14种植物,其中海南海桑的重要值(33.48%)和生态位宽度(Levins指数为14.40)在群落中均排名第1。表明在该群落中海南海桑的分布范围最广,对环境的适应性和资源的利用能力均较强。群落中其他物种的重要值和生态位宽度明显低于海南海桑,表明这些种类在群落中对环境的适应性和资源的利用能力均较弱。其他濒危植物如细果秤锤树(SinojackiamicrocarpaC. T. Chen et G. Y. Li)在群落中生态位宽度也大于其他物种[40]。出现这种现象的原因可能与样地设置有关,此次调查样地位于海南岛惟一仅存的海南海桑集中分布区域,所有海南海桑个体均为高大乔木,在群落垂直结构中处于最顶层,因此海南海桑能够展现出较强的种间竞争能力。 一般认为,生态位宽度越大的物种对环境的适应能力越强,往往也伴随着较高的生态位重叠度[8]。相反,生态位宽度较小的物种,利用资源的能力较弱,生态位重叠程度较低[15]。本研究中,海南海桑、海莲、海漆和榄李的生态位宽度均较大,分布范围广,这些物种组成的种对的生态位重叠程度较高。但这并不是完全绝对的,本研究还发现个别生态位宽度较小的物种间会出现生态位重叠程度较高的现象,如桐花树、海桑、红树和黄槿的生态位宽度较小,但这些物种组成的种对(桐花树和海桑、红树和海桑以及桐花树和黄槿)的生态位重叠程度却较高。这在郄亚栋等[41]的研究中也有相似的结果。这可能是因为群落中物种间同时存在种间竞争和相互促进的关系[31]。此外,本研究中,仅有13.2%种对的生态位重叠程度较高(Pianka指数大于等于0.50),并且所有种对的Pianka指数的均值为0.21,表明海南海桑群落中大部分种对间的生态位重叠程度较低,种间竞争程度较弱。 已有研究结果表明:在湿地环境中,物种间多存在共享资源不足的情况,物种间的生态位重叠除了表征物种生态学特性相似性外,还反映物种间存在竞争关系,且与环境条件的变化相关[42]。本研究中,海南海桑群落中大部分物种间的生态位相似性较低,表明这些物种对环境资源的竞争不激烈。究其原因,一方面可能与各物种对异质性生境的适应性差异有关,另一方面可能是人类的养殖活动导致原有生境破碎化,产生了一系列不适宜物种生长的环境因子(光照、盐度和潮汐),难以满足更多物种生长所需的资源[43]。说明海南海桑群落中物种生态位特征不仅与各物种的生物学特性有关,也与所处环境条件密不可分。 综合以上分析可知,生态位特征可能不是海南海桑濒危的主要因子,导致其种群逐渐衰退且呈斑块状分布的主要原因是人为干扰和自身繁殖受限[25]。 物种间的总体联结性与其所处群落的稳定性紧密相连,反映了该植物群落的演替趋势[11]。研究结果[44]表明:在初级演替阶段,群落物种间的关联程度通常较低,并且常常会产生较大的负联结,竞争相对比较激烈;随着演替的进行,群落物种间的正联结程度逐渐加强;当群落达到顶极阶段时,群落总体上呈现显著的正联结特性。本研究中,海南海桑群落物种间总体表现为不显著负联结,表明该群落还处于不成熟的演替阶段。相关研究结果也表明:海南海桑天然林和人工林林下均未发现幼树或幼苗,各种群的更新状况极差[25,45]。说明海南海桑种群更新不良。原因可能与海南海桑群落的生境片段化有关。受人类活动影响,原先连片分布的海南海桑群落,现在被切割分布于相邻的养殖塘中。由于养殖塘资源有限,加上物种对生境资源存在共同需求,群落中部分物种间竞争激烈,因此,群落整体处于相对不稳定的状态。在桫椤〔Alsophilaspinulosa(Wall. ex Hook.) R. M. Tryon〕和台湾杉(TaiwaniacryptomerioidesHayata)等濒危植物研究中也得到了相似的结果[10,46]。 海南海桑群落14种植物组成的91个种对中有84.6%的种对联结性不显著,说明群落物种间联结性不强,具有一定的独立性。在海南海桑与其他植物组成的13个种对中,显著正联结种对有3个,显著负联结种对有2个,显著联结种对数仅占38.5%,说明海南海桑与群落中大部分物种联结性较弱,促进和竞争均不激烈。原因可能是海南海桑处于群落垂直结构的第1层,占据独特的生态位,对生境资源的利用具有绝对的优势。陈玉凯等[37]认为,种间正联结性越强、生态位重叠值越大,种间负联结性越强、生态位重叠值越小。钟彦龙等[47]的研究结果显示:植物种对的正联结体现了植物利用资源的相似性和生态位的重叠性,负联结体现了物种间的排斥性,是长期适应不同微环境的结果,也是生态位分离的反映。本研究也得到了类似的结果,与海南海桑显著正联结的3个种对,海南海桑和海莲、海南海桑和海漆以及海南海桑和榄李的Pianka指数分别为0.71、0.66和0.71,生态位重叠程度较高,这可能是由于海南海桑为群落优势种,处在最高层,为处于其下层的海莲、海漆和榄李提供了一个良好的生存环境;而与海南海桑显著负联结的2个种对,海南海桑和角果木、海南海桑和尖瓣海莲的Pianka指数分别为0.38和0.23,生态位重叠程度较低,这可能与群落中不同物种的生态适应性差异有关[32],海南海桑主要分布在潮间带的高潮位,角果木分布在中潮位,由于二者对海水潮汐的适应性不同,形成了生态位的分离。另有研究结果表明:群落结构越稳定,群落中无联结种对比例越高[44]。而海南海桑群落由于植被受到破坏和生境退化[43],群落中无联结种对数较少,仅占总种对数的5.5%,海南海桑群落演替为稳定的顶极群落是一个很漫长的过程。因此,应加强对海南海桑的保护,且在保护工作中不仅要保护仅存的成年植株,还要保护其所处的群落与生境。 海南清澜港海南海桑群落各物种间存在一定程度的生态位重叠和生态位相似性,但重叠和相似性程度均不高;群落物种间总体表现为不显著负联结,海南海桑与大多数物种间的联结性均不强,具有一定的独立性。此外,海南海桑在群落中生态位宽度最大,对其群落生境适应性较强,生存状况良好。说明生态位特征和种间联结性可能不是海南海桑濒危的主要因子,导致其种群逐渐衰退且呈斑块状分布的主要原因是人为干扰和自身繁殖受限。因此,可从开展人工辅助育种工作、加大就地保护力度和优化迁地保护策略方面考虑海南海桑种群的保护与恢复。同时,在就地保护过程中,需关注与海南海桑呈负联结的物种,特别是显著负联结的角果木和尖瓣海莲,并对其适度疏伐。这些措施对于进一步保护与恢复海桑属濒危红树植物具有重要意义。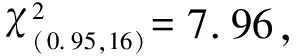
1.3 数据统计和分析
2 结果和分析
2.1 物种组成及生态位宽度分析
2.2 生态位重叠和生态位相似性分析
2.3 联结性分析
3 讨 论
3.1 海南海桑在群落中的生态位特征
3.2 海南海桑群落的种间联结性
4 结 论
