长江水系中华绒螯蟹背甲几何形态的产地差异
2023-09-27薛竣仁刘洪波陈修报
唐 静,薛竣仁,刘洪波,姜 涛,陈修报,杨 健,
( 1.南京农业大学 无锡渔业学院,江苏 无锡 214081;2. 中国水产科学研究院 淡水渔业研究中心 渔业微化学研究室,江苏 无锡 214081 )
中华绒螯蟹(Eriocheirsinensis)俗称大闸蟹、河蟹,属节肢动物门甲壳总纲十足目绒螯蟹属[1],其具有生长快、肉质鲜嫩、营养及经济价值高等特点[2-3]。由于其水产品资源生物的分布地区及生长环境不同,其在外部形态等品质方面可出现地理性的差异[2,4-5]。这种差异在不同产地的大闸蟹上亦有发现[6]。由于一段时期以来假冒知名产地大闸蟹的事件频发[7-8],着力研究和开发可快速且非致死地辨别大闸蟹产地的办法对相关产业的有序发展意义重大。近年来,针对中华绒螯蟹的背甲(头胸甲)的研究越来越受到关注。该近六边形钙质结构硬组织的边缘总体上可分为前缘、眼缘、前侧缘、后侧缘和后缘5部分。前缘正中为额部,有4枚齿突,称额齿,额齿中间的凹陷,以中央的一个为最深;左右前侧缘各有4个锐齿,称侧齿;背甲后侧缘斜向内侧,后缘与腹部交界,比较平直;背甲中部有呈M型的花纹[9]。关注这些形态上的产地适应性变化有利于开展大闸蟹产地溯源和辨别的研究[7]。物种形态学变异可受遗传因素和环境因素等影响,在敞水和浅水环境中养殖湖沼型和底栖型太阳鱼(Lepomisgibbosus),比较其遗传变异和表型可塑性对鱼形态变化权重的研究发现,养殖环境差异所导致的表型可塑性形态学变异贡献率达53%,远大于由遗传分化所造成的贡献率(14%)[5]。因此养殖环境差异导致的形态学变异更需得到重视。
几何形态测量学是主要基于二维或三维形态结构上符合特定定义的同源地标点及其精密的数据系统,来破解形态差异性及其动态和可视化的学科[10]。该法所选取的地标点是具有明显特征且容易辨识的点,且后续可视化处理时可以消除在选取时会发生的位置、取向和比例上的非形态变化的干扰[11]。郑朝臣等[6]首次尝试引入该几何形态测量学分析法研究了中华绒螯蟹背甲及腹甲形态的产地差异性,发现基于雌、雄蟹背甲形态的产地判别准确率可高达100%,与以往通过地理元素“指纹”方式所能够达到的判别准确率[12-14]类似。近期,利用该法分别对不同国家和我国不同水系野生中华绒螯蟹的背甲形态特征差异所开展的研究也取得了较为满意的判别效果[15-18]。然而,基于几何形态测量学的大闸蟹外型及其产地(特别是同一水系,如长江)精细尺度的判别研究较少;其相关理论基础数据仍需完善,且其应用方面的可行性仍需进一步验证。鉴于此,笔者以长江水系中安徽、江苏7个产地中华绒螯蟹背甲作为研究对象,对其产地间的几何形态测量学差异性特征开展了较为详细的研究,以期为建立我国各知名产地中华绒螯蟹背甲的几何形态测量学鉴别手段提供理论支撑。
1 材料与方法
1.1 试验材料
试验所选用的长江水系中华绒螯蟹样本分别于2017年10月采自长江中游地区的湖南大通湖,2018年11月采自长江下游地区的安徽黄陂湖、女山湖、沱湖,2017年12月采自长江无为江段,2017年11月采自江苏溱湖、石臼湖7个产地(表1)。除无为江段的蟹为长江的野生个体外,其余蟹均为放养或围网养殖于相应湖区的个体(苗种均为购自江苏南通繁育场的长江水系扣蟹,种质资源遗传背景相同)。本试验中所采集各湖区产地的养殖蟹均符合国标GB/T 19957-2005所标准定义的原产成蟹(至少从扣蟹养到成蟹这一过程保持在同一水域内)的要求。至采样时,各产地蟹均为商品规格成蟹,已完成全部蜕皮过程[19]。各产地蟹采集后均尽快运回实验室,并活体进行常规及几何形态学测量。
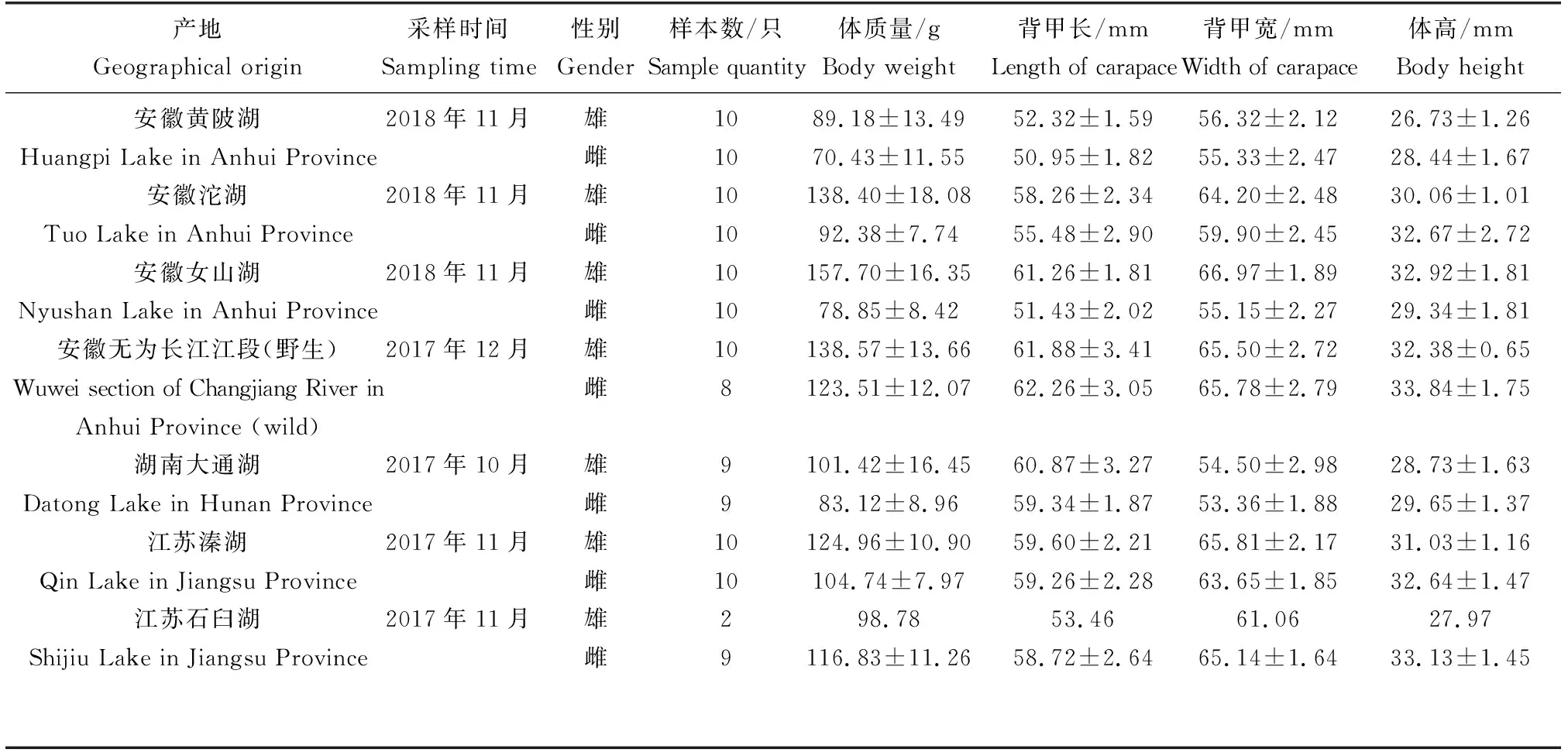
表1 长江水系7个不同产地中华绒螯蟹群体的形态测量信息
1.2 试验方法
1.2.1 地标点的建立与提取
地标点主要分为3类, 即以不同组织间交点为特征的Ⅰ型地标点, 以结构中凹陷或凸起点为特征的Ⅱ型地标点和以结构极值点为特征的Ⅲ型地标点[11]。依据几何形态测量学的地标点选取原则,在蟹背甲上选取了Ⅱ型地标点(33个)和Ⅲ型地标点(2个)共35个(图1、表2)。在LED灯光(6 W)条件下,利用三脚架设定拍摄固定高度(24 cm)的Nikon COOLPIX P6000数码相机,进行蟹背甲拍照。将拍好的照片导入Adobe Photoshop 13.0软件后,再利用tpsDig2软件[20]进行地标点的标记,获取地标点的x、y坐标值,建立二维地标点的数据文件。
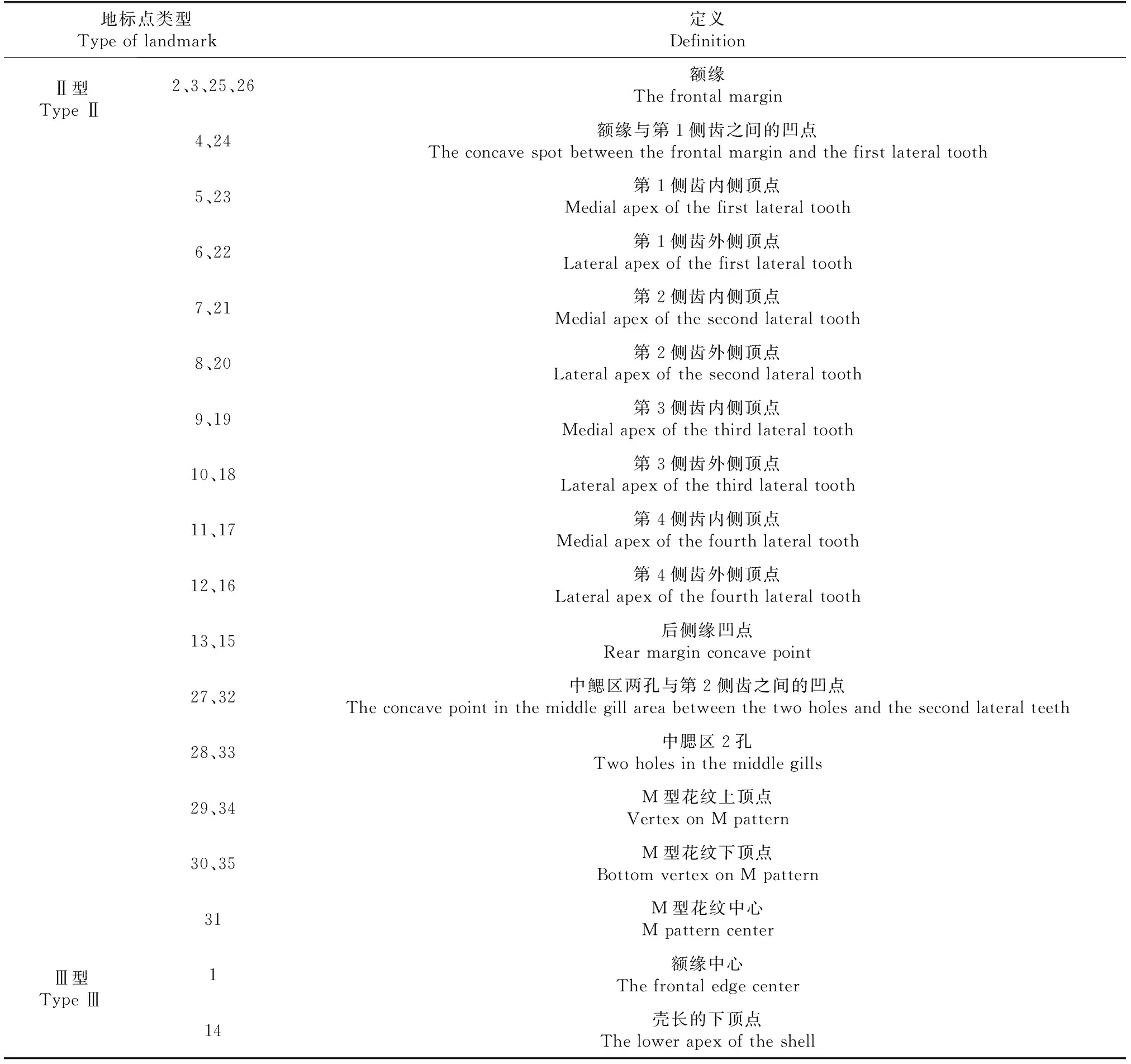
表2 地标点类型与定义
1.2.2 平均形与薄板样条分析
地标点的原始坐标数据包含了许多非形态变异。利用tpsRelw软件[21]普氏叠加运算来消除上述非形态变异,得到标准化数据并求出平均型,以用于后续的判别分析。依据标准化数据的相对扭曲分析获得35个地标点的贡献率[22]。利用tpsRegr软件[23]进行薄板样条分析[24-25],绘出各产地中华绒螯蟹网格变形图,进行差异比较。
1.2.3 判别分析
将经tpsRelw软件得到的相对扭曲得分导入Excel表格,采用Bayes方法[26]进行判别分析。将所有自变量纳入SPSS 23.0软件判别函数进行统计分析。
2 结果与分析
2.1 背甲形态的可视化差异分析
利用tpsRelw软件根据地标点数据文件计算得到背甲形态的平均形(图2),再通过绝对扭曲、回归分析和置换检验进行网格化和差异矢量化后构建网格图(图3、图4)。在对7个产地中华绒螯蟹背甲地标点数据进行的地标点法分析中,雄蟹贡献率最高的地标点分别为第11、12、16、17地标点,共解释了63.26%的形态差异,其他31个地标点解释了36.74%的形态差异;雌蟹贡献率最高的地标点分别为第11、12、16、17地标点,共解释了60.21%的形态差异,其他31个地标点解释了39.79%的形态差异。

图2 中华绒螯蟹背甲平均形Fig. 2 Average shape of the Chinese mitten crab E. sinensis carapace
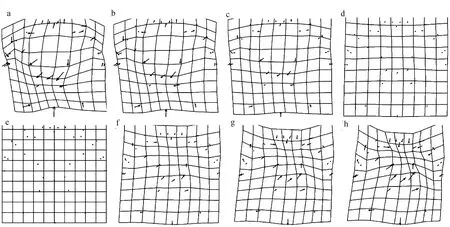
图3 中华绒螯蟹雄蟹背甲网格变形图与变异可视化(变异扩大10倍)Fig. 3 Mesh deformation map and variation visualization (with 10 times magnification) for the carapace of male E. sinensis from different geographical origins
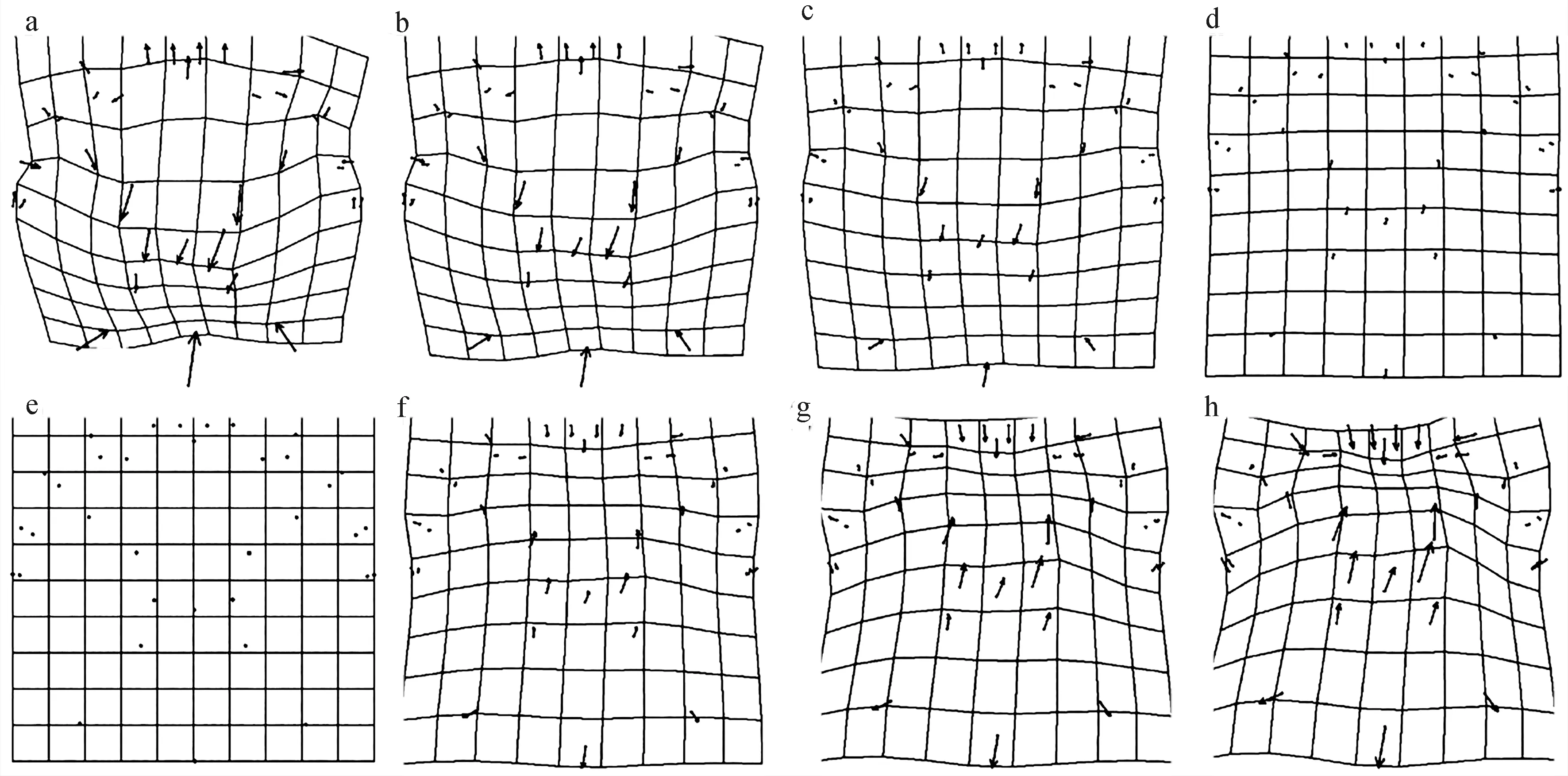
图4 中华绒螯蟹背甲网格变形图与变异可视化(变异扩大10倍)Fig. 4 Mesh deformation map and variation visualization (with 10 times magnification) for the carapace of female E. sinensis from different geographical origins
对雄蟹背甲(图3)而言,大通湖蟹、溱湖蟹、石臼湖蟹与平均形相比,其额缘(第2、3、25、26地标点)整体向下偏移,中腮区左侧孔(第28地标点)向右上伸展,中腮区右侧孔(第33地标点)向上方伸展。大通湖蟹的M型花纹上除下顶点(第30地标点)基本无变化外,其余花纹各点向右上方伸展,花纹变大变宽;而溱湖蟹和石臼湖蟹的M型花纹下顶点(第30地标点)向右上伸展,较花纹上其余各点(第29、31、34、35地标点)向右上方伸展程度小,花纹整体变大变宽。
黄陂湖蟹、女山湖蟹、沱湖蟹的中腮区左侧孔(第28地标点)向左下伸展,中腮区右侧孔(第33地标点)向下伸展。黄陂湖蟹、女山湖蟹的额缘(第2、3、25、26地标点)及额缘中心(第1地标点)与平均形相比整体向上平移,M型花纹上除下顶点(第30地标点)向左下方略微偏移,其余点均向M型中心聚集,花纹整体变小。但沱湖蟹的M型花纹上除下顶点(第30地标点)基本无变化外,其余各点(第29、31、34、35地标点)向M型中心聚集,花纹变小。
安徽无为长江江段的野生蟹和平均形相比,背甲形态则无明显的区别。
对雌蟹背甲(图4)而言,大通湖河蟹、溱湖蟹和石臼湖蟹的额缘(第2、3、25、26地标点)及额缘中心(第1地标点)与平均形相比整体向下平移,凹陷更大。中腮区两孔(第28、33地标点)与平均形相比向上偏移,M型花纹上各点整体向上平移,M型花纹斜竖向伸展,花纹整体更细长。后侧缘凹点(第13、15地标点)和壳长的下顶点(第14地标点)向左下移动,背甲底部变宽大。但其中石臼湖蟹的额缘与第1侧齿之间的凹点(第4、24 地标点)分别向内侧移动,与额缘之间距离变小。第四侧齿内侧顶点(第11、17地标点)、第四侧齿外侧顶点(第12、16地标点)向内下方缩短,侧齿变小。
黄陂湖蟹、女山湖蟹和沱湖蟹的额缘(第2、3、25、26地标点)及额缘中心(第1地标点)与平均形相比整体向上平移,顶端更尖细。中腮区2孔(第28、33地标点)与M型花纹上各点整体向下平移,后侧缘凹点(第13、15地标点)和壳长的下顶点(第14地标点)向M型花纹方向移动,两者之间距离缩短。
安徽无为长江江段的野生蟹和平均形相比,背甲形态亦无明显变化。
2.2 背甲形态差异的判别分析
对7个产地间雌、雄蟹的背甲进行判别,准确率均高达100%。判别分析散点图(图5)更直观地显示出蟹背甲形态地标点法的应用结果在视图上也能明显且有效地显示出判别结果良好。
将每个产地的雌、雄蟹作为一个整体,进行仅7个产地间中华绒螯蟹背甲几何形态的判别分析,准确率为98.4%(表3),判别分析散点图(图6)显示判别效果差于区分性别的判别结果。

表3 基于背甲形态的7产地中华绒螯蟹判别分析
3 讨 论
3.1 中华绒螯蟹背甲的几何形态测量学产地差异特征
笔者利用基于地标点的几何形态测量法来判别同属长江水系7个不同产地养殖和野生中华绒螯蟹形态差异,取得了满意的结果。试验结果表明,湖南、安徽、江苏3个省份间和各省内不同产地间蟹的背甲形态上均存在差异。与平均型相比较:安徽黄陂湖蟹、女山湖蟹、沱湖蟹背甲形态有同趋势变化,额齿和侧齿更尖长,M型花纹变小,背甲后缘缩小;湖南大通湖蟹和江苏溱湖蟹、石臼湖蟹背甲形态有同趋势变化,额齿和侧齿变短,M型花纹变大,背甲后缘变宽大;安徽无为长江江段野生蟹背甲形态则与蟹的平均形态差异不显著。7个产地间雌、雄蟹的背甲形态判别准确率均可达100%。关于长江水系太湖、洪泽湖、高宝湖、长荡湖、阳澄湖,巴城、兴化“泓膏”养殖水域,长江口崇明自然水域等产区所产蟹的背甲形态的研究结果同样表明,中华绒螯蟹背甲的第1侧齿、M型花纹以及背甲底部存在显著差异[6]。关于长江、黄河、辽河、瓯江、闽江及图们江野生中华绒螯蟹和荷兰野生中华绒螯蟹背甲形态特征的研究结果[15-16]也与上述发现一致。
本研究中不同产地间中华绒螯蟹体质量有一些明显的差异。但至9月之后个体均为商品规格的成蟹,对背甲产地适应性变化影响最大的蜕皮过程已全部完成,故此阶段养殖过程中背甲形态对产地环境的适应性变化已趋于稳定。已有研究表明,虽然不同产地蟹的体质量有差异,但未观察到对背甲形态产地差异性的影响[6-7,18-19]。因此,笔者认为蟹体质量上的差异将不会影响分析的结果和准确性。
同样,遗传因素亦不会影响到本研究结果的客观性。由于本研究中无为群体本身就是长江野生蟹,而另6个养殖产地苗种均为购自江苏南通繁育场的长江水系扣蟹,故相关7个产地蟹的种质资源遗传背景相同,不存在遗传多样性导致背甲形态差异性的问题。不同生境应该是导致其产出中华绒螯蟹背甲形态差异的最主要因素。黄朝辉等[27]在关于三疣梭子蟹(Portunustrituberculatus)蜕壳周期肝胰腺、外壳和鳃中钙含量的变化研究中发现,当蟹壳完全硬化后(从完全硬化至下一次蜕壳期间),蟹壳中钙含量将不再发生变化,其新壳硬化所需的钙主要来自周围环境;Greenaway[28]也综述了甲壳动物中的这种现象。蟹壳中含有碳酸钙、甲壳质等物质[29],在蜕皮时会吸收大量水分及水中的无机盐。不同生活环境中不同的矿质元素含量及比例可以使蟹背甲在生长过程中发生细微变化。中华绒螯蟹相关深层次机制性的原因仍需今后开展精准且有针对性的试验来论证。
本研究中雌、雄蟹的背甲形态之间也存在细微差异。Rufino等[30]对梭子蟹(Liocarcinusdepurator)的研究也发现,雄性和雌性之间的背甲形态有明显的差异,雌蟹的腹部底部较长,也更凹,而雄蟹背甲较宽;Snchez等[31]的研究也表明,同种生物的不同种群在不同性别之间会因生存环境的不同而产生形态上的变化。在区分蟹性别研究的基础上,将单个产地雌、雄蟹作为整体,进行不区分性别的产地判别,结果显示存在一定程度的误判,判别准确率为98.4%,低于区分性别时100%的判别结果,虽然判别结果仍然具有较好的区分度,但是从散点图可以发现,性别的差异对产地的差异产生了极大的影响,各产地样本的分散程度大,出现了较多的重叠现象,散点图中的区分度低于区分性别时的结果。因此,在进行不同产地之间中华绒螯蟹背甲几何形态分析时,宜将蟹性别区分开进行研究。这也说明中华绒螯蟹在生长过程中,其背甲会因性别不同和生存地理环境的不同产生形态的差异,并形成与其栖息地相适应的变化。因此,在进行不同产地之间中华绒螯蟹背甲几何形态分析时,宜将蟹区分性别来进行研究,才能获得合理的效果。这也说明中华绒螯蟹在生长过程中,其背甲形成与其栖息地相适应的变化还会受到性别的影响,并很可能产生背甲形态的性二型现象。中华绒螯蟹背甲形态的这些特征性变化为基于地标点的几何形态测量学的发展提供了潜在价值,为不同产地中华绒螯蟹形态品质的评价提供了重要理论依据。
3.2 中华绒螯蟹背甲的几何形态测量学产地判别的实用性
传统的形态测量方法(如框架法等)往往不适用于淡水蟹类的鉴定[32],因为它们的外部形态变化无常。在软体动物的研究中,苏杭[33]通过不同形态学测量方法对柔鱼(Ommastrephidebartramii)和茎柔鱼(Dosidicusgigas)的群体鉴别和差异性进行分析,研究发现,相对于传统测量学法,几何形态测量学中地标点法具有更好的判别效果;在鱼类的研究中,Ponton[34]也比较了传统测量法、地标点几何形态测量法等区分印度小公鱼(Stolephorusindicus)、戴氏半棱鳀(Encrasicholinadevisi)、尖吻半稜鳀(E.heteroloba)、刺公鳀(E.punctifer)等4种鱼耳石的有效性,其结果同样表明,地标点几何形态测量法比传统测量法和其他方法更加有效,并且能够可视化地表达出耳石随着生长形态变异的趋向。作为相关产地初步的探索,由于受到所采样本数的限制,来自江苏石臼湖的雄蟹仅有2只,今后需要采集更多的个体来开展更深入的研究。
笔者选取的雌、雄中华绒螯蟹背甲地标点的切空间距离和普氏距离的回归系数分别为0.999 808和0.999 838,均接近于1,这说明选取的地标点是有效的,且雌、雄蟹背甲判别准确率均为100%。但在姜晓东等[15]关于几何形态测量学区分不同水系野生中华绒螯蟹的研究中,各群体中华绒螯蟹的平均判别准确率仅为87.67%,其选取的地标点个数为30个,包含22个Ⅲ型地标点,7个Ⅱ型地标点;而笔者选取了35个地标点,包含33个Ⅱ型地标点,2个Ⅲ型地标点,且贡献率高的均为Ⅱ型地标点(第11、12、16、17地标点),产地判别准确率也均高达100%。Zheng等[18]关于长江水系部分湖泊、黄河水系的东营地区以及辽河水系的营口和盘锦地区的中华绒螯蟹背甲几何形态的研究结果也表明,以Ⅱ型为主的地标点,其产地区分成功率高达100%。这说明对于中华绒螯蟹背甲几何形态研究方面,地标点选取Ⅱ型为宜。此外,地标点的数量不宜过少,避免造成信息的缺失;也不宜过多,避免多余贡献率低的坐标点,增大试验误差。因此,对不同水生生物开展基于地标点的几何形态测量学法的研究时不能一概而论,要特别关注其形态的物种特点和相应地标点类型的组成特征来进行。
综上所述,基于地标点法的几何形态测量学在中华绒螯蟹产地判别的实用性方面潜力显著且切实可行。相较于通过稳定同位素等分析法[35]判别蟹类产地差异性而言,地标点的几何形态测量学方法有效地解决了需要致死蟹才能进行试验的问题。且该方法还具有成本低、耗时少、结果准确性高的优势。因此,基于地标点的几何形态测量法可为中华绒螯蟹等生物的产地辨别、原产地溯源及保护工作提供更便捷有效方法和非致死的创新途径。
迄今,通过相同的基于地标点法的几何形态测量学方法,本实验室已对诸多产地的中华绒螯蟹进行了背甲形态差异的鉴别[6-7],同时也对阳澄湖“洗澡”蟹与原产蟹背甲形态进行了成功的鉴别[7]。本研究结果又进一步拓展了该法的应用及其相关的数据库,扩充到了更多省份及湖区产地的绒螯蟹,也进一步证实了该方法在同一水系精细尺度产地间中华绒螯蟹相应形态差异判别的可行性。
当前,中华绒螯蟹产业已成为乡村振兴中坚持生态优先和绿色发展,用技术创新提升特色产业品质的重要抓手。近些年江苏阳澄湖等地区已建立了中华绒螯蟹的原产地保护、地理标识等高质量发展的制度[36]。在今后的研究中,还需强化几何形态测量学相关技术的研究,扩展其在判别知名产区来源高质中华绒螯蟹的可靠性等领域的应用,并注重所获判别方法及其产地溯源可行性的再确认工作,以便以新角度为上述制度的有效执行做出贡献,也可为中华绒螯蟹产业乡村振兴、特色强农战略提供理论和技术支持。
4 结 论
长江中下游7个产地的中华绒螯蟹背甲形态存在明显的差异。基于背甲地标点的几何形态测量学法对产地间雌、雄蟹个体的判别准确率均可达100%。背甲几何形态测量学法可为精细尺度产区中华绒螯蟹原产地溯源提供更为便捷且非致死的技术途径。
