金钱鱼agrp基因cDNA克隆及饥饿与复投喂对其表达的影响
2023-09-27蔡博生陈华慧罗翠婷李广丽邓思平
蔡博生,陈华慧,罗翠婷,翟 毅,李广丽,邓思平
( 1.广东海洋大学 水产学院,广东 湛江 524088; 2.广东省名特优鱼类生殖调控与繁育工程技术研究中心,广东 湛江 524088 )
下丘脑刺鼠相关蛋白/神经肽Y(AgRP)神经元,是摄食调节的关键中枢。AgRP是AgRP神经元分泌的一种神经肽,是一种作用较强的食欲刺激因子[1-2]。在机体能量缺乏时,下丘脑内AgRP神经元通过感知神经信号和代谢信号被激活,agrp基因过表达合成AgRP并参与瘦素信号传导,进而影响摄食行为,实现食欲调节和能量平衡的中枢调控[3-6]。
目前,对AgRP的研究主要集中在哺乳动物。在四足动物中发现,agrp为单拷贝基因,而在硬骨鱼类中普遍存在2种agrp亚型基因[7]。在人和家鼠中,agrp基因主要在下丘脑中表达[8]。鸟类的agrp基因主要在下丘脑、毛囊和皮肤中表达[9]。硬骨鱼类中agrp基因的不同亚型在组织表达上存在差异,有的在脑中高表达,有的在鳃和性腺中高表达[10-11]。AgRP在食欲和体质量的调控中起着重要作用。研究证实,药物损伤成年小鼠(Musmusculus)的AgRP神经元或向其脑内注射AgRP会导致摄食行为丧失或摄食量显著增加[12-13]。近年来,关于AgRP在鱼类摄食调控中的研究相继被报道。在斑马鱼(Daniorerio)中,损伤AgRP1神经元可导致斑马鱼食物消耗量显著减少[14];禁食状态下,斑马鱼下丘脑中AgRP1神经元数量会增加并参与食欲调节[15]。AgRP参与了欧洲舌齿鲈(Dicentrarchuslabrax)[16]和团头鲂(Megalobramaamblycephala)[17]的摄食调节,长期饥饿状态下agrp基因的表达量均显著上调。
金钱鱼(Scatophagusargus)属鲈形目金钱鱼科金钱鱼属。金钱鱼具有极强的环境适应性和抗逆性,但生长较为缓慢,是我国南海的一种名特优鱼种。目前,在金钱鱼的生物学[18]、性别调控[19]、生长调控[20-21]方面已有了初步的研究。也有学者对金钱鱼下丘脑神经肽Phoenixin[22]和Spexin[23]在摄食调节中的作用进行了初步探讨:AgRP也是鱼类下丘脑分泌的一种神经肽,但其在金钱鱼摄食调节中的作用尚未见报道。为探明AgRP在金钱鱼摄食调节中的作用,笔者克隆了金钱鱼agrp1基因和agrp2基因,采用实时荧光定量PCR检测这2种agrp基因在金钱鱼各组织中的分布和饥饿及复投喂处理后其在下丘脑、肠道和肝脏中的表达水平变化情况,旨在为阐明金钱鱼下丘脑神经肽在摄食调节中的作用以及为该鱼的人工养殖提供理论依据。
1 材料与方法
1.1 试验材料
基因克隆所用的金钱鱼购自广东省湛江市霞山区水产品批发市场。组织表达和饥饿及复投喂试验所用金钱鱼均来自广东海洋大学东海岛海洋生物研究所基地。所有试验鱼取样前用间氨基苯甲酸乙脂乙磺酸盐麻醉,并置于冰上取所需组织,组织取出后立刻投入液氮中保存用于总RNA提取、反转录和基因表达。
1.2 试验方法
1.2.1 总RNA提取及反转录
总RNA提取参照Trizol®reagent(Invitrogen,USA)试剂盒的说明书操作,用1%琼脂糖凝胶电泳检测片段RNA完整性和采用核酸蛋白测定仪测定RNA浓度。采用PrimeScriptTMRT reagent Kit with gDNA Eraser (TaKaRa)合成第1链cDNA。
1.2.2 基因克隆
基于大黄鱼(Larimichthyscrocea)agrp基因,在金钱鱼转录组数据库中筛选出与其相似度较高的unigene片段,根据筛选的序列,利用Primer premier 6.0设计特异性引物对(agrp1-F: ATGTTTGGCTCTGTGCTGC, agrp1-R: GCAGCACAGAGCCAAACAT; agrp2-F: ATGTTTGGCT CTGTGCTGC, agrp2-R: GCAGCACAGAGCCAAACAT)。以下丘脑cDNA为模板进行PCR扩增:94 ℃变性30 s,56 ℃退火30 s,72 ℃延伸1 min,33个循环。PCR扩增结束后,取5 μL PCR产物在1.0%琼脂糖凝胶电泳中检测。采用琼脂糖回收试剂盒回收、纯化目的片段:将回收纯化后的产物与Peasy-T3载体连接,使用溶菌肉汤培养基进行培养,挑取单克隆的菌落送至生工生物工程(上海)股份有限公司测序,所有的引物均由生工生物工程(上海)股份有限公司合成。
1.2.3 序列分析和聚类分析
运用DNAMAN 8.0软件对所得到的测序结果进行剪切拼接,获得推导的金钱鱼agrp1、agrp2基因的开放阅读框和氨基酸序列。以Signal IP 4.1(http://www.cbs.dtu.dk/services/SignalP/)预测金钱鱼agrp信号肽。由美国生物技术信息中心数据库获得其余物种的AgRP1、AgRP2的氨基酸序列。利用DNAstar软件对金钱鱼与其他AgRP氨基酸进行同源性分析。运用MEGA 8.0邻位相接法,基于AgRP1、AgRP2氨基酸序列构建系统进化树。
1.2.4 组织分布
自养殖池塘随机挑选3尾试验鱼立即取下丘脑、垂体、鳃、肝脏、心脏、脾脏、肾脏、肠道、头肾、肌肉和胃11个组织,置于液氮保存,用于总RNA提取和反转录,获得组织表达的cDNA模板。以金钱鱼agrp1和agrp2基因 (特异性引物:agrp1-F1,CT GTGCTGCTGTGCTGTT; agrp1-R1,CAAGGCT GCTGATGAGTC。agrp2-F1,AGCGATGAGGACCG-ACAA;agrp2-R1,GCAGTGGCAGATGGTGTT)为目的基因,β-actin基因(β-actin-F: GAGAGGTTCCGTTGCCCAGAG; β-actin-R: CAGACAGCA-CAGTGTTGGCGT)为内参基因,采用ABI 7500 PCR仪,SYBR Green Realtime PCR Master mix试剂盒(TOYOBO,日本)操作说明建立反应体系,实时荧光定量PCR检测各组织中的表达情况。实时荧光定量PCR的反应条件:95 ℃预变性1 min;94 ℃变性15 s,56 ℃退火15 s,72 ℃延伸45 s并收集荧光信号,40个循环;然后进行熔解曲线分析。
1.2.5 饥饿及复投喂对agrp1和agrp2基因表达水平的影响
选取体质量52~60 g的金钱鱼用于饥饿及复投喂试验。将选取的金钱鱼随机分为正常投喂组、饥饿组和复投喂组。正常投喂组(对照组)每日定量(饵料为鱼体质量的2%)、定时(9:00)投喂(饵料购于中国悦群海洋生物研究开发公司),连续投喂7 d;饥饿组分别在饥饿2 d和7 d后取样;复投喂组为饥饿7 d后再投喂,并在投喂3 h后取样。饥饿组和复投喂组在每个取样时间点均取3尾试验鱼的下丘脑、肝脏和肠道用于总RNA提取和反转录。实时荧光定量PCR检测饥饿和复投喂后agrp1和agrp2基因表达情况,实时荧光定量PCR方法与组织分布所用的方法相同。
1.2.6 数据分析
每个样品进行2次实时荧光定量PCR,取2次的平均Ct值,运用2-ΔΔCt计算agrp1和agrp2基因相对表达量。所有数据均以平均值±标准差表示。利用SPSS 19.0软件进行单因素方差分析,采用邓肯多重比较进行各组之间agrp1和agrp2基因的表达量差异比较,当P<0.05时,各组间具有显著性差异。
2 结 果
2.1 agrp基因序列分析与系统进化树
从金钱鱼转录组数据库中得到agrp1和agrp2基因的unigene片段,经过亚克隆测序验证后,得到金钱鱼agrp1(GenBank登录号:MW988084)和agrp2(GenBank登录号:MW988085)基因的开放阅读框序列。其中agrp1基因的开放阅读框的长度为429 bp,共编码142个氨基酸,预测其编码的蛋白质的分子质量为36.73 ku,等电点为5.16,蛋白质N端1~24位氨基酸为信号肽,第24~25位为切割位点(图1);agrp2基因的开放阅读框的长度为351 bp,共编码116个氨基酸,预测其编码的蛋白质的分子质量为64.19 ku,等电点为8.32,蛋白质N端1~22位氨基酸为信号肽,第22~23位为切割位点(图2)。预测的金钱鱼AgRP1和AgRP2具有10个半胱氨酸残基,并且形成5个保守的二硫键(图3)。

图1 金钱鱼agrp1基因的cDNA序列及其推导的氨基酸序列Fig.1 cDNA sequence and the corresponding amino acid sequence of the S. argus agrp1 gene

图2 金钱鱼agrp2基因的cDNA序列及推导的氨基酸序列Fig.2 cDNA sequence and the corresponding amino acid sequence of the S. argus agrp2 gene
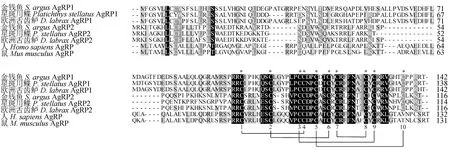
图3 不同物种AgRP氨基酸序列多重比较Fig.3 Multiple alignment of the amino acid sequences of AgRP in different species
同源性比较发现,金钱鱼的AgRP1和AgRP2 2种亚型的同源性为24.6%。在所选取的物种中,金钱鱼AgRP1氨基酸序列与其他硬骨鱼类的AgRP1同源性较高(70.4%~87.3%),金钱鱼AgRP2氨基酸序列与其他硬骨鱼类的AgRP2同源性较高(66.1%~86.2%)。金钱鱼AgRP1与日本花鲈(Lateolabraxjaponicus) AgRP1、欧洲舌齿鲈(Dicentrarchuslabrax) AgRP1、金头鲷(Sparusaurata)AgRP1、大菱鲆(Scophthalmusmaximus) AgRP1、红鳍东方鲀(Takifugurubripes) AgRP1的同源性分别为87.3%、86.6%、85.2%、83.0%和70.4%。金钱鱼AgRP2与欧洲舌齿鲈AgRP2、日本花鲈AgRP2、金头鲷AgRP-like、大菱鲆AgRP2、红鳍东方鲀AgRP2的同源性分别为86.2%、84.5%、83.6%、73.9%、66.1%。系统进化树聚类结果显示,AgRP在进化上分化为两类,其中AgRP1与其他硬骨鱼类的AgRP1和AgRP聚为一类,AgRP2则与其他硬骨鱼类的AgRP2和AgRP-like聚为一类(图4)。

图4 基于AgRP1、AgRP2氨基酸序列构建金钱鱼与其他硬骨鱼类的AgRP系统进化树Fig.4 Phylogenetic analysis of the AgRP proteins in the butterfish S. argus and other teleosts
2.2 agrp基因在各组织中的表达
采用实时荧光定量PCR检测agrp1和agrp2基因在金钱鱼各组织的表达。agrp1基因在金钱鱼各组织中均有表达,其中下丘脑和肌肉的表达量最高,其次在垂体、鳃、肝脏、心脏、脾脏和肠道等组织中也检测到其表达(图5a);而agrp2基因在下丘脑和垂体中表达较高,在鳃和心脏等组织中也检测到其表达(图5b)。
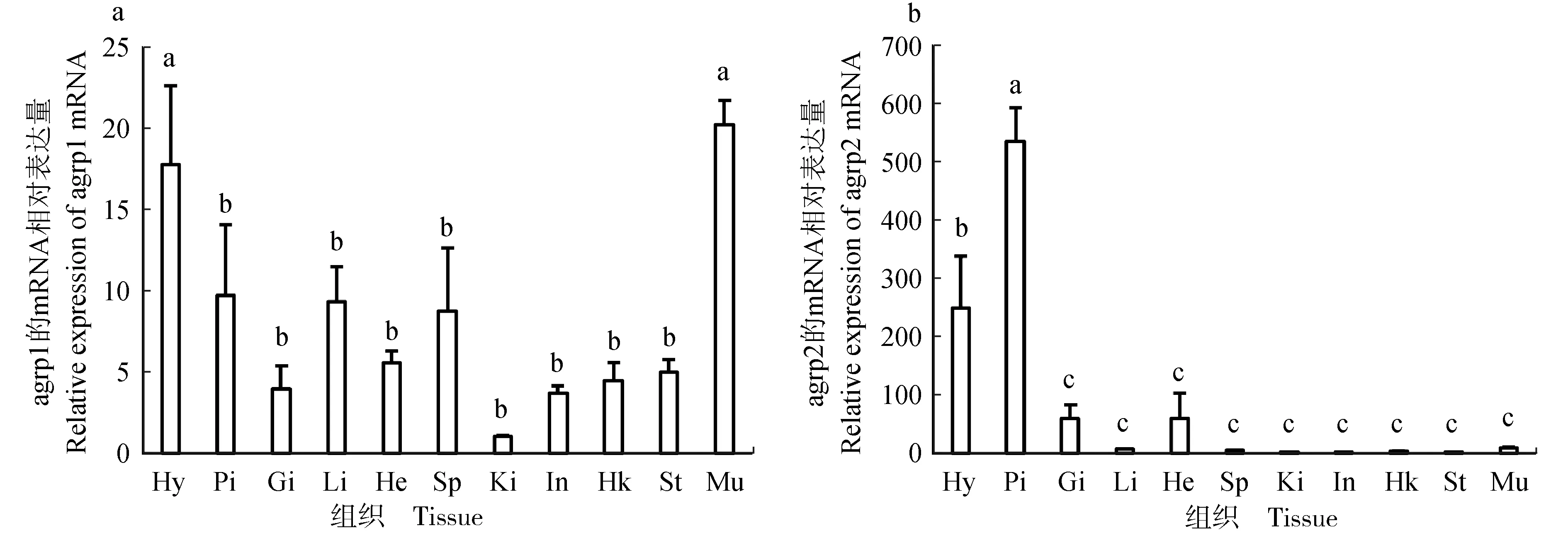
图5 实时荧光定量PCR检测金钱鱼组织中agrp1(a)与agrp2(b)基因的表达情况Fig.5 Tissue distribution of the agrp1 (a) and agrp2 (b) in butterfish S. argus by qPCR
2.3 饥饿及复投喂对下丘脑、肝脏和肠道中agrp基因表达的影响
饥饿及复投喂试验结果表明:下丘脑中,饥饿2 d后,agrp1基因表达水平与对照组相比差异不显著(P>0.05),饥饿7 d后,agrp1基因表达水平与对照组相比显著下调(P<0.05),饥饿7 d后再进行复投喂,发现agrp1基因表达水平上调到正常水平(图6a);agrp2基因无论饥饿2、7 d还是复投喂后,其基因表达水平与对照组相比均无显著变化(P>0.05)(图6b)。肝脏中,饥饿2 d后,agrp1基因表达水平与对照组相比显著下调(P<0.05),饥饿7 d后,agrp1基因表达水平显著上调(P<0.05),复投喂后,agrp1基因表达水平与对照组相比差异不显著(P>0.05)(图7a);agrp2基因无论饥饿2、7 d还是复投喂后,其基因表达水平与对照组相比无显著性变化(P>0.05)(图7b)。在肠道中,饥饿2 d后,agrp1基因表达水平与对照组相比无显著性变化(P>0.05),饥饿7 d后,agrp1基因表达水平显著上调(P<0.05),复投喂后,agrp1基因表达水平下调到正常水平(图8a);agrp2基因无论饥饿2、7 d还是复投喂后,其基因表达水平与对照组相比均无显著性变化(P>0.05)(图8b)。
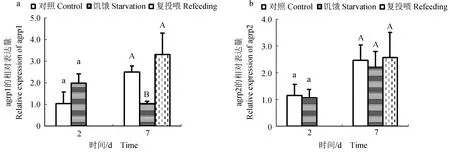
图6 饥饿及复投喂对金钱鱼下丘脑中agrp1(a)和agrp2(b)基因表达的影响Fig.6 Effects of starvation and refeeding on the expression of agrp1(a) and agrp2(b) in the hypothalamus of the butterfish S. argus
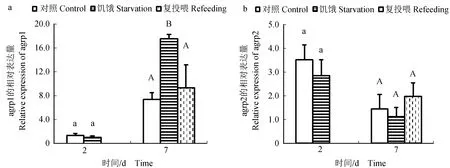
图8 饥饿及复投喂对金钱鱼肠道中agrp1(a)和agrp2(b)基因表达的影响Fig.8 Effects of starvation and refeeding on the expression of agrp1(a) and agrp2(b) in the intestine of buterfish S. argus
3 讨 论
3.1 金钱鱼agrp基因序列、编码氨基酸序列比对及系统进化分析
AgRP属于Agouti信号蛋白家族的一员,其C端具有一个富含半胱氨酸残基的区域。2个半胱氨酸残基之间形成1个二硫键,而二硫键对于蛋白质三级结构的稳定性具有重要作用[24]。本试验结果显示,金钱鱼2种亚型AgRP在C端均具有10个半胱氨酸残基,并形成5个二硫键。这一特征与金鱼(Carassiusauratus)[25]、齐口裂腹鱼(Schizothoraxprenanti)[26]、欧洲舌齿鲈[16]、红鳍东方鲀[27]中的情况一致,且该结构域高度保守,由此推测该结构域在不同物种间可能具有相似的功能。
从美国国家生物技术信息中心数据库以及现有的文献发现,硬骨鱼类agrp基因命名方式主要有3种:第1种是agrp1和agrp2,如日本花鲈等;第2种是agrp-like和agrp,如攀鲈(Anabastestudineus)、大黄鱼和尖吻鲈(Latescalcarifer)等;第3种是agrp1和agrp,如金头鲷等。氨基酸序列系统进化树显示,金钱鱼AgRP1和AgRP2明显聚为两支,其中金钱鱼AgRP1与其他硬骨鱼类AgRP1、AgRP-like聚为一类,金钱鱼AgRP2与其他硬骨鱼类AgRP2、AgRP聚为一类。金钱鱼AgRP1与其他硬骨鱼类的AgRP1、AgRP-like同源性较高(66.9%~87.3%),与其他硬骨鱼类AgRP2、AgRP同源性较低(19.7%~24.1%);金钱鱼AgRP2氨基酸序列与其他硬骨鱼类的AgRP2、AgRP同源性较高(59.0%~86.2%),与其他硬骨鱼类AgRP1、AgRP-like同源性较低(20.20%~26.67%);此外,金钱鱼AgRP1与金钱鱼AgRP2的同源性较低,只有15.2%。系统进化树以及同源性比对结果表明,金钱鱼AgRP1与其他硬骨鱼类AgRP1和AgRP-like属于同一亚型,而金钱鱼AgRP2与其他硬骨鱼类的AgRP2、AgRP属于另一种亚型。因此本试验中,将金钱鱼agrp基因的2种亚型命名为agrp1和agrp2。
3.2 金钱鱼agrp基因在不同组织中表达的分析
在哺乳动物中,agrp基因主要在下丘脑中的弓状核表达,通过拮抗黑素皮质素-4受体的α-MSH效应,从而参与生物体的摄食调节[28-29]。金钱鱼的组织分布结果显示,agrp1和agrp2基因均在下丘脑中有高表达,这与齐口裂腹鱼[24]、红鳍东方鲀[27]、斑马鱼[15]和金鱼[25]的情况吻合。研究结果提示,与哺乳动物相似,鱼类的下丘脑可能也是调节食欲和能量平衡的关键区域。在金钱鱼中,agrp1基因还在垂体和肌肉中检测到高表达,agrp2基因在垂体中检测到高表达。在其他鱼类欧洲舌齿鲈中,agrp1基因主要在脑、卵巢和后肾中表达,agrp2基因主要在脑中表达[16];在大西洋鲑中,agrp1基因主要在脑垂体和皮肤中表达,agrp2基因主要在肠道、肌肉和性腺中表达[11];在鲤(Cyprinuscarpio)中,agrp1基因在脑、眼和肠道中高表达,而agrp2基因在鳃和眼中高表达[10]。这些结果表明,agrp的2种亚型基因的表达在鱼类存在组织表达差异和物种差异,而且在不同鱼类之间也存在物种差异,这种差异性表达需要进一步深入研究才能阐明其原因。
3.3 饥饿及复投喂状态下金钱鱼agrp基因表达分析
已有研究表明,AgRP1主要参与摄食行为的调节,AgRP2主要作为神经内分泌因子参与应激调节[14]。本试验结果显示,金钱鱼下丘脑中agrp1基因表达水平在饥饿7 d后下调。与金钱鱼类似,禁食6 d的大西洋鲑agrp1基因表达水平下调[11];团头鲂短期饥饿后,脑组织中agrp基因表达水平先降后升再降[17];鲤短期饥饿后,其agrp基因表达水平下调[10]。饥饿2 d后,斑马鱼agrp基因表达水平上调,复投喂后迅速恢复到对照组水平[15];与斑马鱼类似,金钱鱼下丘脑中agrp1基因表达水平在复投喂3 h后也恢复到正常水平。鱼类禁食后agrp1基因表达的下调可能与饥饿期间瘦素表达上升,AgRP表达受到抑制有关[30]。但金钱鱼中饥饿导致的agrp1基因表达下调是否与瘦素抑制有关,需要进一步深入研究。与金钱鱼不同,短期饥饿会上调金鱼[25]和欧洲舌齿鲈[16]agrp基因的表达。这种不同物种中AgRP在摄食调节中的差异性表明,在短期饥饿情况下,生物体内存在某种机制使得原本正常分泌的AgRP减少,以适应环境的变化[31],但具体机理有待进一步研究。食物缺乏时,鲈(Moronesaxatilis)[32]和露斯塔野鲮(Labeorohita)[33]肝脏中的瘦素分泌减少。在小鼠中发现,瘦素分泌减少可导致AgRP的分泌增加[34]。金钱鱼agrp1基因在肝脏和肠道中的表达模式与下丘脑中的相反,饥饿期间agrp1基因表达量显著上调,复投喂后显著下调到正常水平。饥饿和复投喂中,金钱鱼肝脏和肠道中agrp1基因的这种变化是否受到瘦素表达水平的影响有待进一步深入研究。在金钱鱼下丘脑、肝脏和肠道中agrp2基因表达水平在饥饿及复投喂过程均无显著变化。在欧洲舌齿鲈[16]和大西洋鲑[11]中均发现,饥饿对agrp2基因表达水平影响不大,这提示AgRP2可能不调节金钱鱼摄食[14]。在本研究中,金钱鱼下丘脑、肝脏和肠道中agrp1基因在饥饿及复投喂后的表达具有显著的变化,这表明参与金钱鱼摄食过程的主要是AgRP1。但鱼类的摄食调节是一个复杂的过程,AgRP在金钱鱼摄食调节中的作用及作用机制有待进一步证实。
4 结 论
笔者克隆了金钱鱼agrp1和agrp2 2种亚型基因,agrp1基因主要在下丘脑和肌肉中表达,agrp2基因主要在下丘脑和脑垂体中表达,饥饿及复投喂显著影响了金钱鱼下丘脑、肝脏和肠道组织中agrp1基因的表达。这些结果表明,下丘脑、肝脏和肠道中的agrp1基因参与了金钱鱼摄食过程的调节。
