亚热带森林生物多样性与生态系统功能实验研究基地 (BEF-China)研究进展
2023-09-21李珊刘晓娟马克平
李珊 刘晓娟 马克平

摘要:生物多样性与生态系统功能的关系 (BEF) 及其内在机制是当前生物多样性研究领域的热点问题。长期以来,以草地生态系统为主的BEF研究积累了大量研究成果,而基于森林生态系统的相关研究则相对较少。亚热带森林生物多样性与生态系统功能实验研究基地 (BEF-China) 是目前包含树种最多、涉及多样性水平最高的大型森林控制实验样地。该文总结了基于BEF-China平台的研究进展,特别是生物多样性对生态系统生产力、养分循环以及多营养级相互作用关系等方面的影响,并提出了未来BEF-China的研究应注重高通量测序和遥感等新兴技术的应用,在生物多样性的多维度、生态系统的多种组分与多种功能以及BEF研究的多种尺度等交叉方向上持续开展深入研究。针对BEF-China研究成果的梳理有助于理解驱动亚热带森林生物多样性与生态系统功能关系的内在机理,为生物多样性保护和生态修复提供科学依据。
关键词: 生物多样性, 生态系统功能, BEF-China, 亚热带森林, 研究综述
中图分类号:Q948 文献标识码:A 文章编号:1000-3142(2023)08-1524-13
Research progress of Biodiversity-Ecosystem Functioning Experiment China Platform (BEF-China)
LI Shan1,2,3*, LIU Xiaojuan1,2,3, MA Keping1,2,3
( 1. State Key Laboratory of Vegetation and Environmental Change, Institute of Botany,Chinese Academy of Sciences, Beijing 100093, China; 2. Zhejiang Qianjiangyuan Forest Biodiversity National Observation and Research Station, Quzhou 324000, Zhejiang, China; 3. China National Botanical Garden, Beijing 100093, China )
Abstract:The relationship between biodiversity and ecosystem functioning (BEF) and the underlying mechanisms have been a hot issue in ecological research. Whereas many BEF studies have focused on grassland ecosystems, relatively few have been conducted in forest ecosystems which account for a large proportion of primary production in the terrestrial ecosystem. Biodiversity-Ecosystem Functioning Experiment China Platform (BEF-China) contains the most tree species with the highest level of diversity in subtropical forests. In this review, we first summarized research progress in BEF-China, and then gave suggestions for future BEF research. The research based on BEF-China mainly focused on the impact of biodiversity on multi-ecosystem functioning, e.g. productivity, nutrient cycling and multi-trophic interaction. We suggest that future studies in BEF-China should strengthen the application of new techniques such as high-throughout sequencing and remote sensing, and continue research on multi-dimensions, multi-components, multi-ecosystem functions, and multi-scales in BEF study. The research progress of BEF-China can help in better understanding the BEF relationships in forest ecosystem, and provide a scientific support for biodiversity conservation and ecological restoration.
Key words: biodiversity, ecosystem functioning, BEF-China, subtropical forest, research summary
在世界范圍内,森林为各种各样的植物、动物和微生物提供了栖息地,并为人类福祉提供多种重要的生态系统服务。基于草地生态系统的生物多样性与生态系统功能 (biodiversity and ecosystem functioning, BEF) 研究结果表明,植物物种多样性对植物生产力有促进作用(Cardinale et al., 2012)。物种多样性通过调节生态系统的养分循环、多营养级相互作用等途径,从而影响生态系统的生产力及稳定性 (Ma et al., 2017)。由于森林生态系统结构复杂且植株个体生长周期长,因此开展BEF研究相对困难。森林BEF实验较之草地系统有其突出的优势:更加方便在个体水平上开展实验研究,控制密度和均匀度;更加充分地观察到物种间及其与环境间的相互作用随时间的变化 (马克平, 2013)。
森林BEF的研究起步较晚,自2000年以来,在全球范围内陆续建立了29个森林BEF的实验,形成了全球森林BEF研究网络 (TreeDivNet, www.treedivnet.ugent.be),研究区域包括寒温带、温带、亚热带和地中海地区,树种多样性水平设置为0~24种不等,涉及230个树种,百万余棵树木 (Paquette et al., 2018)。这些实验的具体设计虽然各不相同,但都是通过控制木本植物的物种多样性来量化树种多样性与生态系统功能的关系。在这些森林BEF实验中,亚热带森林生物多样性与生态系统功能实验基地 (Biodiversity-Ecosystem Functioning Experiment China Platform, BEF-China) 是亚热带首个人工控制生物多样性实验,通过开展长期的系统性研究,探究亚热带森林植物、动物、微生物等多营养级的生物多样性对生态系统功能和稳定性的影响及其内在机制。
1实验样地
BEF-China实验样地包括两个部分,即位于江西省德兴市新岗山镇的人工控制实验样地和钱江源国家公园内的比较实验样地 (图1)。
1.1 人工控制实验样地
该样地包含样地A和样地B。样地A于2009年建成,海拔105~275 m;样地B于2010年建成,海拔105~190 m(图2)。两个样地的物种库共包含42种乔木和18种灌木,按照随机断棍和物种直接丧失两种方式进行多样性设计。两个样地均以666.67 m2 (25.82 m × 25.82 m, 1亩) 为基本单元样方进行幼苗种植,共计566个样方。在一个基本样方中,乔木物种水平分别为1、2、4、8、16、24种。灌木配植在由4个基本单元样方组成的超级样方中,多样性水平分别为0、2、4、8种。在实验样地的每个基本单元样方内栽植400棵乔木幼苗,间距为1.29 m,灌木设置同样的密度,栽植在乔木之间。两个样地栽植的木本植物总数超过30万株 (马克平, 2013; Bruelheide et al., 2014)。
1.2 比较实验样地
该样地于2008年建立,共设27个30 m × 30 m的小样地。样地覆盖5个演替阶段,当年的群落年龄分别为<20、20~40、40~60、60~80、>80年。根据第一次普查的结果,株高大于1 m的木本植物有148种,隶属于46科 (Bruelheide et al., 2011)。
2新岗山主实验样地研究进展
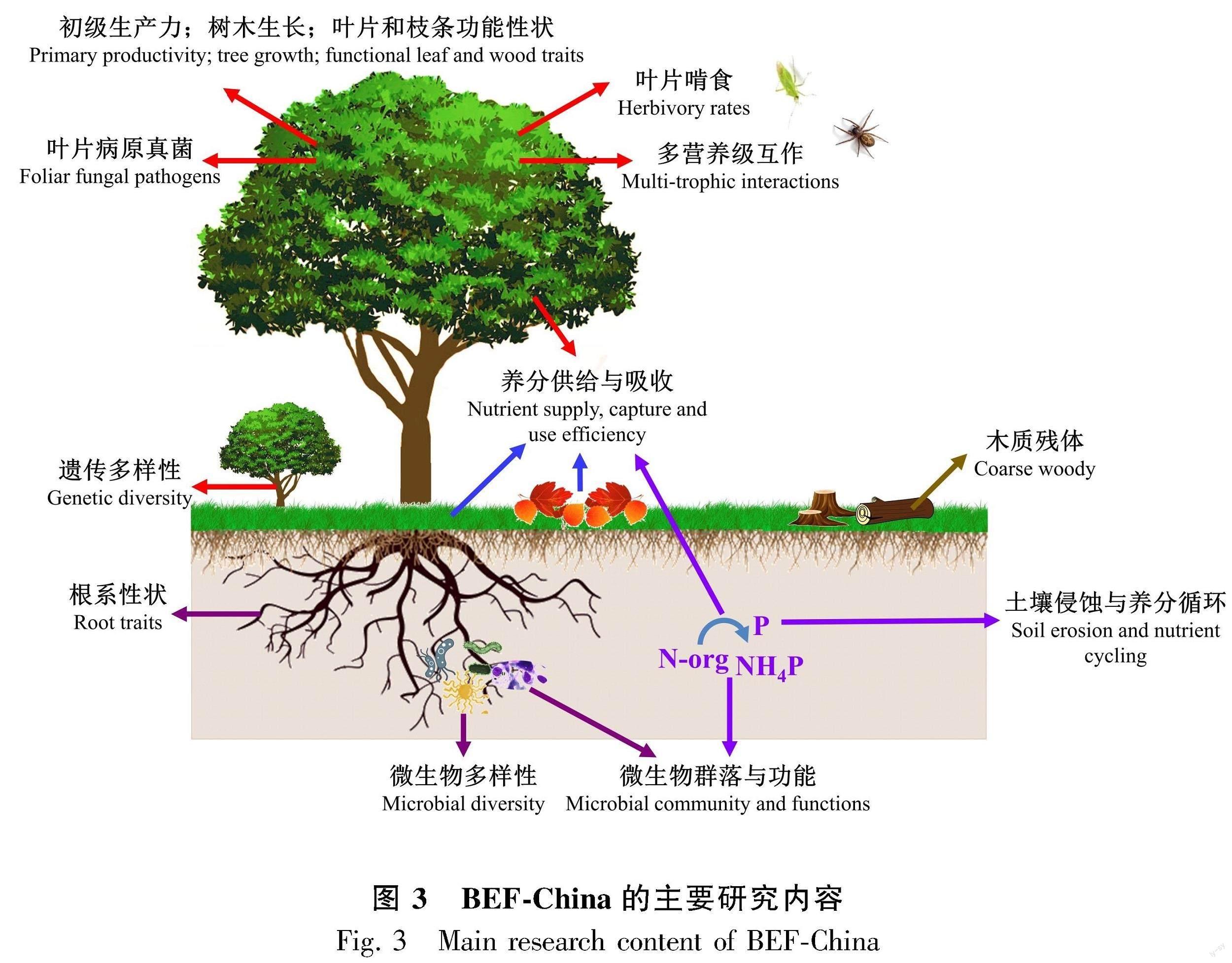
截至2023年3月,基于BEF-China平台已发表论文229篇,内容涉及生物多样性对多种生态系统功能的影响,包括生产力、养分循环、多营养级相互作用关系等方面 (图3)。
2.1 树种多样性对树木生长和生产力的影响
在实验的早期阶段,通过对幼树生长速率(Lang et al., 2012; Li et al., 2014; Hahn et al., 2017b)、冠层结构(Lang et al., 2012)、吸水特性 (Trogisch et al., 2016)等方面的研究表明,树种多样性对生产力的促进作用因年限不足而未显现。幼树生长表现出明显的物种特异性,受物种本身功能性状和所处环境的影响,树木生长和性状关系可以随着树种多样性的变化而改变,在树种多样性较高的群落,性状对生长的影响更为显著( Li et al., 2014; Krber et al., 2015;Bongers et al., 2020b)。
随着时间的推移,BEF-China平台陆续报道了关于树种多样性对生产力的促进作用及其作用机制的研究。种植8年后,每公顷16个物种混交林的碳储量约32 t,而每公顷纯林的碳储量约为12 t,不及混交林一半。所有多样性水平群落的生物多样性效应随时间的推移而增加,并主要由互补效应驱动(Huang et al., 2018a)。从物种丧失的角度来研究,Chen等(2020)通過模拟BEF-China平台的随机和非随机灭绝情景,发现物种丧失在所有情景下都对生产力产生损害,并且这些影响随着林龄的增长而增强。
土壤中的某些真菌与植物的根系形成共生体,即菌根。丛枝菌根 (AM) 和外生菌根(ECM) 是森林中常见的菌根类型,菌根类型在很大程度上影响了树种多样性对生产力的影响 (Ma et al., 2021)。随着树种多样性的增加,AM树种的净初级生产力增加,而ECM树种则减少,这主要是因为随着树种多样性的增加,AM树种增强了养分吸收和加快了凋落物分解,而ECM树种的凋落物分解和养分吸收之间存在权衡。这些结果证实了AM树种通过使用不同于ECM树种的养分获取策略,在亚热带森林中占据主导地位 (Deng et al., 2023)。
邻体间相互作用解释了群落生产力沿多样性梯度50%以上的变异,并随着群落树种多样性的增加,局部尺度邻居效应对调节群落生产力的重要性明显增加(Fichtner et al., 2018)。树冠的互补性和可塑性随着树种多样性的增加而增大,进一步促进了光吸收和树木生长(Forrester et al., 2019; Perles-Garcia et al., 2021)。随着时间的推移,邻居树种多样性导致木材体积分配发生变化,更有利于枝条,尤其是形态灵活的树种生长。树种多样性介导的空间分配模式和树冠形态的变化是树冠互补的基本机制,也是群落超产的重要驱动因素 (Kunz et al., 2019)。
另外,树种多样性能够显著促进群落生产力的稳定性,其内在的机制在于树种多样性促进了与导水、气孔控制相关的功能多样性,从而导致树种具有不同的异步性生长,最终决定了群落稳定性这一关键机制(Schnabel et al., 2021)。
树种多样性通过影响物种水平的功能性状,包括树冠 (Perles-Garcia et al., 2022)、叶片、根系等方面的功能性状(Weinhold et al., 2022),从而显著影响树木生长 (Li et al., 2017b)。这种物种水平的影响,能否累积到群落水平上,并且如何受到树种多样性调控?基于BEF-China连续10年的监测数据分析发现,随着森林的发展,功能多样性和功能均值的影响强度都有增强,功能多样性的重要性在上升,而功能均值则在下降。这意味着随着时间的推移,群落中拥有更多样化的树种,对生产力提高的重要性和影响强度都在加强 (Bongers et al., 2021)。
森林是由树木个体组成,个体水平的相互作用往往在森林BEF研究中被忽略。来自不同母树个体形成的遗传多样性对生产力的影响如何?Bongers等(2020b) 对具有明确母树来源的树木个体进行了连续8年的生长追踪,并测取了关键的叶性状和枝条性状。结果发现,个体水平的功能性状对生长的影响在高多样性群落中作用更强,并且这种影响强度在单种和混种中差异随时间而增大。进一步将这些个体的遗传信息加入发现,遗传多样性促进了种内个体功能性状的分化,这些分化形成了对资源利用的功能空间差异,最终影响了群落生产力(Bongers et al., 2020a)。树种多样性和遗传多样性可以通过增加树木的功能多样性,抑制食草昆虫损伤和与土壤真菌的资源竞争,进而间接促进森林群落生产力。树木的遗传多样性在不同的树种多样性下对生产力的影响不同:在纯林中,没有发现种内遗传多样性会降低森林中土壤真菌多样性或食草昆虫的资源竞争压力;在混交林中,树木遗传多样性通过降低与土壤真菌竞争资源促进了群落生产力 (Tang et al., 2022)。
在地下生产力方面,Sun等 (2017) 的研究表明,细根产量随着树种多样性的增加而增大,这种影响能用树种间的地下生态位互补性来解释,是导致地上生物多样性超产效应的机制之一。
Bu等(2017)研究发现,树种多样性显著影响了5个树种的比根长,说明资源分配和物种相互作用是亚热带森林树种多样性对树木生长影响的潜在机制。
2.2 树种多样性对存活率的影响
2.2.1 乔木存活率根据2009年11月和2010年6月对样地A中26个乔木树种的两次调查统计,栽植14个月后的树苗成活率为87%,其中常绿树种成活率为84%,落叶树种成活率为93%。幼苗的存活率受树种多样性、叶的特性 (落叶或常绿)、树种功能特性、种植时间和海拔高度的显著影响(Yang et al., 2013)。Liu 等 (2022b)分析了第3至第12年的個体存活数据集发现,尽管树木的总体存活率随样方树种多样性的增加而增高,但树种多样性对存活率的影响在物种间和年际间存在显著差异。这些显著差异与物种的功能性状和当年的气候条件密切相关。保守型树种的存活率随树种多样性、林龄和年降水量的增加而提高,而获取型物种则相反。这表明树种多样性、 功能性状和气候条件之间的相互作用可以平衡森林中不同树种的存活率。
2.2.2 灌木存活率在实验的早期阶段,乔木和灌木的多样性都不影响灌木的存活。非生物环境因素对灌木存活的影响最大。干旱是灌木死亡的主要原因。在生物因子中,只有草本层树种丰富度和优势蕨类物种 [双鳞毛蕨(Dryopteris sp.)] 的盖度才能影响灌木的存活(Yang et al., 2017)。薛玉洁等 (2023)基于2012 年和 2019 年的灌木个体存活数据发现,不同物种的灌木存活率具有显著差异。地形和土壤碳氮比对灌木存活率影响较大,乔木和灌木多样性对灌木存活影响有限,但乔木多样性的增加可通过形成更郁闭的林冠提高灌木存活率,灌木功能性状显著影响灌木存活率,并且保守型生活策略的灌木物种具有更高的存活率。
2.3 树种多样性对凋落物分解和养分循环的影响
基于BEF-China的研究表明,树种多样性加快了凋落物的分解 (Deng et al., 2023),分别是通过影响凋落物分解过程中组分变化(Ristok et al., 2017)、胞外酶活性 (潘玉梅和张乃莉, 2021)和土壤微生物群落组成及活性(Pei et al., 2017) 来实现的。凋落物物种多样性对凋落物分解的影响在很大程度上取决于混合凋落物的物种组成 (Huang et al., 2018b)。遗传多样性对凋落物的分解有着非常重要的影响:分解速率随个体遗传多样性的增加而增大 (Li et al., 2017a)。土壤大型动物和叶的功能特征能够驱动凋落物次生代谢产物的分解(Ristok et al., 2019)。除了生物多样性的作用之外,地形和微气候等非生物因素也对凋落物的分解速率有着至关重要的作用 (Seidelmann et al., 2016)。在枯木分解方面,节肢动物分解者群落 (尤其是白蚁的存在) 是最主要的影响因子,而树种多样性本身并没有直接的影响 (Eichenberg et al., 2017)。树冠密度的增加延缓细木分解,主要是通过促进捕食性蚂蚁的数量和形成更潮湿凉爽的小气候,从而抑制白蚁觅食细木的强度(Wu et al., 2021a,b)。
树种多样性促进了土壤有机碳在不同土壤深度的积累,凋落物输入是重要的影响因素(Li et al., 2019)。另外,土壤碳含量随着树木生产力和根系直径的增加而增加,并随着凋落物碳氮比的增加而减少。此外,树木生产力和树木功能性状 (如菌根类型和凋落物碳氮比) 调节了微环境条件,对土壤微生物生物量产生了较大影响,进而影响土壤碳含量 (Beugnon et al., 2023)。
树种多样性促进氮素的获取和存留,并随着时间的推移而增强,氮吸收方面的生态位互补促进了共存物种之间的资源利用(Lang et al., 2014)。Liu等(2022a)的盆栽实验研究同样证明,随着树种多样性的增加,生态系统生物量的增加在很大程度上可以通过树种之间氮吸收的生态位互补来解释。
2.4 树种多样性对多营养级相互作用关系的影响
树种多样性对多营养级相互作用关系的影响在蜜腺植物访问、食草动物啃食、调节高营养级之间关系和微生物群落等方面均有报道。蜜腺访问者的数量和物种多样性随着树种多样性的增加而下降,这体现了资源稀释效应,由于树种越多的样方,蜜腺植物的个体比例越小,因此花蜜可用性越低 (Staab et al., 2017)。在食草动物啃食方面,多篇报道指出,树种多样性促进食草动物啃食程度的增加(Schuldt et al., 2017c; Yang et al., 2018)。这是广食性的食草动物 (特别是成年食草动物)能啃食更多样的树木造成的(Zhang et al., 2018)。树种多样性对半翅目昆虫和蚂蚁的物种多样性呈现促进作用,加强了植物-蚂蚁的互惠作用 (Staab et al., 2015),减弱了共生蚂蚁-半翅目昆虫相互作用与食草动物之间的相关关系(Schuldt et al., 2017b)。树木系统发育多样性而不是树种多样性决定了不同营养级的节肢动物群落组成,树木系统发育多样性增加了捕食性节肢动物的多样性,但降低了草食性节肢动物多样性 (Staab et al., 2021)。总体而言,树种多样性促进更高营养级的物种丰富度,通过提高植物群落的结构和功能多样性来促进高营养级物种之间的相互作用 (拮抗作用和互惠作用),从而增强了森林生态系统应对环境条件变化的稳定性(Cao et al., 2018; Fornoff et al., 2019; Schuldt et al., 2019; Albert et al., 2022)。
基于BEF-China样地的研究,有报道指出,树种多样性对高营养级的作用并不显著。例如,树种多样性对鳞翅目昆虫多样性的影响在很大程度上是间接的,是通过改变鳞翅目昆虫的多样性来实现的,进化依赖性决定了食草动物群落对树种多样性变化的反应 (Wang et al., 2020)。植食性鳞翅目幼虫共现指数与树木和鳞翅目的系统发生距离(mean pairwise phylogenetic distance, MPD)呈显著负相关,与营养功能性状 (碳氮比) 呈正相关,但与防御性状 (如叶韧度),以及树种多样性 (包括树的物种丰富度和MPD) 呈负相关。Wang等(2022)研究表明,在亚热带森林生态系统中,以环境过滤为主的多种驱动力能通过物种间的相互作用,显著影响植食者群落的物种共现共存。
微生物方面,树木菌根类型、树种多样性及其相互作用对微生物群落组成有显著影响(Yang et al., 2022)。树木菌根类型是决定土壤微生物多样性和群落组成的重要因素,较高的树种多样性促进了土壤微生物群落组成的趋同(Singavarapu et al., 2022)。樹种多样性是通过增加微生物生物量而不是改变微生物种类或功能多样性来增加土壤微生物呼吸 (Beugnon et al., 2021)。真菌共生网络结构受树种多样性、群落组成以及土壤碳氮比的影响,但细菌共生网络结构受到土壤pH值和空间距离的影响 (Gan et al., 2022)。树种多样性增加促进了叶片真菌病原体的多样性 (Rutten et al., 2021),并抑制了叶片真菌的感染(Saadani et al., 2021),这主要是由宿主稀释效应引起的。在更多样化的植物群落中,个体的平均易感率降低,宿主空间距离的增加可能会减少病原体的传播,从而减少病原体在更多样的植物群落中的影响。病原菌危害减少,尤其对叶片质地柔软、气候生态位狭窄的树种影响最为显著(Schuldt et al., 2017c)。
2.5 树种多样性对生态系统功能其他方面的影响
树种多样性对生态系统功能的影响体现在抵抗干旱胁迫(Salmon et al., 2018; Fichtner et al., 2020)、林冠穿透雨(Geissler et al., 2012; Goebes et al., 2015b)、抵抗土壤侵蚀(Song et al., 2019) 和物候变化 (Du et al., 2019)等方面。树种多样性能在一定程度上缓解幼树生长的干旱胁迫。树种多样性的促进作用在干旱期间最强,并随着树种多样性的增加而增加。干旱期间,土壤水分分配有利于最脆弱的个体 (Fichtner et al., 2020)。早期林冠穿透雨实验的研究表明,在样方水平上,是邻居树种多样性而不是群落树种多样性对林冠穿透雨产生较大的影响,说明幼林中树种多样性只在邻体水平的小空间上产生影响(Goebes et al., 2015a)。随着森林的发展,树种多样性的作用逐渐增强,在多样性高的样方中,较高的树冠盖度和叶面积指数降低了森林生态系统的林间侵蚀(Seitz et al., 2016; Song et al., 2018)。另外,石头、生物结皮、凋落物对土壤表面的覆盖也是控制土壤侵蚀的重要因素(Seitz et al., 2015; Seitz et al., 2016; Song et al., 2019),凋落物的保护作用随着凋落物的分解而消退,保护效果受土壤中大型动物存在与否的影响 (Seitz et al., 2015)。在物候方面,树种多样性的波动可以改变亚热带森林的展叶时间,非生物变量比生物变量对展叶时间的影响更为重要。总体而言,树种多样性的下降可能会加剧全球气温上升引起的物候变化 (Du et al., 2019)。
3比较实验样地研究进展
3.1 不同演替阶段树种多样性对生产力的影响
植物生长速率在季节和年度间存在显著差异,虽然随林龄的增加而降低,但与地形和邻居树种多样性或密度无显著相关(Chi et al., 2017)。在碳储量方面,树种多样性会促进群落碳的累积,即树种多样性高的林分具有更高的碳储量和碳通量。林龄对碳储量的影响,表现为老龄林的碳储量高于中龄林和幼龄林。在研究区域每增加一个树种,森林群落的总碳储量增加了6.4%(Liu et al., 2018)。在草本层方面,草本层生产力既不受乔木多样性的影响,也不受草本多样性的影响。尽管在演替过程中草本层物种的多样性显著降低,但在演替的各个阶段树种多样性对草本层物种多样性的贡献都非常大,而环境因素的影响并不大(Both et al., 2011)。
3.2 不同演替阶段树种多样性对凋落物分解和养分循环的影响
凋落物总量随树种多样性的增加而增大,老龄林具有更多的凋落物。增加树种多样性可提高凋落物质量 (较低的碳氮比),而林龄对凋落物质量无影响。凋落物中的氮含量越高,养分循环速度更快,进一步促进了森林生长(Huang et al., 2017)。Eichenberg 等(2015)研究发现,凋落物分解不是随林龄发生显著变化,而是主要受到叶片功能性状的影响。在木质残体分解方面,Pietsch等(2019) 研究发现,亚热带森林木材分解主要受温度的控制,树种多样性、真菌操作分类学单元 (OTUs) 多样性和大型无脊椎动物多样性对木材分解速率的影响相对较弱。
众所周知,树种多样性会影响土壤有机碳的存储,而驱动二者作用关系的机制尚不清楚。树种多样性和土壤有机碳关系在表层土和底土中有着不同的驱动途径。在表层(0~10 cm)土壤中,树种多样性通过调节植物衍生成分 (木质素酚类、轻组有机碳和颗粒有机物) 影响土壤有机碳含量 (Jia et al., 2021)。在底层(30~40 cm)土壤中,树种多样性对微生物衍生成分 (氨基糖类和矿物结合有机质) 的积累有促进作用,从而在底层土的土壤有机碳含量变化中占主导地位 (Jia et al., 2021)。进一步地,根际土壤有机碳的来源及其调控机制也不清楚。与非根际土壤有机碳相比,木质素酚类物质在根际土壤有机碳中的浓度更高,由于ECM真菌的贡献,因此氨基糖更集中在ECM树的根际。但是,在AM树下之所以不集中,是因为AM树表现出降低的真菌坏死体积累。该结果突出了与不同菌根树种有关的根际土壤有机碳来源差异的新机制,对于利用植物-菌根共生增强土壤有机碳固存具有重要意义 (Jia et al., 2022)。
3.3 不同演替阶段树种多样性对多营养级互作的影响
随着树种多样性和系统发育多样性的增加,草食性节肢动物的数量增加,所造成的叶片啃食程度增加。冠层中叶生物量比例较低的树种往往遭受更高程度的啃食伤害。广食性的食草动物在树种多样性较高的森林系统中受益最大 (Brezzi et al., 2017; Schuldt et al., 2010)。木本植物的功能多样性 (叶片的化学组分) 和系统发育多样性驱动着高度多样性森林中的啃食伤害情况(Schuldt et al., 2014a)。
幼龄林和老龄林中的蚂蚁群落组成存在显著差异,幼龄林中蚂蚁群落具有更高的物种多样性,而在老龄林中存在一个独特的蚂蚁群落(Staab et al., 2014a)。杂食性蚂蚁均匀度随树木均匀度的增加而降低,随林龄的增加而增大(Staab et al., 2014b)。捕食性蚂蚁的物种多样性随树种多样性的增加而增大,随叶片的功能多样性和灌木盖度的增加而降低。海拔高度对蚂蚁物种多样性有负的影响(Staab et al., 2014a)。
随着树种多样性和林龄的增加,蜘蛛群落功能的差异增大。蜘蛛的均匀度和分化度随着树种多样性和林龄的增加而增加。尽管蜘蛛物种多样性随着树种多样性的增加而降低(Schuldt et al., 2011),但功能多样性仍保持在恒定水平。这表明蜘蛛的功能冗余度随着树种多样性的增加而降低 (Schuldt et al., 2013b)。另外,环境异质性 (Schuldt et al., 2012)和尺度差异 (Schuldt et al., 2013a)也是影响蜘蛛群落的重要因素。
总体而言,亚热带森林系统中的多年生群落的地下生物群落是通过树种多样性的下行控制效应来构建的,而地上节肢动物群落是通过树种多样性的上行控制效应来构建的(Schuldt et al., 2017a)。在高度多样化的森林中,树种多样性加强了蚂蚁与蜘蛛群落组成和功能之间的关系,即在蚂蚁存在的情况下,蜘蛛家族的多样性增加,造网蜘蛛和猎网蜘蛛的生物量比例发生了变化,形成了更多以造网功能为主的蜘蛛群落 (Schuldt & Staab 2015)。除了树种多样性的影响之外,植物系统发育多样性、植物功能多样性、多营养级多样性的重要作用也有报道。植物系统发育多样性对草食性动物群落的上行控制作用强于树种多样性的作用(Schuldt et al., 2014b)。树木系统发育多样性促进寄主-寄生蜂相互作用,并且比树种多样性的效应强得多 (Staab et al., 2016)。植物功能性状多样性和组成比树种多样性对个体功能和多功能性的影响更大,多营养级多样性是理解多功能驱动因素的关键(Schuldt et al., 2018)。除了生物多样性的影响之外,海拔的作用也不容忽视,即使在相对较小的梯度上,海拔对动物群落的物种多样性也有很强的直接作用,但其影响取决于当地资源的可用性和高营养级生物自身的特征(Binkenstein et al., 2018)。
在微生物方面,Gao等(2015) 利用454高通量測序技术对幼龄林、中龄林和老龄林中的外生菌根真菌群落进行了研究,发现外生菌根真菌群落由393个OTUs组成,隶属于21个外生菌根真菌谱系,其中3个外生菌根真菌谱系和11个外生菌根真菌OTUs在幼龄林、中龄林和老龄林中表现出明显的偏向性。宿主植物和真菌群落之间存在显著的相关性。亚热带森林植物群落组成是土壤真菌多样性和群落组成的主要驱动因子 (Wu et al., 2012)。Gao等(2013)研究发现,在属水平上,亚热带森林树种多样性是其外生菌根真菌多样性的最主要影响因素;共生植物的组成决定了外生菌根真菌特性及多样性对亚热带树木竞争的影响 (Shi et al., 2017)。另外,Wu等(2013) 发现,海拔、土壤有机碳、土壤pH值也是影响真菌群落组成的重要因素。
4BEF-China研究展望
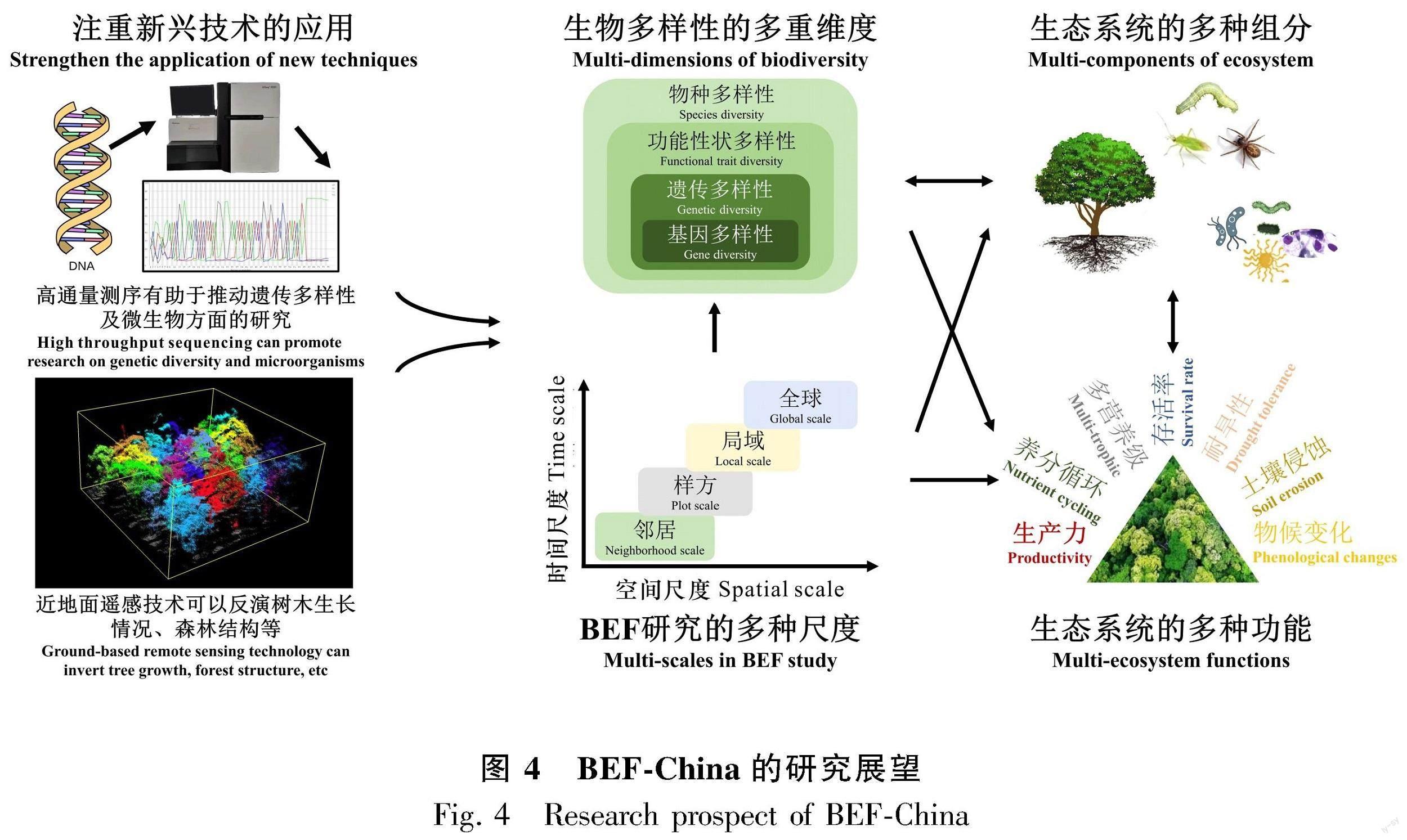
基于BEF-China研究进展的汇总,我们提出未来BEF-China的研究可以在多个生物多样性维度、多种生态系统组分、多种生态系统功能以及多种时空尺度等方面进行综合考虑 (图4)。
4.1 生物多样性维度方面
目前BEF-China的研究对树种多样性和功能多样性的考虑较多,但由于受研究专业、技术手段和研究经费的限制,因此对遗传多样性的涉猎并不多。已涉及的方面有乔木遗传多样性(Schuldt et al., 2014a, b; Staab et al., 2016; Hahn et al., 2017a; Purschke et al., 2017; Tang et al., 2022)和食草动物遗传多样性 (Wang et al., 2020)。遗传多样性既是生物体内的遗传变异性,也是物种拥有的遗传物质和遗传资源的总和,主要指在分子水平上的,特别是基因和基因组水平上的多样性,包括由遗传物质决定的形态遗传性状变异和适应性性状变异。Forest等(2007)研究指出,一些地区虽然物种多样性并不占优势,但却包括了一些在系统发育树上隔离孑遗的古老分支,这些堪称活化石的物种包含了更独特和稀有的进化历史信息,这些信息保留在其基因和基因组中。未来BEF-China的研究,结合多种多样性维度,从树种多样性、功能多样性、遗传多样性等视角来分析BEF的关系,以期得到更深层次的理解。
4.2 生态系统组分
目前,BEF-China的研究对植物、动物的探讨很多,而对微生物群落的研究报道并不多,背后的原因和遗传多样性的研究相对不多是一样的。生物多样性与地上-地下生态学过程研究主要涉及凋落物分解和根系吸收这两个方面,而土壤微生物 (真菌和病原菌) 的组成和多样性通过参与凋落物分解、形成共生菌根、产生病菌危害等形式对上述两个过程发挥着重要作用,进而影响地上生态系统多营养级的相互作用关系(Delgado-Baquerizo et al., 2016; Chen et al., 2019 )。
高通量测序技术的发展能对遗传多样性和微生物群落方面的研究提供有利的帮助。随着测序费用的降低以及DNA序列片段长度和质量的提升,过去只有大型研究机构才能开展的测序工作如今也可在普通的实验室进行(Schneider et al., 2010)。技术进步将会带来更多的方法,推动BEF-China研究在微生物群落、系统进化等方面有更好的发展。
4.3 生态系统功能
目前,大部分基于BEF-China的研究都只考虑了生物多样性对单一生态过程的影响,建立能够合理解释生态系统各功能综合响应的多功能性指标,是摆在BEF研究者面前的一个挑战。卫星遥感可用于估计整个研究区域土地覆被的种类、类型和范围,提供大量研究区域的长期生物物理和生境条件,遥感数据结合实地资源、土地覆被分类,可以确定非常具体的生境,并进一步用于栖息地建模、物种分布预测、物种生境丧失,检测模型灭绝率的变化(Parmesan & Yohe, 2003)。基于实地的生态系统功能度量不容易转化为对整个生态系统功能的估计,而通过遥感手段可以同时估计整个生态系统的功能 (Rocchini et al., 2004)。
4.4 时空尺度方面
一般来讲,在小到中等空间尺度水平内,生物多样性的相对作用是很大的。例如,小到邻居树木尺度上,邻居树种多样性影响自身性状的变异,进而影响了邻居物种与目标物种的相互作用 (Lankau, 2011);邻居树种多样性既能改变邻居物种与目标物种的相对竞争强度,也能通过生态位互补增加种群的竞争强度 (Dostal,2011)。但是,在区域尺度内,环境的异质性较大,生物因子作为生态系统过程的预测因子的重要性将被削弱,非生物因子成为生态系统对环境梯度变化响应的主要驱动力。因此,正确理解和预测大尺度水平上生物多样性和生态系统过程所发生的变化,如当考虑BEF-China与别的国家、地区的BEF森林研究成果进行综合比较分析时,就必须强调生物多样性的变化、生态系统功能和环境因子之间的反馈效应 (Loreau et al., 2001)。
4.5 時间尺度方面
BEF-China目前最主要的研究成果之一,是植物生产力随树种多样性增加而增加,并且这种增加在逐年增大(Huang et al., 2018a)。那么这种增加何时能达到稳定?在演替过程中,生态系统内的各种组分、生态学过程和功能又发生了怎样的变化?对伴随发生的全球气候变化产生了怎样的反应?这些问题也许都将在BEF-China未来的研究报道中得到解答。
参考文献:
ALBERT G, GAUZENS B, LOREAU M, et al., 2022. The hidden role of multi-trophic interactions in driving diversity-productivity relationships [J]. Ecol Lett, 25(2): 405-415.
BEUGNON R, BU W, BRUELHEIDE H, et al., 2023. Abiotic and biotic drivers of tree trait effects on soil microbial biomass and soil carbon concentration [J]. Ecol Monogr, 93(2): e1563.
BEUGNON R, DU J, CESARZ S, et al., 2021. Tree diversity and soil chemical properties drive the linkages between soil microbial community and ecosystem functioning [J]. ISME Commun, 1: 41.
BINKENSTEIN J, KLEIN AM, ASSMANN T, et al., 2018. Multi-trophic guilds respond differently to changing elevation in a subtropical forest [J]. Ecography, 41(6): 1013-1023.
BONGERS FJ, SCHMID B, BRUELHEIDE H, et al., 2021. Functional diversity effects on productivity increase with age in a forest biodiversity experiment [J]. Nat Ecol Evol, 5: 1594-1603.
BONGERS FJ, SCHMID B, DURKA W, et al., 2020a. Genetic richness affects trait variation but not community productivity in a tree diversity experiment [J]. New Phytol, 227(3): 744-756.
BONGERS FJ, SCHMID B, SUN Z, et al., 2020b. Growth-trait relationships in subtropical forest are stronger at higher diversity [J]. J Ecol, 108(1): 256-266.
BOTH S, FANG T, BOHNKE M, et al., 2011. Lack of tree layer control on herb layer characteristics in a subtropical forest, China [J]. J Veg Sci, 22(6): 1120-1131.
BREZZI M, SCHMID B, NIKLAUS PA, et al., 2017. Tree diversity increases levels of herbivore damage in a subtropical forest canopy: evidence for dietary mixing by arthropods? [J]. J Plant Ecol, 10(1): 13-27.
BRUELHEIDE H, BOHNKE M, BOTH S, et al., 2011. Community assembly during secondary forest succession in a Chinese subtropical forest [J]. Ecol Monogr, 81(1): 25-41.
BRUELHEIDE H, NADROWSKI K, ASSMANN T, et al., 2014. Designing forest biodiversity experiments: general considerations illustrated by a new large experiment in subtropical China [J]. Methods Ecol Evol, 5(1): 74-89.
BU WS, SCHMID B, LIU XJ, et al., 2017. Interspecific and intraspecific variation in specific root length drives aboveground biodiversity effects in young experimental forest stands [J]. J Plant Ecol, 10(1): 158-169.
CAO HX, KLEIN A-M, ZHU C, et al., 2018. Intra- and inter-specific tree diversity promotes multitrophic plant-Hemiptera-ant interactions in a forest diversity experiment [J]. Basic Appl Ecol, 29: 89-97.
CARDINALE BJ, DUFFY JE, GONZALEZ A, et al., 2012. Biodiversity loss and its impact on humanity [J]. Nature, 486(7401): 59-67.
CHEN L, SWENSON NG, JI NN, et al., 2019. Differential soil fungus accumulation and density dependence of trees in a subtropical forest [J]. Science, 366(6461): 124-128
CHEN Y, HUANG Y, NIKLAUS PA, et al., 2020. Directed species loss reduces community productivity in a subtropical forest biodiversity experiment [J]. Nat Ecol Evol, 4(4): 550-559.
CHI XL, GUO Q, FANG JY, et al., 2017. Seasonal characteristics and determinants of tree growth in a Chinese subtropical forest [J]. J Plant Ecol, 10(1): 4-12.
DELGADO-BAQUERIZO M, MAESTRE FT, REICH PB, et al., 2016. Microbial diversity drives multifunctionality in terrestrial ecosystems [J]. Nat Commun, 7: 10541.
DENG M, HU S, GUO L, et al., 2023. Tree mycorrhizal association types control biodiversity-productivity relationship in a subtropical forest [J]. Sci Adv, 9(3): eadd4468.
DOSTAL P, 2011. Plant competitive interactions and invasiveness: searching for the effects of phylogenetic relatedness and origin on competition intensity [J]. Am Nat, 177(5): 655-667.
DU YJ, YANG B, CHEN SC, et al., 2019. Diverging shifts in spring phenology in response to biodiversity loss in a subtropical forest [J]. J Veg Sci, 30(6): 1175-1183.
EICHENBERG D, PIETSCH K, MEISTER C, et al., 2017. The effect of microclimate on wood decay is indirectly altered by tree species diversity in a litterbag study [J]. J Plant Ecol, 10(1): 170-178.
EICHENBERG D, TROGISCH S, HUANG YY, et al., 2015. Shifts in community leaf functional traits are related to litter decomposition along a secondary forest succession series in subtropical China [J]. J Plant Ecol, 8(4): 401-410.
FICHTNER A, HARDTLE W, BRUELHEIDE H, et al., 2018. Neighbourhood interactions drive overyielding in mixed-species tree communities [J]. Nat Commun, 9: 1144.
FICHTNER A, SCHNABEL F, BRUELHEIDE H, et al., 2020. Neighbourhood diversity mitigates drought impacts on tree growth [J]. J Ecol, 108(3): 865-875.
FOREST F, GRENYER R, ROUGET M, et al., 2007. Preserving the evolutionary potential of floras in biodiversity hotspots [J]. Nature, 445(7129): 757-760.
FORNOFF F, KLEIN AM, BLUTHGEN N, et al., 2019. Tree diversity increases robustness of multi-trophic interactions [J]. P Roy Soc B-Biol Sci, 286(1898): 20182399.
FORRESTER D, RODENFELS P, HAASE J, et al., 2019. Tree-species interactions increase light absorption and growth in Chinese subtropical mixed-species plantations [J]. Oecologia, 191(2): 421-432.
GAN H, LI X, WANG Y, et al., 2022. Plants play stronger effects on soil fungal than bacterial communities and co-occurrence network structures in a subtropical tree diversity experiment [J]. Microbiol Spectrum, 10(3): e0013422.
GAO C, SHI NN, LIU YX, et al., 2013. Host plant genus-level diversity is the best predictor of ectomycorrhizal fungal diversity in a Chinese subtropical forest [J]. Mol Ecol, 22(12): 3403-3414.
GAO C, ZHANG Y, SHI NN, et al., 2015. Community assembly of ectomycorrhizal fungi along a subtropical secondary forest succession [J]. New Phytol, 205(2): 771-785.
GEISSLER C, KHN P, BHNKE M, et al., 2012. Splash erosion potential under tree canopies in subtropical SE China [J]. Catena, 91: 85-93.
GOEBES P, BRUELHEIDE H, HARDTLE W, et al., 2015a. Species-specific effects on throughfall kinetic energy in subtropical forest plantations are related to leaf traits and tree architecture [J]. PLoS ONE, 10(6): e128084.
GOEBES P, SEITZ S, KUEHN P, et al., 2015b. Throughfall kinetic energy in young subtropical forests: Investigation on tree species richness effects and spatial variability [J]. Agric For Meteorol, 213: 148-159.
HAHN CZ, MICHALSKI SG, FISCHER M, et al., 2017a. Genetic diversity and differentiation follow secondary succession in a multi-species study on woody plants from subtropical China [J]. J Plant Ecol, 10(1): 213-221.
HAHN CZ, NIKLAUS PA, BRUELHEIDE H, et al., 2017b. Opposing intraspecific vs. interspecific diversity effects on herbivory and growth in subtropical experimental tree assemblages [J]. J Plant Ecol, 10(1): 242-251.
HUANG YY, CHEN YX, CASTRO-IZAGUIRRE N, et al., 2018a. Impacts of species richness on productivity in a large-scale subtropical forest experiment [J]. Science, 362(6410): 80-83.
HUANG YY, MA YL, ZHAO K, et al., 2017. Positive effects of tree species diversity on litterfall quantity and quality along a secondary successional chronosequence in a subtropical forest [J]. J Plant Ecol, 10(1): 28-35.
HUANG YY, MA KP, NIKLAUS PA, et al., 2018b. Leaf-litter overyielding in a forest biodiversity experiment in subtropical China [J]. For Ecosyst, 5: 38.
JIA Y, LIU Z, ZHOU L, et al., 2022. Soil organic carbon sourcing variance in the rhizosphere vs. non-rhizosphere of two mycorrhizal tree species [J]. Soil Biol Biochem, 176: 108884.
JIA YF, ZHAI GQ, ZHU SS, et al., 2021. Plant and microbial pathways driving plant diversity effects on soil carbon accumulation in subtropical forest [J]. Soil Biol Biochem, 161: 108375.
KRBER W, LI Y, HAERDTLE W, et al., 2015. Early subtropical forest growth is driven by community mean trait values and functional diversity rather than the abiotic environment [J]. Ecol Evol, 5(17): 3541-3556.
KUNZ M, FICHTNER A, HARDTLE W, et al., 2019. Neighbour species richness and local structural variability modulate aboveground allocation patterns and crown morphology of individual trees [J]. Ecol Lett, 22(12): 2130-2140.
LANG AC, HARDTLE W, BARUFFOL M, et al., 2012. Mechanisms promoting tree species co-existence: Experimental evidence with saplings of subtropical forest ecosystems of China [J]. J Veg Sci, 23(5): 837-846.
LANG AC, VON OHEIMB G, SCHERER-LORENZEN M, et al., 2014. Mixed afforestation of young subtropical trees promotes nitrogen acquisition and retention [J]. J Appl Ecol, 51(1): 224-233.
LANKAU RA, 2011. Rapid evolutionary change and the coexistence of species [J]. Ann Rev Ecol Evol Syst, 42(1): 335-354.
LI SS, TONG YW, WANG ZW, 2017a. Species and genetic diversity affect leaf litter decomposition in subtropical broadleaved forest in southern China [J]. J Plant Ecol, 10(1): 232-241.
LI Y, BRUEHEIDE H, SCHOLTEN T, et al., 2019. Early positive effects of tree species richness on soil organic carbon accumulation in a large-scale forest biodiversity experiment [J]. J Plant Ecol, 12(5): 882-893.
LI Y, HARDTLE W, BRUELHEIDE H, et al., 2014. Site and neighborhood effects on growth of tree saplings in subtropical plantations (China) [J]. For Ecol Manag, 327: 118-127.
LI Y, KRBER W, BRUELHEIDE H, et al., 2017b. Crown and leaf traits as predictors of subtropical tree sapling growth rates [J]. J Plant Ecol, 10(1): 136-145.
LIU M, XU XL, YANG B, et al., 2022a. Niche partitioning in nitrogen uptake among subtropical tree species enhances biomass production [J]. Sci Total Environ, 823: 153716.
LIU X, HUANG Y, CHEN L, et al., 2022b. Species richness, functional traits and climate interactively affect tree survival in a large forest biodiversity experiment [J]. J Ecol, 110(10): 2522-2531.
LIU XJ, TROGISCH S, HE JS, et al., 2018. Tree species richness increases ecosystem carbon storage in subtropical forests [J]. P Roy Soc B-Biol Sci, 285(1885): 20181240.
LOREAU M, NAEEM S, INCHAUSTI P, et al., 2001. Biodiversity and ecosystem functioning: Current knowledge and future challenges [J]. Science, 294(5543): 804-808.
MA KP, 2013. Studies on biodiversity and ecosystem function via manipulation experiments. [J]. Biodivers Sci, 21(3): 247-248. [馬克平, 2013.生物多样性与生态系统功能的实验研究 [J]. 生物多样性, 21(3): 247-248.]
MA KP, HE JS, BRUELHEIDE H, et al., 2017. Biodiversity-ecosystem functioning research in Chinese subtropical forests [J]. J Plant Ecol, 10(1): 1-3.
MA LW, BONGERS FJ, LI S, et al., 2021. Species identity and composition effects on community productivity in a subtropical forest [J]. Basic Appl Ecol, 55: 87-97.
PAN YM, ZHANG NL, 2021. Effects of tree diversity on enzyme activity in litter of a subtropical forest ecosystem [J]. Biodivers Sci, 29(11): 1447-1460.[潘玉梅, 张乃莉, 2021. 亚热带森林树种多样性对凋落叶分解胞外酶活性的影响 [J]. 生物多样性, 29(11): 1447-1460.]
PAQUETTE A, HECTOR A, CASTAGNEYROL B, et al., 2018. A million and more trees for science [J]. Nat Ecol Evol, 2(5): 763-766.
PARMESAN C, YOHE G, 2003. A globally coherent fingerprint of climate change impacts across natural systems [J]. Nature, 421: 37-42.
PEI ZQ, LEPPERT KN, EICHENBERG D, et al., 2017. Leaf litter diversity alters microbial activity, microbial abundances, and nutrient cycling in a subtropical forest ecosystem [J]. Biogeochemistry, 134(1/2): 163-181.
PERLES-GARCIA MD, KUNZ M, FICHTNER A, et al., 2022. Neighbourhood species richness reduces crown asymmetry of subtropical trees in sloping terrain [J]. Remote Sens-Basel, 14(6): 1441.
PERLES-GARCIA MD, KUNZ M, FICHTNER A, et al., 2021. Tree species richness promotes an early increase of stand structural complexity in young subtropical plantations [J]. J Appl Ecol, 58(10): 2305-2314.
PIETSCH KA, EICHENBERG D, NADROWSKI K, et al., 2019. Wood decomposition is more strongly controlled by temperature than by tree species and decomposer diversity in highly species rich subtropical forests [J]. Oikos, 128(5): 701-715.
PURSCHKE O, MICHALSKI SG, BRUELHEIDE H, et al., 2017. Phylogenetic turnover during subtropical forest succession across environmental and phylogenetic scales [J]. Ecol Evol, 7(24): 11079-11091.
RISTOK C, LEPPERT KN, FRANKE K, et al., 2017. Leaf litter diversity positively affects the decomposition of plant polyphenols [J]. Plant Soil, 419(1/2): 305-317.
RISTOK C, LEPPERT KN, SCHERER-LORENZEN M, et al., 2019. Soil macrofauna and leaf functional traits drive the decomposition of secondary metabolites in leaf litter [J]. Soil Biol Biochem, 135: 429-437.
ROCCHINI D, CHIARUCCI A, LOISELLE SA, 2004. Testing the spectral variation hypothesis by using satellite multispectral images [J]. Acta Oecologica, 26: 117-120.
RUTTEN G, HNIG L, SCHWA R, et al., 2021. More diverse tree communities promote foliar fungal pathogen diversity, but decrease infestation rates per tree species, in a subtropical biodiversity experiment [J]. J Ecol, 109(5): 2068-2080.
SAADANI M, HNIG L, BIEN S, et al., 2021. Local tree diversity suppresses foliar fungal infestation and decreases morphological but not molecular richness in a young subtropical forest [J]. J Fungi, 7(3): 173.
SALMON YZ, LI XF, YANG B, et al., 2018. Surrounding species diversity improves subtropical seedlings carbon dynamics [J]. Ecol Evol, 8(14): 7055-7067.
SCHNABEL F, LIU X, KUNZ M, et al., 2021. Species richness stabilizes productivity via asynchrony and drought-tolerance diversity in a large-scale tree biodiversity experiment [J]. Sci Adv, 7(51): eabk1643.
SCHNEIDER GF, KOWALCZYK SW, CALADO VE, et al., 2010. DNA translocation through graphene nanopores [J]. Nano Lett, 10(8): 3163-3167.
SCHULDT A, ASSMANN T, BREZZI M, et al., 2018. Biodiversity across trophic levels drives multifunctionality in highly diverse forests [J]. Nat Commun, 9: 2989.
SCHULDT A, ASSMANN T, BRUELHEIDE H, et al., 2014a. Functional and phylogenetic diversity of woody plants drive herbivory in a highly diverse forest [J]. New Phytol, 202(3): 864-873.
SCHULDT A, ASSMANN T, SCHAEFER M, 2013a. Scale-dependent diversity patterns affect spider assemblages of two contrasting forest ecosystems [J]. Acta Oecol-Int J Ecol, 49: 17-22.
SCHULDT A, BARUFFOL M, BOHNKE M, et al., 2010. Tree diversity promotes insect herbivory in subtropical forests of south-east China [J]. J Ecol, 98(4): 917-926.
SCHULDT A, BARUFFOL M, BRUELHEIDE H, et al., 2014b. Woody plant phylogenetic diversity mediates bottom-up control of arthropod biomass in species-rich forests [J]. Oecologia, 176(1): 171-182.
SCHULDT A, BOTH S, BRUELHEIDE H, et al., 2011. Predator diversity and abundance provide little support for the enemies hypothesis in forests of high tree diversity [J]. PLoS ONE, 6(7): e22905.
SCHULDT A, BRUELHEIDE H, BUSCOT F, et al., 2017a. Belowground top-down and aboveground bottom-up effects structure multitrophic community relationships in a biodiverse forest [J]. Sci Rep, 7: 4222.
SCHULDT A, BRUELHEIDE H, DURKA W, et al., 2013b. Tree diversity promotes functional dissimilarity and maintains functional richness despite species loss in predator assemblages [J]. Oecologia, 174: 533-543.
SCHULDT A, BRUELHEIDE H, HRDTLE W, et al., 2012. Predator assemblage structure and temporal variability of species richness and abundance in forests of high tree diversity [J]. Biotropica, 44(6): 793-800.
SCHULDT A, EBELING A, KUNZ M, et al., 2019. Multiple plant diversity components drive consumer communities across ecosystems [J]. Nat Commun, 10: 1460.
SCHULDT A, FORNOFF F, BRUELHEIDE H, et al., 2017b. Tree species richness attenuates the positive relationship between mutualistic ant - hemipteran interactions and leaf chewer herbivory [J]. P Roy Soc B-Biol Sci, 284(1862): 20171489.
SCHULDT A, HONIG L, LI Y, et al., 2017c. Herbivore and pathogen effects on tree growth are additive, but mediated by tree diversity and plant traits [J]. Ecol Evol, 7(18): 7462-7474.
SCHULDT A, STAAB M, 2015. Tree species richness strengthens relationships between ants and the functional composition of spider assemblages in a highly diverse forest [J]. Biotropica, 47(3): 339-346.
SEIDELMANN KN, SCHERER-LORENZEN M, NIKLAUS PA, 2016. Direct vs. microclimate-driven effects of tree species diversity on litter decomposition in young subtropical forest stands [J]. PLoS ONE, 11(8): e0160569.
SEITZ S, GOEBES P, SONG Z, et al., 2016. Tree species and functional traits but not species richness affect interrill erosion processes in young subtropical forests [J]. Soil, 2(1): 49-61.
SEITZ S, GOEBES P, ZUMSTEIN P, et al., 2015. The influence of leaf litter diversity and soil fauna on initial soil erosion in subtropical forests [J]. Earth Surf Proc Land, 40(11): 1439-1447.
SHI NN, GAO C, ZHENG Y, et al., 2017. Effects of ectomycorrhizal fungal identity and diversity on subtropical tree competition [J]. J Plant Ecol, 10(1): 47-55.
SINGAVARAPU B, BEUGNON R, BRUELHEIDE H, et al., 2022. Tree mycorrhizal type and tree diversity shape the forest soil microbiota [J]. Environ Microbiol, 24(9): 4236-4255.
SONG ZS, SEITZ S, LI J, et al., 2019. Tree diversity reduced soil erosion by affecting tree canopy and biological soil crust development in a subtropical forest experiment [J]. For Ecol Manag, 444: 69-77.
SONG ZS, SEITZ S, ZHU PP, et al., 2018. Spatial distribution of LAI and its relationship with throughfall kinetic energy of common tree species in a Chinese subtropical forest plantation [J]. For Ecol Manag, 425: 189-195.
STAAB M, BLUTHGEN N, KLEIN AM, 2015. Tree diversity alters the structure of a tri-trophic network in a biodiversity experiment [J]. Oikos, 124(7): 827-834.
STAAB M, BRUELHEIDE H, DURKA W, et al., 2016. Tree phylogenetic diversity promotes host-parasitoid interactions [J]. P Roy Soc B-Biol Sci, 283(1834): 20160275.
STAAB M, LIU X, ASSMANN T, et al., 2021. Tree phylogenetic diversity structures multitrophic communities [J]. Funct Ecol, 35(2): 521-534.
STAAB M, METHORST J, PETERS J, et al., 2017. Tree diversity and nectar composition affect arthropod visitors on extrafloral nectaries in a diversity experiment [J]. J Plant Ecol, 10(1): 201-212.
STAAB M, SCHULDT A, ASSMANN T, et al., 2014a. Ant community structure during forest succession in a subtropical forest in South-East China [J]. Acta Oecol, 61: 32-40.
STAAB M, SCHULDT A, ASSMANN T, et al., 2014b. Tree diversity promotes predator but not omnivore ants in a subtropical Chinese forest [J]. Ecol Entomol, 39(5): 637-647.
SUN ZK, LIU XJ, SCHMID B, et al., 2017. Positive effects of tree species richness on fine-root production in a subtropical forest in SE-China [J]. J Plant Ecol, 10(1): 146-157.
TANG T, ZHANG N, BONGERS FJ, et al., 2022. Tree species and genetic diversity increase productivity via functional diversity and trophic feedbacks [J]. eLife, 11: e78703.
TROGISCH S, SALMON Y, HE JS, et al., 2016. Spatio-temporal water uptake patterns of tree saplings are not altered by interspecific interaction in the early stage of a subtropical forest [J]. For Ecol Manag, 367: 52-61.
WANG MQ, LI Y, CHESTERS D, et al., 2020. Multiple components of plant diversity loss determine herbivore phylogenetic diversity in a subtropical forest experiment [J]. J Ecol, 107(6): 2697-2712.
WANG MQ, YAN C, LUO A, et al., 2022. Phylogenetic relatedness, functional traits, and spatial scale determine herbivore co-occurrence in a subtropical forest [J]. Ecol Monogr, 92(1): e01492.
WEINHOLD A, DOLL S, LIU M, et al., 2022. Tree species richness differentially affects the chemical composition of leaves, roots and root exudates in four subtropical tree species [J]. J Ecol, 110(1): 97-116.
WU D, PIETSCH KA, STAAB M, et al., 2021a. Wood species identity alters dominant factors driving fine wood decomposition along a tree diversity gradient in subtropical plantation forests [J]. Biotropica, 53(2): 643-657.
WU D, STAAB M, YU M, 2021b. Canopy closure retards fine wood decomposition in subtropical regenerating forests [J]. Ecosystems, 24(8): 1875-1890.
WU YT, GUTKNECHT J, NADROWSKI K, et al., 2012. Relationships between soil microorganisms, plant communities, and soil characteristics in Chinese subtropical forests [J]. Ecosystems, 15(4): 624-636.
WU YT, WUBET T, TROGISCH S, et al., 2013. Forest age and plant species composition determine the soil fungal community composition in a Chinese subtropical forest [J]. PLoS ONE, 8(6): e66829.
XUE YJ, CHENG AP, LI S, et al., 2023. The effects of environment and species diversity on shrub survival in subtropical forests [J]. Biodivers Sci, 31(3): 22443.[薛玉潔, 程安鹏, 李珊, 等, 2023. 亚热带森林中环境和物种多样性对灌木存活率的影响 [J]. 生物多样性, 31(3): 22443.]
YANG B, LI B, HE YX, et al., 2018. Tree diversity has contrasting effects on predation rates by birds and arthropods on three broadleaved, subtropical tree species [J]. Ecol Res, 33(1): 205-212.
YANG B, LI Y, DING BY, et al., 2017. Impact of tree diversity and environmental conditions on the survival of shrub species in a forest biodiversity experiment in subtropical China [J]. J Plant Ecol, 10(1): 179-189.
YANG B, LIANG Y, SCHMID B, et al., 2022. Soil fungi promote biodiversity-productivity relationships in experimental communities of young trees [J]. Ecosystems, 25: 858-871.
YANG X, BAUHUS J, BOTH S, et al., 2013. Establishment success in a forest biodiversity and ecosystem functioning experiment in subtropical China (BEF-China) [J]. Eur J For Res, 132(4): 593-606.
ZHANG J, QIAN H, GIRARDELLO M, et al., 2018. Trophic interactions among vertebrate guilds and plants shape global patterns in species diversity [J]. P Roy Soc B-Biol Sci, 285(1883): 20180949.
