N6-甲基腺苷修饰在肾脏疾病中的研究进展
2023-09-19综述曾彩虹审校
林 瑶 综述 曾彩虹 审校
N6-甲基腺苷(m6A)修饰是指在RNA的腺苷N6位点添加一个甲基,是真核 RNA 中最普遍、最丰富和最保守的内部共转录修饰,在转录后水平的RNA剪接、转运、稳定性和翻译等下游分子事件和生物学功能的调节中发挥着关键作用。m6A修饰的失调与代谢异常、心血管疾病、糖尿病及肿瘤等有关。最近对人类、动物模型和细胞水平的研究揭示了m6A修饰在肾脏相关疾病中的关键作用。全面了解肾脏疾病中m6A调控的复杂性将有助于我们了解肾脏疾病的发病机制。本文简述了m6A修饰在肾脏疾病中的调节作用及其临床意义。
m6A修饰
表观遗传学是对 DNA、组蛋白和RNA水平的可逆、动态修饰,主要包括DNA甲基化、组蛋白乙酰化、RNA修饰和染色质重排[1]。在表观遗传学的修饰中,DNA甲基化和组蛋白修饰已经得到深入研究,而RNA修饰则是一个新兴领域。细胞RNA[包括核糖体(rRNA)、转运RNA(tRNA)、小核RNA(snRNA)、信使RNA(mRNA)和长链非偏码RNA(lncRNA)]中存在100多种转录后修饰[2],主要包括 m6A、N1-甲基腺苷 (m1A)、5-甲基胞嘧啶(m5C)、假尿苷(Ψ)、N7-甲基鸟苷(m7G)等,这些修饰在调控RNA的翻译、稳定、剪切、转运及定位等方面发挥重要作用[3-4]。
RNA甲基化修饰是真核生物中最常见的RNA修饰类型,占总RNA修饰的60%。其中以m6A 修饰最常见,约占所有甲基化核糖核苷酸的50%。据估计,m6A存在于0.1%~0.4%的腺苷中,能够修饰哺乳动物细胞中的7 000 多个mRNA[5],被认为是最普遍、可逆和动态的真核mRNA转录修饰。
m6A修饰是一种可逆的动态修饰,主要涉及3种蛋白(图1):甲基转移酶、去甲基化酶、结合蛋白。其中,甲基基团的安装是通过高度保守的甲基转移酶复合物(MTC) 完成的,该复合体包括含有S-腺苷甲硫氨酸(SAM)结合基序的甲基转移酶3(METTL3)、甲基转移酶14(METTL14)和稳定MTC定位、底物募集的调节亚基——Wilms肿瘤 1相关蛋白(WTAP)[6-9]。去甲基化酶介导甲基化的逆转,主要包括ALKB同源物5(ALKBH5)和 ALKB亚家族的肥胖相关基因(FTO),它们以Fe2+和 a-酮戊二酸依赖性方式催化m6A的去甲基化[10]。结合蛋白主要包括 YT521-B 同源结构域家族(YTHDF1/2/3、YTHDC1/2)、异质核核糖核蛋白(HNRNPA2/B1)和胰岛素样生长因子2 mRNA 结合蛋白(IGF2BP1/2/3),通过识别 m6A 修饰的靶向 RNA 发挥作用[11]。最近的研究表明,m6A修饰普遍存在于真核生物RNA上,且在生理和病理条件下都发挥着重要作用,因此,m6A甲基化修饰在肾脏疾病中的作用机制值得探究。
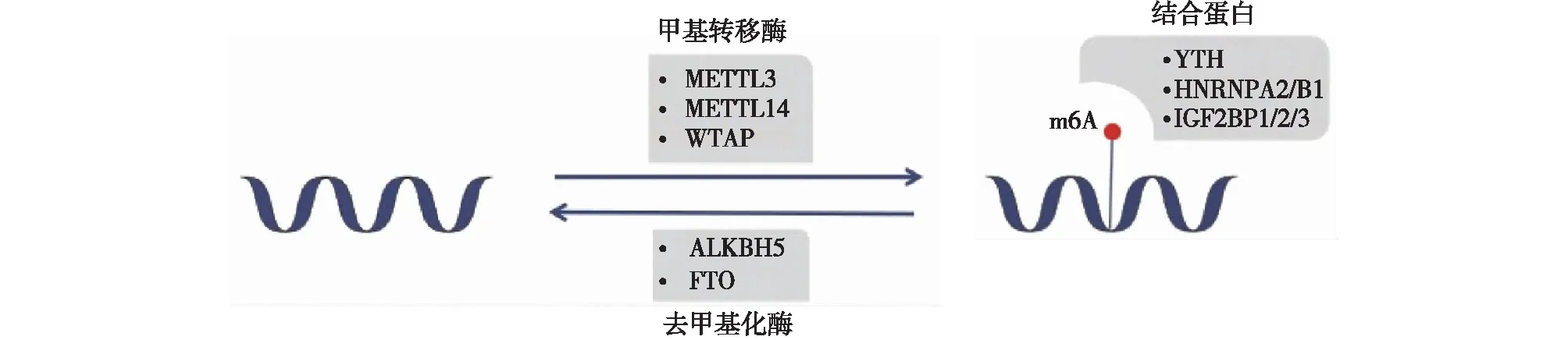
图1 m6A甲基化修饰
m6A甲基化修饰与肾脏疾病
越来越多的研究证实m6A甲基化参与肾脏疾病的发生和发展。例如,在顺铂诱导的急性肾损伤(CI-AKI)体内外模型中,发现m6A的水平显著升高[12]。同时结合CI-AKI小鼠肾组织的RNA甲基化测序(MeRIP-seq)结果、基因调控及GO富集分析和KEGG通路分析,研究人员还发现了显著改变的m6A修饰主要富集在代谢、细胞死亡、氧化还原和转运过程中,而这些恰好是CI-AKI的生物学和病理学基础[13],这为AKI的研究和治疗方案的创新提供了新思路。
此外,Wang等[14]发现在慢性肾脏病(CKD)患者外周血单个核细胞中m6A水平显著降低,为CKD相关细胞功能障碍的潜在机制提供了新的见解。在狼疮性肾炎(LN)中,研究人员利用LASSO-Logistic回归分析、单样本基因集富集分析和基因集变异分析等研究手段发现,m6A调控因子在肾小球中的差异表达最显著,并与自然杀伤细胞激活、免疫微环境等相关,其中7种m6A相关蛋白被证实与肾小球滤过率(GFR)有显著相关性,提示其可能是潜在的预后生物标志物,这为LN新治疗方案的研发提供了基础[15]。研究人员还比较了正常葡萄糖耐受者、2型糖尿病(T2DM)患者和糖尿病肾病(DN)患者尿液中m6A水平,发现DN患者尿液中m6A水平发生了显著下调,并随DN的恶化而进一步降低,这提示尿液中m6A水平可作为诊断DN的前瞻性生物标志物[16]。同时,Yin等[17]利用GEO数据库中的差异表达基因、CIBERSORT算法和加权基因共表达网络分析筛选出DN患者m6A甲基化相关的43个关键基因,其中一些关键基因的异常表达与肾脏肥大、蛋白尿和肾小球硬化的发生有关,为DN的诊断和治疗提供了依据。此外,还有研究证实了多种m6A调控因子,如WTAP、RBM15、FTO等能够调节各种lncRNA的甲基化和表达,影响M1巨噬细胞的表达,参与DN的炎症和免疫过程[18]。
尽管m6A在肾脏疾病中的作用、机制和潜在靶点备受关注,但目前对m6A的研究仍处于初步和前沿探索阶段,靶点的功能复杂且不确定。因此,全面描述m6A与肾脏疾病的相关性,即其在肾脏疾病相关的细胞过程中特定位点的改变,有望为肾脏疾病潜在靶点的识别和治疗策略的建立提供重要指导。
甲基转移酶与肾脏疾病
METTL3 最近,Wang等[19]在AKI患者肾活检组织和体内外模型中发现了METTL3表达的上调,并且上调的METTL3通过依赖IGF2BP2的方式增强了转化生长因子β(TGF-β)激活激酶1结合蛋白3(TAB3)mRNA的m6A修饰,增强了TAB3 mRNA的稳定性以促进其表达,通过促进炎症反应加速AKI的进展;同时,METTL3的遗传学和药理学抑制均能够减轻肾脏炎症和损伤,表明METTL3/TAB3轴是治疗AKI的潜在靶点。还有一项研究表明,METTL3能够直接结合mmu-lncRNA 121686和hsa-lncRNA 520657的m6A位点,并增强其修饰和表达,从而促进lncRNA对miR-328-5p的直接抑制作用,加重了高温需求因子a3(Htra3)介导的管状上皮细胞凋亡和缺血性、脓毒症和万古霉素诱导的AKI。这一发现再次验证了METTL3介导的m6A修饰在AKI进展中的靶点潜力和干预价值[20]。
Liu等[21]在梗阻性肾病(ON)患者肾纤维化组织中发现了人肺腺癌转移相关转录本1(MALAT1)表达的上调,紧接着他们利用RNA结合蛋白免疫沉淀(RIP)、RNA甲基化免疫共沉淀 (MeRIP)、生物信息学分析和荧光素酶报告基因实验证实,METTL3介导的MALAT1的m6A修饰能够促进MALAT1的表达,从而通过MALAT1/miR-145/FAK通路参与TGF-β1诱导的体内和体外肾纤维化。因此,调控METTL3能够以m6A依赖的方式激活MALAT1/miR-145/FAK轴,从而促进肾脏纤维化,这为ON的研究提供了新的线索。此外,在ON体内体外模型中,研究人员发现高表达且核局域化的METTL3也能在HNRNPA2B1的协助下介导pri-miR-21多个位点的m6A修饰,从而能够促进迪乔治综合征危象区基因8(DGCR8)对pri-miR-21的招募、切割和成熟以及miR-21-5p的形成。随后,miR-21-5p激活SPRY1/ERK/NF-κB信号通路,驱动炎症,促进管状结构塌陷、胶原沉积和ON[22]。上述研究结果为探索肾脏纤维化提供了更深入的见解,显示靶向调控因子介导的m6A修饰在肾脏纤维化治疗中的重要潜力。
Jiang等[23]在DN患者和糖尿病小鼠的肾脏中发现了METTL3介导的m6A修饰的显著上调。并且还发现METTL3通过依赖IGF2BP2的方式促进了组织金属蛋白酶抑制因子2(TIMP2)mRNA的m6A修饰,增强了TIMP2 mRNA的稳定性,促进其翻译。而TIMP2的上调能够激活Notch信号通路,诱发足细胞凋亡和炎症反应,引起足细胞损伤和丢失,促进蛋白尿和肾小球病变,从而促进DN进展。因此干预METTL3的表达对足细胞损伤、肾脏炎症和蛋白尿的产生以及DN的发展具有潜在价值。
此外,还有研究发现,饮食中限制蛋氨酸或SAM的摄入会显著降低METTL3的表达,从而降低c-Myc和精氨酸加压素受体2(Avpr2)mRNA的m6A水平,抑制其翻译和信号传导,最终抑制蛋白质合成和囊肿增殖[24]。通过饮食限制靶向调控METTL3和m6A修饰,为多囊肾以及肾脏疾病的治疗提供了一种新的策略和独特的见解。
METTL14 研究发现METTL14通过m6A依赖性机制增强Yes相关蛋白1(YAP1)mRNA的甲基化,并通过加速YAP1 mRNA的衰减来降低其表达水平,从而抑制YAP1-TEAD信号,促进缺血缺氧性AKI的进展[25]。Lu等[26]在局灶节段性肾小球硬化和DN患者肾活检样本及蛋白尿性肾病的体内体外模型中发现了m6A 和METTL14水平的显著上调;更深入的研究表明,METTL14通过YTHDF2依赖的方式增加了抗衰老酶1(Sirt1)mRNA的m6A修饰,引起Sirt1 mRNA的降解和蛋白表达的下调。而Sirt1的下调会抑制自噬,促进足细胞凋亡和炎症反应,最终加重DN中蛋白尿和足细胞病变的进展。Xu等[27]还发现低水平的METTL14能够直接降低磷酸酶和张力蛋白同源蛋白(PTEN)mRNA的甲基化,导致PTEN mRNA和蛋白水平的显著下调,从而激活PI3K/Akt信号通路,引起组蛋白脱乙酰化酶5(HDAC5)的上调并促进其所介导的小管细胞上皮间质转化(EMT)和DN进展。此外,METTL14还能增强α-klotho mRNA的m6A修饰,抑制其mRNA和蛋白表达,从而加重高糖诱导的肾小球内皮细胞损伤和炎症,促进DN进展[28]。METTL14的敲除或过表达可通过m6A依赖机制逆转异常变化,故靶向METTL14介导的m6A有望成为治疗肾脏疾病足细胞和小管细胞损伤的靶点。
去甲基化酶与肾脏疾病
FTO在CI-AKI的体内外研究中,FTO抑制剂甲氯芬那酸(MA)能够进一步降低受抑制的FTO,增强m6A修饰,促进p53 mRNA和蛋白的表达,加速肾小管细胞凋亡和AKI的发生[12]。相反,FTO的过表达可降低m6A修饰水平和p53蛋白表达,最终减少肾小管凋亡和逆转肾损伤。此外,FTO还被发现通过影响多个基因的m6A修饰,促进纤维化反应、EMT和ON的发展[29];Li等[30]在TGF-β1处理的细胞模型和单侧输尿管梗阻的小鼠模型中,发现FTO mRNA和蛋白质表达的上调,能够减少lncRNA GAS5的m6A修饰,促进人肾皮质近曲小管上皮细胞(HK-2)的EMT和炎症反应,引起肾脏间质纤维化。在酒精诱导的肾脏损伤中,FTO的低表达以YTHDF2依赖的方式增加过氧化物酶体增殖物激活受体α(PPAR-α)mRNA的m6A修饰,最终导致NLRP3炎症小体的激活和肾脏中NF-κB驱动的炎症反应[31]。在T2DM患者外周血中发现FTO mRNA表达水平升高,而m6A显著降低,提示低表达的m6A可能是T2DM的一种新的潜在生物标志[32]。最近,Sun等[33]对DN患者肾组织进行组织病理学的高维度单细胞分析以及成像质谱流式分析,发现FTO的显著减少抑制了细胞因子信号传导抑制蛋白1(SOCS1)mRNA和蛋白质的表达,促进Janus激酶2(JAK2)和信号转导及转录激活因子3(STAT3)的磷酸化,引起肾脏的炎症反应和损伤。以上这些研究为与m6A相关肾脏疾病的治疗提供了新的潜在靶点和证据。然而,还需要更深入的实验来验证。
ALKBH5最近的一项研究表明[34],在ON小鼠模型中,低水平的ALKBH5会增加肾组织总m6A水平,从而促进EMT和炎症病理过程,最终影响肾脏间质纤维化。此外,染料木黄酮可减少肾组织中ALKBH5的损失,减少总m6A修饰,从而在一定程度上降低肾纤维化相关蛋白的表达、EMT和炎症反应,从而抑制ON的进展。重要的是,这些变化与沉默和过表达ALKBH5的结果是一致的,进一步显示了ALKBH5在阻塞性肾纤维化研究中的潜力。然而,目前还缺乏对其完整的全链机制的探索。
结合蛋白与肾脏疾病
结合蛋白主要与发生了m6A甲基化修饰的位点结合,从而介导其功能的实现。目前对结合蛋白的研究主要是在甲基转移酶和去甲基化酶的基础上开展的。然而,最近有研究表明,阅读蛋白在肾脏纤维化过程中起着重要作用。在阻塞性和大剂量叶酸诱导的纤维化肾脏中,YTHDF1的表达显著升高,并与纤维化程度和YAP水平呈正相关[35]。在此基础上,降低YTHDF1可显著抑制YAP的表达,抑制纤维化相关分子的上调,从而缓解纤维化进展,这提示抑制YTHDF1可能是缓解肾脏纤维化的潜在靶点。
总结和展望
m6A是RNA最主要的表观遗传修饰类型,几乎参与RNA代谢的所有过程。大量的研究表明其参与多种疾病的发生和发展,为相关疾病的临床早期防治提供了靶点和方向。高糖、氧化应激、肾毒性物质等刺激能够引起肾脏内m6A修饰水平的改变,通过调控 RNA的转运、稳定性和可变剪接等,引起细胞状态及内环境的改变,这些生物学功能改变与AKI、ON、DN等疾病的进程密切相关。虽然目前 m6A修饰与肾脏疾病的联系仍有许多空白有待填补,其在相关疾病中的具体作用和分子机制也有待阐明,但是随着对表观遗传学的了解以及高通量测序技术的发展,相信未来靶向m6A修饰因子的研究可以为肾脏疾病的诊断、治疗及预后评估提供帮助。
