基于SRAP分子标记的春大豆杂交种核心亲本杂种优势群划分
2023-09-07韩博文杨绪磊林春晶赵丽梅吴松权张春宝
韩博文 姜 楠 杨绪磊 林春晶 彭 宝 赵丽梅 吴松权 张春宝*
(1.延边大学 农学院,吉林 延吉 133002;2.吉林省农业科学院 大豆研究所/农业农村部杂交大豆育种重点实验室,长春 130033)
大豆(Glycinemax(L.) merr.)是我国重要的油料、粮食及饲料作物,营养价值丰富,应用领域广阔[1]。随着近年来社会经济的高速发展,我国对大豆的需求日益增高,然而国内大豆种植面积和产量均有所下降,严重依赖国外进口。根据海关总署公开数据,2020年,我国大豆进口量首次突破1亿t;2022年中央一号文件提出加力扩种大豆油料,这使得大豆种植面积创下了1958年以来新高,进口量降至9 108万t,但我国大豆对外依存度仍然在80%以上的高位。由于我国耕地资源有限,提高单位面积产量是提升大豆产能的有效出路。杂种优势利用是快速提高作物单产的有效途径,已在水稻、玉米、高粱和棉花等多种作物的品种选育中获得成功[2-4]。大豆同样具有较强的杂种优势,开展杂交大豆方面的研究对于提升大豆单产具有重要意义。我国从20世纪八十年代就开始大豆杂种优势利用研究,于2002年审定了世界上第一个大豆杂交种‘杂交豆1号’,在区域试验中较对照平均增产21.9%[5],目前已审定的杂交大豆品种近40个,区试增产幅度达5.3%~22.7%,大豆杂种优势的利用对增产效果明显[6]。
亲本资源是杂交种选育的基础,然而在众多材料中选择适合的亲本并配制强优势杂交组合,是杂交种选育考虑的首要问题,关系到其产出效率[7]。杂种优势群理论由Melchinger等[8]提出,指来自相同或不同群体的一组相关或不相关基因型,当与其他遗传背景不同的种质群的基因型杂交时,能表现出相似的配合力和杂种优势。杂种优势群理论已在玉米、油菜和小麦等作物中被广泛应用[9-11],其主要划分方法有表型聚类分群法、配合力分群法、分子标记聚类分群法等[12]。白志元等[13]利用64对SSR分子标记将68份杂交大豆亲本中的恢复系和保持系分别聚为两大类5个亚类,并发现聚类结果与地理来源具有一定相关性。雷蕾等[14]利用14个产量性状相关SSR分子标记将30个已审定杂交大豆品种的43份亲本分为2个类群,其中25个杂交种的亲本分属于2个类群。
SRAP分子标记技术能准确地分析大豆材料的遗传多样性和亲缘关系,但利用其对杂交大豆亲本进行优势群的划分鲜见报道。本研究利用条带清晰、多态性好的12对SRAP分子标记组合,对东北三省春大豆区育成的100份核心亲本进行遗传多样性评价和亲缘关系划分,并利用已审定的10个强优势杂交种的亲本进行杂种优势群验证,旨在明确杂交大豆核心亲本的遗传多样性特点及群体结构,以期为春大豆杂交种亲本的遗传改良及杂交组合配制提供参考。
1 材料与方法
1.1 试验材料
1.1.1植物材料
所用材料包括来源于中国(东北三省)、美国和意大利的49份不育系的同型保持系与51份恢复系,共100份杂交大豆亲本材料(表1),均为吉林省农业科学院杂交大豆研究团队选育及保存。所有试验材料种植于吉林省长春市范家屯镇杂交大豆核心育种基地(124°83′ E, 43°43′ N)。
1.1.2SRAP引物
使用8个正向引物和6个反向引物(表2),共计48种组合,通过前期筛选获得条带清晰,多态性好的12种组合。本研究利用上述引物组合进行100份亲本材料的多态性分析。
1.2 试验方法
1.2.1DNA提取和PCR
从田间剪取亲本材料的二节期(V2)叶片,共100份,液氮研磨后,使用CTAB法[21]提取DNA。经0.8%琼脂糖凝胶检验,条带清晰无拖尾,符合SRAP试验要求,置于-20 ℃冰箱保存备用。利用12种SRAP引物(表3)组合对亲本DNA进行PCR扩增,PCR反应体系为25 μL,其中DNA 模板2.0 μL,正向反向引物各0.5 μL(10 μmol/L),dNTPs 0.5 μL(10 mmol/L,北京鼎国),10×PCR buffer 2.5 μL(北京鼎国),Taq酶0.5 μL(2 U/μL,北京鼎国),ddH2O 18.5 μL。DNA扩增程序:94 ℃预变性5 min;94 ℃变性1 min,35 ℃退火1 min,72 ℃延伸1 min,5个循环;94 ℃变性1 min,50 ℃退火1 min,72 ℃延伸1 min,35个循环;72 ℃延伸10 min。利用4%聚丙烯酰胺凝胶电泳检测后,使用ABI PRISM 377 sequencer测序仪检测SPAP多态性结果,通过GENESCAN软件进行分析,在70~500 bp片段的DNA Marker范围内,每2个碱基读数1次,并记录最终条带数据。

表3 SRAP分子标记多态性分析结果Table 3 SRAP molecular marker polymorphism analysis results
1.2.2数据处理
双边匹配是研究由不可分对象构成的两不相交集合中主体的相互匹配过程,起源于Gale等[1]对学生入学和婚姻匹配问题的研究,该文开启了双边匹配研究的先河,奠定了双边匹配的理论基石。现实社会中存在着大量的双边匹配问题,如劳动力市场中的员工与岗位匹配[2]、金融市场中的风险投资匹配[3]、电子商务中的买卖交易匹配[4]、供应链管理[5]及知识服务[6]中的供需匹配等。由此可见,深入研究双边匹配问题具有十分重要的理论意义和实际应用价值。
对原始数据进行处理,将有条带的位点标记为1,无条带的位点标记为0,转化为0、1矩阵。使用NTsys2.10e软件,计算遗传相似系数(genetic similarity,GS),利用算术平均非加权方法(unweighted pair-group method with arithmetic means,UPGMA)对亲本材料进行聚类分析,获得聚类树状图。利用Popgene 32软件计算每个标记的多态性信息含量(polymorphism information content,PIC),公式如下:
式中:Pi为群体中含有第i个等位变异的比例。主效基因频率(major alle frequency,MAF)、有效等位基因数(Effective number of alleles,Ne)、基因多样性指数(gene diversity,h)、Shannon信息指数(Shannon’s Information index,I)。根据SRAP标记对100份亲本材料分析得到的遗传相似系数及分群结果,利用R语言ggplot 2作主坐标分析(principal co-ordinates analysis,PCoA),最终将各点PCoA结果坐标绘制在二维平面上,制成散点图。
2 结果与分析
2.1 SRAP分子标记多态性分析
利用12对多态性较好的引物组合,包括Me 1×Em 3、Me 1×Em 5、Me 2×Em 3、Me 2×Em 7、Me 3×Em 3、Me 4×Em 4、Me 5×Em 1、Me 5×Em 8、Me 6×Em 4、Me 6×Em 8、Me 7×Em 5和Me 8×Em 1,对49份保持系和51份恢复系进行多态性分析(表3)。通过检测,共扩增出2 135条条带,其中多态性条带2 130条,每对引物组合扩增出的多态性谱带数为137~204条,平均为178条,多态性位点百分率平均值高达99.76%,PIC含量范围在0.074 0~0.153 7,平均值为0.125 7,按照Botstein等[22]的标准,PIC<0.25表明SRAP分子标记携带信息量较少,100份亲本材料间遗传差异较大。

2.2 100份亲本的聚类分析
由表4可知,100份杂交大豆亲本材料间的GS在0.329 2~0.637 6,平均GS为0.473 3。GS最小的是来自黑龙江的JLCMS112B和美国材料JLCMS164B,GS最大的是来自美国的JLR60和JLR1。
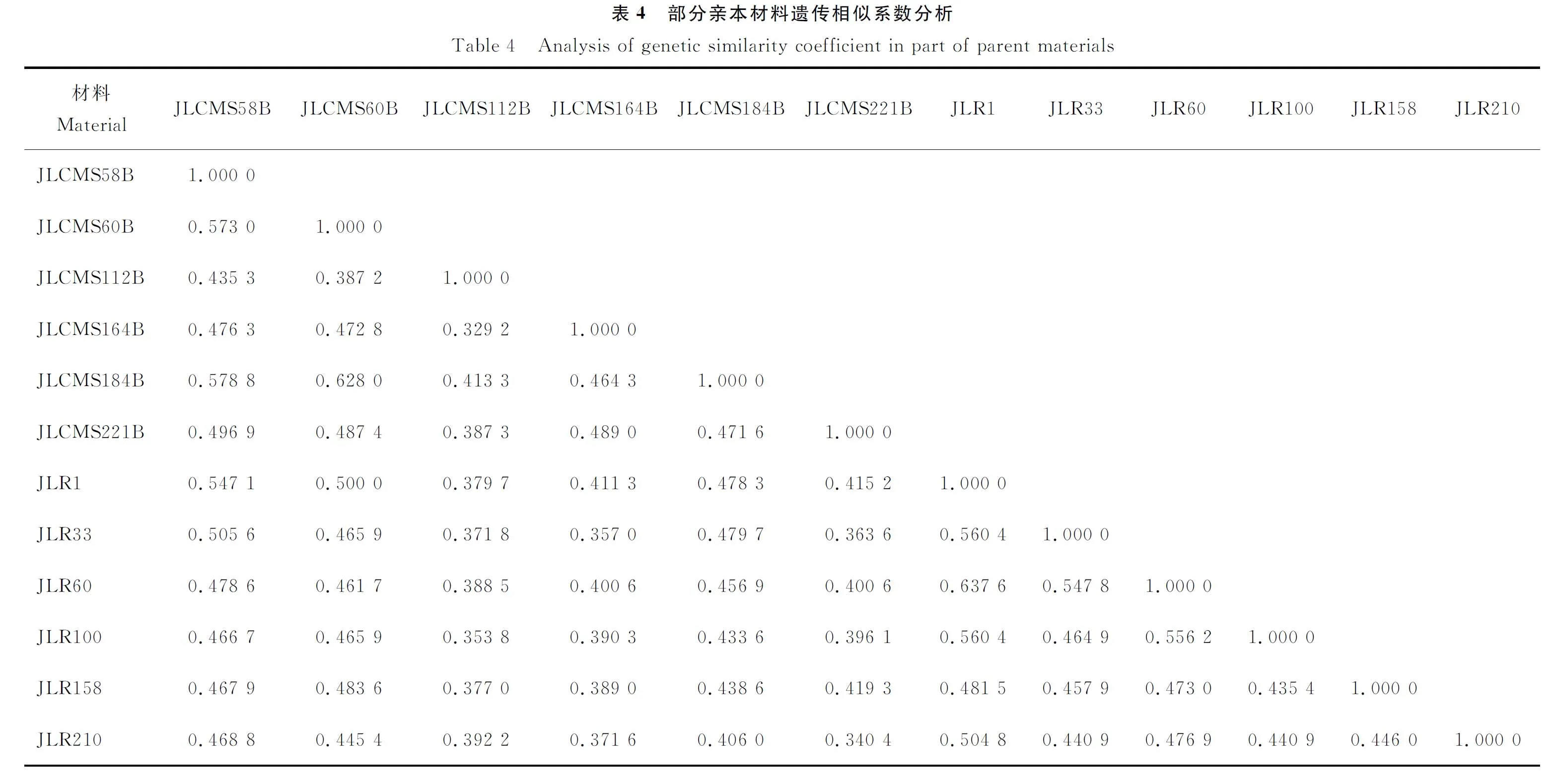
由图1可知,在GS为0.450 0处可划分为I和II两大类群,在GS为0.460 0处,类群I可划分为2个亚群I-1和I-2,在GS为0.480 0处,类群II可划分为II-1和II-2亚群。第I类群全部为保持系,共49份材料,其亚群I-1中有20个保持系,群体样本数最少,主要以中国东北背景为主,包括有JLCMS103B、JLCMS91B、JLCMS188B、JLCMS9B等17个保持系,仅含JLCMS42B、JLCMS101B、JLCMS171B这3个美国背景的保持系;亚群I-2中有29份保持系,包括中国东北的JLCMS185B、JLCMS48B、JLCMS84B等20份保持系以及美国来源的JLCMS199B、JLCMS181B、JLCMS176B等10份保持系。51份恢复系全部被划入第II类群,其中亚群II-1有26份恢复系,包括JLR522、JLR531、JLR12等20个来源美国的恢复系,JLR194、JLR192、JLR22等6份来源中国东北的恢复系;亚群II-2有25份恢复系,包括JLR46、JLR119、JLR107等来源美国的恢复系12份,来源意大利的1份为JLR8,来源中国东北的有10份,包括JLR210、JLR547、JLR209等。
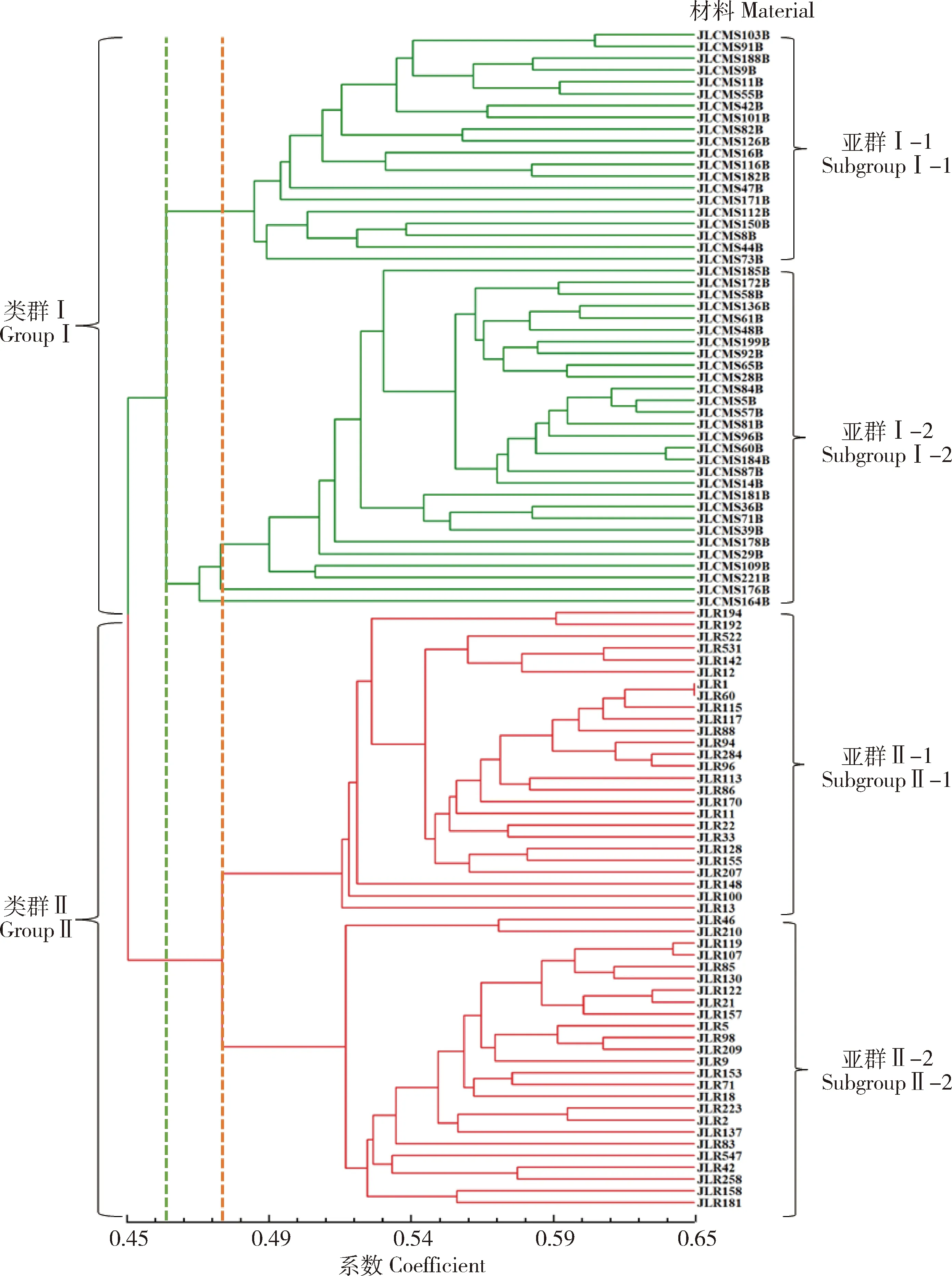
GS为0.460 0处的绿色虚线将类群Ⅰ划分为Ⅰ-1和Ⅰ-2两个亚群; GS为0.480 0处的红色虚线将类群Ⅱ划分为Ⅱ-1和Ⅱ-2两个亚群。The green dotted line at GS 0.460 0 divides group I into two subgroups, I-1 and I-2; The red dotted line at GS 0.4800 divides Group II into two subgroups, II-1 and II-2.图1 SRAP标记对亲本材料的UPGMA聚类分析Fig.1 UPGMA cluster analysis of SRAP markers on parent materials
2.3 强优势杂交种亲本的群内验证
由表5可知,所有母本保持系均来自于类群I,其中3份母本划入亚群I-1,4份母本划入亚群I-2;所有父本恢复系都被划入类群II,其中3份父本划入亚群II-1,3份父本划入亚群II-2,表明10个强优势杂交种的13份亲本均来自本研究划分的两大类群。进一步对亲本来源地信息进行分析发现,所有母本均来源于中国东北的黑龙江和吉林;而所有父本的来源地均为国外,分别是美国和意大利。上述结果表明,10个强优势杂交种亲本均来自于本研究所划分的2个类群,且为中国(♀)×外国(♂)组合,说明通过SRAP分子标记划分的杂种优势类群结果与地理来源分布基本一致,且不同来源地之间选配有助于充分发挥大豆杂种优势。从亲本间遗传相似程度分析发现,13份杂交种亲本的遗传相似性差异较大,GS在0.387 9~0.507 5,说明强优势杂交种亲本间遗传多样性较为丰富。

表5 已审定的10个强优势杂交种的分群结果Table 5 Group division results of 10 released strong heterosis hybrids
2.4 100份亲本的遗传多样性分析
由表6可知,总群的主效基因频率(MAF)均值为0.927 0,类群I的MAF为0.927 5,其中亚群I-1的MAF为0.926 1,亚群I-2的MAF为0.928 4;类群II的MAF为0.926 6,其中亚群II-1的MAF为0.931 0,亚群II-2的MAF为0.921 7。相较于类群和亚群间的MAF差异不大。总群的有效等位基因数(Ne)为1.156 9,类群I的Ne为1.115 7,其中亚群I-1的Ne为1.159 1,亚群I-2的Ne为1.153 8;类群II的Ne为1.158 2,其中亚群II-1的Ne为1.158 2,亚群II-2的Ne为1.169 3。类群Ⅱ的Ne高于类群Ⅰ,亚群I-1和亚群II-2的等位变异数更高。总群的基因多样性指数(h)为0.135 1,其中类群I的h为0.134 1,其亚群I-1和I-2的h分别为0.136 4和0.132 3;类群II的h为0.135 0,其亚群II-1和II-2的h为分别为0.126 7和0.143 8。类群Ⅰ和Ⅱ的总体基因多样性相似,其中亚群II-2的基因多样性丰度高于II-1。总群I的h为0.260 2,其中类群I的h为0.258 6,其亚群I-1和I-2的h分别为0.261 6和0.255 5;类群II的I为0.260 0,其亚群II-1和II-2的I分别为0.244 3和0.272 6。类群间的I无显著差异,亚群I-1和II-2的物种多样性丰度大于亚群I-2和II-1。总群多态信息量(PIC)为0.126 2,其中类群I的PIC为0.125 4,其亚群I-1和I-2的PIC分别为0.127 5和0.124 1;类群II的PIC为0.126 8,其亚群II-1和II-2的PIC分别为0.120 2和0.133 9。表明类群I与类群II存在较丰富的遗传多样性,4个亚群中,亚群I-1的遗传多样性丰度大于亚群I-2;亚群II-2的遗传多样性丰度则大于亚群II-1。

表6 不同群内亲本材料的遗传多样性分析Table 6 Genetic diversity analysis in different populations of parent material
由图2可知,主坐标1(Pco1)和主坐标2(PCo2)分别解释了38.29%和19.96%的群体遗传变异。通过Pco1坐标结果对100份亲本划分的4个亚群进行分析,其中亚群I-1和I-2重叠位于PCo1为-0.06~0.01,表明2个亚群间可能存在遗传亲缘的交叉。亚群II-1和II-2重叠位于Pco1在0.00~0.04,2个亚群间遗传亲缘的交叉更为显著。这表明杂交大豆亲本在改良过程中,存在同一类群内的不同亚群间亲缘混合的情况。类群Ⅰ与类群Ⅱ的遗传分布之间几乎没有重合,说明2个类群间遗传亲缘存在差距,并且类群Ⅰ的2个亚群的遗传距离分布范围更广,散点密集度较小,证明类群Ⅰ的遗传资源更为丰富。

图2 100份核心亲本材料主坐标分析Fig.2 Principal co-ordinate analysis of one hundred core parent materials
3 讨 论
3.1 杂交大豆亲本遗传多样性
本研究利用SRAP分子标记根据遗传相似系数对100份杂交大豆亲本材料进行了聚类分析,并划分了两大类群,即主要由中国东北保持系材料构成的类群I和主要来源于美国恢复系材料的类群II;每个类群还可进一步划分为2个亚群。从来源地来看,尽管类群I包含的材料主要来自中国东北,但也有部分美国材料;类群II主要来自美国,但同样也有部分中国东北材料。李琼等[23]对黄淮海50份大豆资源的SSR分子标记分析发现,大豆类群的形成与其来源地地理环境与生态类型相关。徐泽俊[24]对303份大豆种质进行聚类分析发现,同一来源地的种质不一定全都聚为一个群。造成上述情况的原因可能是由于中国是大豆原产国,早期由于资源外流,国外大豆亲本的先祖源于我国,而我国近几十年来对国外资源的引进及杂交利用,使得部分优异材料混有国外遗传背景。本研究所用亲本基本上符合按不同来源地划分类群,但在亚群的划分并没有按地理来源而细化成诸如黑龙江省亚群或吉林省亚群,表明东北大豆材料遗传关系较近。这与张振宇[25]对50份东北大豆骨干亲本基于聚类分析结果显示东北大豆材料间遗传同源性较大,遗传差异较小的结论相一致。
3.2 大豆杂种优势群的验证及利用
在本研究中,100份大豆亲本之间的GS在0.329 2~0.637 6,R2为0.473 3,于0.450 0处划分为2个类群,又在遗传相似系数0.460 0和0.480 0处分别划分为2个亚群。这与白志元等[13]基于SSR引物将68个大豆品种根据分析得出的GS聚类分析,将材料划分为两大类5个亚类的结果相似。雷蕾等[14]通过SSR标记将已审定强优势大豆杂交种的亲本划分为来源于中国东北与国外材料的2个大群,且GS在0.400 0~0.600 0时,杂种优势利用程度较高。本研究所分析的10个强优势杂交种的13份亲本同样是分别来自于中国×外国组合,其GS在0.387 9~0.507 5,表明利用SSR标记和SRAP标记均可较为明确的划分大豆杂种优势群,且能得出相近的试验结果;同时发现,目前已育成的强优势杂交种的亲本组合并非遗传相似系数越小越好,与Moll等[26]提出的杂交种亲本不应超过最佳遗传距离的结论相符。然而Boeven等[27]对1 903个小麦杂交组合的分析发现,其杂种优势随着亲本遗传距离的增加而稳步增加。因此,后续需要配制更多的杂交组合,获得更多的产量数据,结合亲本间的GS数据,更加全面的评估大豆杂种优势与亲本间遗传距离的相关性。
4 结 论
1)SRAP标记共扩增出2 135条条带,其中多态性条带2 130条,多态性位点占比99.76%,每对引物组合扩增出的多态性谱带数为137~204条,平均为178条,引物的多态性信息含量(PIC)范围在0.074 0~0.153 7,平均值为0.125 7。
2)根据遗传相似系数于0.450 0处划分为2个类群,在遗传相似系数0.460 0处将类群Ⅰ划分2个亚群,又在遗传相似系数0.480 0处将类群Ⅱ划分2个亚群,并将保持系且主要来源于中国东北的材料划分为类群Ⅰ,恢复系且主要来源于美国的材料划分为类群Ⅱ。
3)通过对10个已审定强优势杂交种的13个亲本进行分析发现,杂交种亲本间的遗传相似性多分布在0.329 2~0.637 6;其中类群Ⅰ的中国东北材料与类群Ⅱ的国外材料之间杂交配制的组合存在较强的杂种优势。
4)对不同群内亲本材料的遗传多样性分析发现,类群I与Ⅱ的遗传多样性相似度较高,4个亚群中的亚群Ⅰ-1和Ⅱ-2遗传多样性高于亚群Ⅰ-2和Ⅱ-1;通过主坐标分析,同样将亲本划分为2个类群,验证了聚类分析结果,且类群Ⅰ的2个亚群存在较为丰富的遗传多样性。
