乙氧氟草醚对桂味荔枝冬梢控杀效果及内源激素影响的研究
2023-09-07邱燕萍袁沛元
金 峰 向 旭 邱燕萍 袁沛元 凡 超
(广东省农业科学院果树研究所/农业农村部南亚热带果树生物学与遗传资源利用重点实验室 / 广东省热带亚热带果树研究重点实验室,广东 广州 510640)
荔枝(Litchi chinensisSonn.)为无患子科(Spindaceae)常绿乔木,在广东、广西、海南和福建广泛种植[1]。荔枝冬梢生长消耗树体营养,如不严格管理嫩梢,则对开花、坐果率及产量影响较大[2-3]。桂味荔枝是我国种植面积前十的主栽荔枝品种,其控梢成花问题是业内公认的难题。已报道的控梢剂主要有百草枯、乙烯利、多效唑和乙氧氟草醚[4]。百草枯作为一种高毒农药,在市场上受到严格管控[5],现已停止使用。乙烯利和多效唑处理荔枝冬梢时易受环境和树体情况等因素影响,使用不当会对树体周边的成熟叶片造成伤害[6-7]。乙氧氟草醚是一种使用广泛的二苯醚类除草剂[8],具有高效、低毒和低残留特点[9-11],目前在荔枝栽培中被用于控杀冬梢。
乙氧氟草醚是一种抑制植物色素合成的强触杀性除草剂[12-15]。在植物中,在分子氧存在的条件下,原卟啉原氧化酶(protoporphyrinogen oxidase,PROTOX)催化原卟啉原IX 生成原卟啉IX[16]。在光和氧的作用下,原卟啉IX 产生大量单线态氧,这些单线态氧可以氧化官能团,使光合作用单元停止工作[17-18],也可以通过加速细胞膜的氧化引起植物细胞受损[19-20]。单线态氧信号途径可以与乙烯、水杨酸和茉莉酸等信号分子交叉连接,形成信号网络[21-22]。研究表明,乙烯、茉莉酸和水杨酸信号途径不仅是植物抵御生物和非生物胁迫的重要途径,而且在一定条件下还会参与植物细胞凋亡信号的传递[23-24]。拟南芥flu突变体在黑暗条件下处理15 h 后被转移至光照条件,其所产生的大量单态氧导致植株死亡,这一过程被证实与乙烯、茉莉酸和水杨酸激素信号途径被激活有关[25-26]。植物内源乙烯是以甲硫氨酸为底物,依次经过腺苷甲硫氨酸合成酶、1-氨基环丙烷羧酸(1-aminocyclopropane-1-carboxylic acid,ACC)合成酶和ACC 氧化酶三步催化而成[27]。ERF1基因是乙烯信号传导途径中重要的应答因子之一,在植物乙烯的信号传导途径中被EIN3/EIL1 蛋白激活[28-31]。LOX、AOS和AOC基因是植物茉莉酸生物合成途径中的关键基因,茉莉酸从硬脂酸合成途径中合成[32-35]。有研究表明,EDS1 和PAD4 是水杨酸介导的系统诱导抗病(systemic acquired resistance,SAR)反应中传递信号的重要基因[36]。乙烯、茉莉酸和水杨酸介导的防御系统形成代谢网络交叉互作,水杨酸信号主要在防御早期发挥作用,而乙烯和茉莉酸信号主要参与后期抗逆的建立[37-38]。
本研究以桂味荔枝为材料,利用不同浓度乙氧氟草醚处理其冬梢,在组织和细胞水平上对冬梢进行形态和结构观察,筛选出最佳处理浓度。并对水处理和乙氧氟草醚处理的冬梢进行比较转录组研究,检测主要内源激素的含量变化,同时对胁迫相关激素信号途径关键基因的表达量进行验证。旨在初步了解乙氧氟草醚控梢的分子机理。
1 材料与方法
1.1 试验材料
试验荔枝样本种植于广东省农业科学院荔枝试验园(113.37848°E,23.15922°N),取自20 年左右树龄的桂味荔枝。试验前,选择生长良好、树势相近的荔枝树作为样株(n=21)。每次取样时,用锋利刀片从荔枝果树上分别切下长度约为5~8 cm 的荔枝嫩梢,同时采集对应嫩梢下的成熟叶片(3 片),随后立即用固定液或液氮保存并带回实验室用于进一步的观察和检测。脱色液配制:将10 g 水合氯醛溶解于10 mL 水中配成2.5 g·mL-1水合氯醛溶液。台盼蓝染色剂配制:将9 mL 甲醇、3 mL 水饱和酚、3 mL 甘油、3 mL 乳酸混合后加入3 mg台盼蓝配制而成[39]。
1.2 仪器设备与试验试剂
乙氧氟草醚、百草枯,广州佳途科技有限公司;乙腈,德国Fisher Chemica 公司;RN40 RNA 提取试剂盒,北京艾德莱生物科技有限公司;ZH120 多糖多酚植物RNA 提取试剂盒,北京华越洋生物科技公司;Nanodrop 2000 超微量紫外可见分光光度计、ABI7500 fast Real-Time PCR system,美国Thermo Fisher 公司;2100 芯片实验室GX,安捷伦科技(中国)有限公司;Tissuelyser-48研磨仪,中国净信公司;SHIMADZUCBM 30A 超高效液相色谱,日本岛津公司;4500QTRAP 串联质谱,美国AppliedBiosystems公司;SteREO Discovery V20体视镜,德国蔡司公司;EcLipse 80i显微镜,日本尼康株式会社;Novaseq 6000 测序仪,美国因美的公司;BE-110四维旋转仪,上海量壹科学有限公司。
1.3 试验方法
1.3.1 乙氧氟草醚和百草枯处理 配制浓度为40、60、80、100、120 μmol·L-1乙氧氟草醚和50 μmol·L-1百草枯,选择无风、晴朗、干燥的早晨,以水处理为对照,利用喷雾器对11月末到12月初有新的嫩梢抽出、长势相近的桂味荔枝果树自上而下均匀喷雾,至叶片均匀喷湿为止,剩余药液切忌反复喷施。样品组织处理时间及试验用途等见表1。

表1 试验材料及其处理方式和用途Table 1 Experimental materials and their preservation process and uses
台盼蓝染色具体步骤:将80 μmol·L-1乙氧氟草醚和50 μmol·L-1百草枯处理20 h 后的嫩梢的叶片剪下,置于5 mL离心管中,于试管中加入2 mL台盼蓝染色剂以浸没叶片,再将试管置于水浴锅中开盖沸煮2 min,利用脱色液将叶片材料背景洗净,置于体视镜及显微镜下拍照,组织死细胞将会被染成蓝色斑。
1.3.2 总RNA 提取和转录组建库及测序 使用CLB+RN40 RNA 提取试剂盒、CTAB+RN40 RNA 提取试剂盒进行测序。使用2100 芯片实验室GX 对RNA完整性进行检测。将mRNA Capture Beads置于四维旋转仪上充分混匀,平衡30 min 后建库。对于构建好的文库使用Novaseq 6000进行上机测序。
1.3.3 荔枝数据处理、拼装和激素信号途径相关基因的检索 对获得的原始读数(raw reads)进行过滤,去除带接头(adapter)的、含N 的和低质量的reads,对干净读数(clean reads)进行Q20 和Q30 含量计算。使用HISAT2 v2.1.0 将配对末端clean reads 与参考基因组比对。使用feature Count(1.5.0-p3)计算映射到每个基因的读数,然后根据基因的长度计算每个基因每千个碱基的转录每百万映射读取的片段。
1.3.4 植物激素色谱与质谱联用含量检测流程 取桂味荔枝冬梢嫩梢鲜样,加入液氮,利用研磨仪研磨(50 Hz,1 min)至粉末状。称量(100±1) mg 鲜样粉末,加入1 mL 提取液(甲醇∶水∶甲酸=75∶23∶2),室温下振荡15 min,离心(12 000 r·min-1,4 ℃,10 min)后,取上清于新离心管中。向残渣中加入0.5 mL 提取液,重复振荡和离心,合并上清,4 ℃浓缩仪中浓缩干,加入200 μL 复溶液(甲醇∶水=4∶1)复溶,离心(转速12 000 r·min-1,10 min)后,吸取上清,用0.22 μm 微孔滤膜过滤样品,并保存于进样瓶中,用于液相色谱串联质谱分析。利用超高效液相色谱和串联质谱采集数据,基于代谢物标准品质谱分析的离子对与保留时间等信息对目标代谢物进行定性分析。根据代谢物保留时间与峰型信息,对每个代谢物在不同样本中检测到的质谱峰进行校正,确保定性定量的准确。利用标样制取不同浓度的标准品溶液,获取各个浓度标准品对应定量信号的质谱峰强度数据:以标准品浓度为横坐标,质谱峰的峰面积为纵坐标,绘制不同代谢物的标准曲线。将检测到的所有样本中目标代谢物的积分峰面积代入标准曲线线性方程进行计算,最终得到实际样本中目标代谢物的绝对含量数据。
1.3.5 总RNA 提取和cDNA 反转录 选用ZH120 RNA 提取试剂盒,按照说明书提取荔枝总RNA,利用Nanodrop 2000 超微量紫外可见分光光度计测定总RNA 浓度及纯度。为验证乙氧氟草醚处理荔枝冬梢过程中植物细胞凋亡相关的激素途径(乙烯、茉莉酸和水杨酸途径)的关键基因的表达模式,利用qRT-PCR技术对处理后10、20 和36 h 这3 个时间点的15 个关键基因进行验证。mRNA 第一链cDNA 合成使用2 μg RNA和北京全式金生物TransScript®mRNA RT酶混合系统,使用Nanodrop 2000超微量紫外可见分光光度计对提取的核酸进行浓度检测。利用常规qRT-PCR 对差异表达基因进行验证。cDNA 合成使用2 μg RNA、基因组DNA 清除剂和TransScript®- Uni RT/RI 酶混合系统。
1.3.6 qRT-PCR检测 qRT-PCR均使用PerfectStart®Green qPCR Super Mix 系统,在ABI7500 fast Real-Time PCR system 仪器上操作。为了获取荔枝基因组中乙烯、茉莉酸和水杨酸信号途径相关基因序列,从拟南芥信息资源(theArabidopsisinformation resource,TAIR)数据库(http://www. arabidopsis. org/)中查找乙烯生物合成关键基因(AtACS1)、信号途径关键基因(AtERF1)、茉莉酸生物合成关键基因(AtLOX1、AtAOS1、AtAOC1和AtOPR3)、水杨酸生物合成关键基因(AtEDS1和AtPAD4)的核酸序列,然后在无患子科基因组数据库(http://www.sapindaceae.com)中比对搜索,获得荔枝中对应的同源基因。最后选取同源性高的15 个同源基因作为候选基因进行表达量的检测,包括:2个1-氨基环丙烷-1-羧酸合成酶(1-aminocyclopropane-1-carboxylate synthase,ACS)基因(LITCHI025775和LITCHI022037),4 个乙烯反应因子(ethylene response factor,ERF)基因(LITCHI026054、LITCHI015333、LITCHI003587和LITCHI015335),2 个脂氧合酶(lipoxygenase,LOX)基因(LITCHI003969和LITCHI004066),1 个丙二烯氧化物合成酶(allene oxide synthase,AOS)基因(LITCHI011121),1个氧化烯环化酶(allene oxide cyclase,AOC)基因(LITCHI005484),2 个氧化植物二烯酸还原酶(oxo-phytodienoic acid reductase,OPR)基因(LITCHI011707和LITCHI017668),1 个疾病增强易感性(enhanced disease susceptibility,EDS)基 因(LITCHI023136),2 个植物抗毒素缺陷(phytoalexin deficient,PAD)基因(LITCHI003725和LITCHI016561)。利用Primer Premier 5 软件设计这些基因的qRT-PCR 引物(表2)。反应体系共20 μL:2×SYBR Premix Ex Taq(日本TaKaRa公司)10 μL,10 μmol·L-1上、下游引物各0.4 μL,cDNA 模板2 μL,ROX Reference Dye 0.4 μL,加ddH2O 至20 μL。反应程序(两步法):95 ℃预变性30 s,95 ℃变性5 s,60 ℃退火、延伸30 s,40 个循环;每个样品3 次生物学重复。使用2-△△Ct法对数据进行处理。
现代物流不仅指原材料、产品等从生产到消费的全程实物流动,还包括伴随物流活动过程中的物流信息交流,而且,现代物流在使用信息技术以及网络技术,将以往分离的物流、商流、信息流和运输、采购、代理、仓储、配送等环节紧密联系起来,成为了一条完整的供应链。因此可以讲,现代物流又是信息流、货物流、资金流和人才流的统一。

表2 目标基因的qRT-PCR引物相关信息Table 2 Information about qRT-PCR primers of the target genes
1.4 数据处理
计算校正内源激素含量及实时荧光定量试验结果的平均值和标准误差,采用SPSS 22.0 软件进行数据统计分析,通过方差分析及P检验比较数据间的差异。
2 结果与分析
2.1 乙氧氟草醚浓度梯度的筛选
由图1 可知,水处理的冬梢在整个试验过程中一直保持绿色状态。10 h 时,各浓度处理的荔枝冬梢嫩叶和嫩梢没有明显的萎蔫,或只有零星水渍状斑点。20 h 时,40、100 和120 μmol·L-1浓度乙氧氟草醚处理的嫩叶出现轻度萎蔫、轻微卷缩或皱褶,或有少量水渍状斑点,嫩梢生长受抑制,不弯曲;而60和80 μmol·L-1浓度乙氧氟草醚处理的嫩叶明显褐化、萎蔫、卷曲,有较多水渍状斑块转为灼烧状斑块,嫩梢卷曲或有大量斑点。36 h 时,40 μmol·L-1浓度乙氧氟草醚处理的嫩叶有少量水渍状斑点,嫩梢生长受抑制,嫩叶微卷曲;60 μmol·L-1浓度乙氧氟草醚处理的嫩叶高度褐化、萎蔫,严重卷曲甚至凋落,有较大面积灼烧;80、100 和120 μmol·L-1浓度乙氧氟草醚处理的嫩叶严重褐化、萎蔫,干枯或凋落,嫩梢严重褐化、卷曲甚至干枯。

图1 对照、乙氧氟草醚处理后0、10、20、36 h时的荔枝冬梢形态变化Fig.1 Morphological changes of litchi winter shoots at 0, 10, 20 and 36 h after treatment with control and oxyfluorfen
控梢反应时间显示,60和80 μmol·L-1浓度乙氧氟草醚处理的反应时间较40、100和120 μmol·L-1浓度的时间早,在20 h 即可看到明显的褐化表型。控梢效果显示,40 μmol·L-1浓度处理的效果不足以抑制荔枝冬梢的生长,60 μmol·L-1浓度处理在36 h 时的杀梢效果较更高浓度较差,80、100和120 μmol·L-1浓度乙氧氟草醚处理的荔枝冬梢在36 h之后均可达到较理想的抑制效果(图1)。综合处理时间和抑制效果,80 μmol·L-1乙氧氟草醚处理是抑制5~8 cm 荔枝冬梢生长最有效的处理浓度。
2.2 乙氧氟草醚与百草枯处理对荔枝冬梢的抑制效果
由图2 可知,50 μmol·L-1百草枯处理荔枝冬梢在20 h时嫩叶有较多灼烧状斑块,明显萎蔫卷曲,嫩梢卷曲或有大量斑点;36 h时嫩叶严重褐化,嫩梢严重褐化干枯。与50 μmol·L-1百草枯处理相比,80 μmol·L-1以上浓度的乙氧氟草醚处理均可达到相近的效果(图1、2),可以有效抑制荔枝冬梢生长,使其褐化死亡。

图2 对照、乙氧氟草醚和百草枯处理后20、36 h时的荔枝冬梢形态变化Fig.2 Winter shoots in Litchi treated with oxyfluorfen,paraquat and control, respectively, changed in color after 20 and 36 hours
细胞膜受损死亡的细胞可以被台盼蓝染液染色。观察用台盼蓝处理20 h 后的3 组荔枝冬梢细胞的染色结果,发现被80 μmol·L-1乙氧氟草醚和50 μmol·L-1百草枯处理后的荔枝冬梢细胞均能着色,而对照组的离体嫩梢细胞几乎未被染色(图3)。说明80 μmol·L-1乙氧氟草醚和50 μmol·L-1百草枯处理20 h的荔枝冬梢细胞膜受损,部分细胞已经死亡,该处理可有效杀死冬梢。
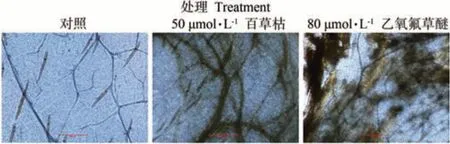
图3 台盼蓝分别被水(CK)、乙氧氟草醚和百草枯处理20 h后的荔枝冬梢细胞的染色情况Fig.3 Trypan blue staining of Litchi winter shoot cells treated with H2O(CK), oxyfluorfen and paraquat for 20 h, respectively
由图4可知,被80 μmol·L-1乙氧氟草醚和50 μmol·L-1百草枯处理72 h后,无切口的成熟叶片(包括完整叶片的内部组织和边缘组织)仍然保持与对照组相似的绿色。对有切口的成熟叶片,相比水处理的对照组,百草枯处理能够明显造成靠近切口附近的多层细胞的死亡,而乙氧氟草醚处理切口附近1~2 层组织损伤。因此,在无伤口的成熟叶片正常使用上述两种试剂无明显毒害作用,但对有伤口的叶片,乙氧氟草醚的伤害性更弱。

图4 显微镜下,对照、50 μmol·L-1百草枯和80 μmol·L-1乙氧氟草醚处理72 h后不同状态成熟叶片组织的情况Fig.4 Photograph from microscope about the tissue in three different position of mature leaves of Litchi tree from treated with H2O (CK), 50 μmol·L-1 paraquat and 80 μmol·L-1 oxyfluorfen for 72 h
2.3 乙氧氟草醚处理荔枝冬梢转录组学研究结果
为了进一步分析差异基因(differential expression gene, DEG)的生物学功能,采用京都基因和基因组大百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)数据库对DEGs 进行通路富集分析。在处理组中,4 482 个差异基因涉及57 个KEGG 途径,包括细胞过程、遗传信息处理、环境信息处理、新陈代谢、有机体系统等5 个方面,大部分途径属于信号转导类别。差异基因富集的代谢通路有植物激素信号转导途径、植物与病原菌互作、丝裂原活化蛋白激酶(mitogenactivated protein kinase,MAPK)信号通路、植物和淀粉蔗糖代谢途径(图5-A),分别涉及118、114、68和67个相关基因,聚类基因数最多的KEGG 途径是植物激素信号转导信号途径。Mapman分析结果显示,激素信号途径中,与胁迫反应相关的乙烯、茉莉酸和水杨酸途径中有大量的基因差异表达(图5-B)。
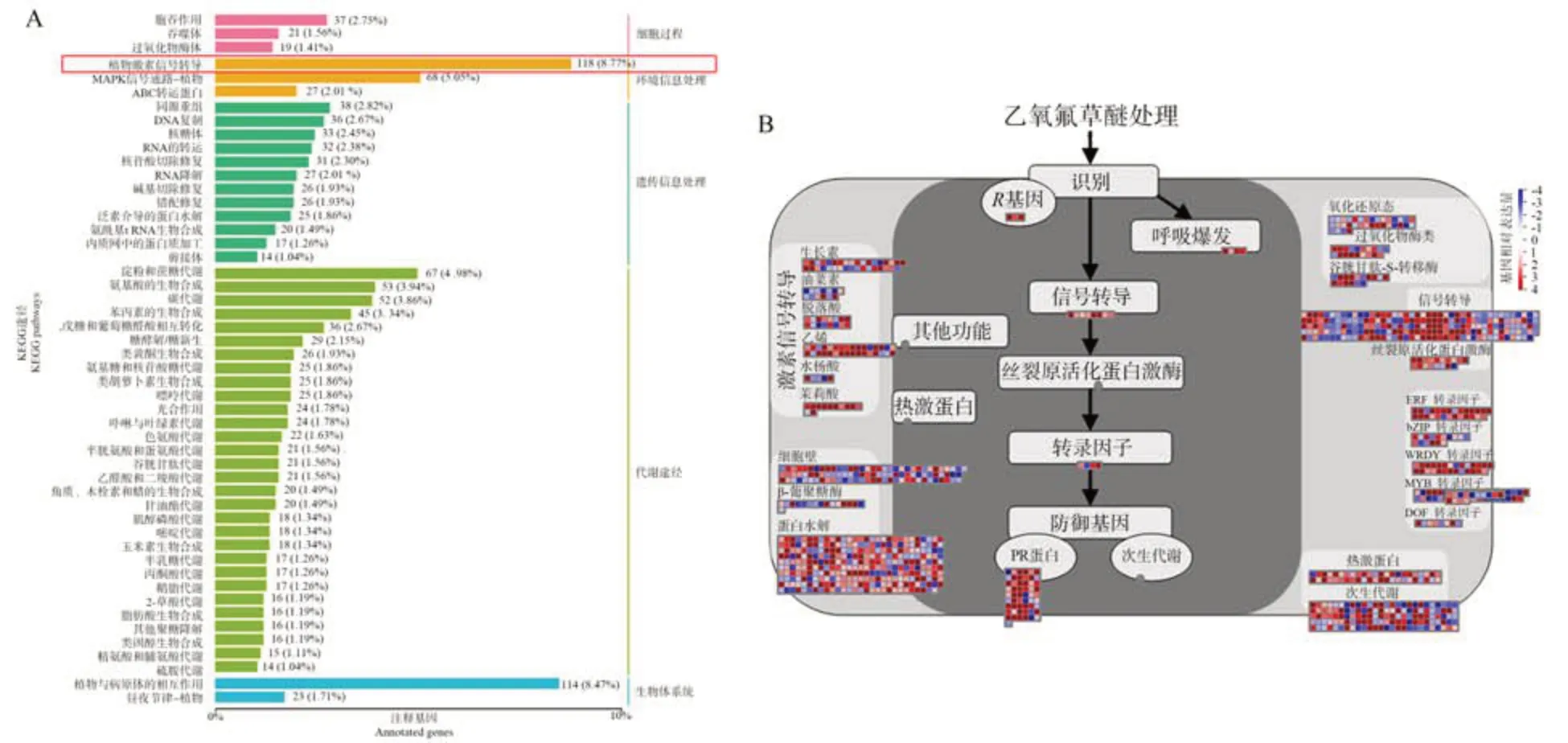
图5 乙氧氟草醚处理荔枝冬梢中差异基因最富集的KEGG途径和Mapman胁迫处理后富集途径Fig.5 The enriched KEGG and Mapman pathways between the DEGs identified in litchi winter shoots treatment with control and oxyfluorfen
2.4 乙氧氟草醚处理荔枝冬梢激素含量测定结果
转录组学结果显示(图5),显著差异基因富集代谢通路中,基因数量最多的是植物激素信号转导途径,推测激素可能在乙氧氟草醚抑制的荔枝冬梢生长过程中起到关键性的作用。利用超高效液相色谱和串联质谱技术对水处理和80 μmol·L-1乙氧氟草醚处理荔枝冬梢样品中5 大类激素(生长素、细胞分裂素、脱落酸、水杨酸、茉莉酸)中20 种激素物质和乙烯前体ACC 的含量进行测定。结果显示(表3),生长素和细胞分裂素类激素中各种物质的含量变化趋势不一致,其中吲哚-3-乙酸、吲哚-3-甲醛、吲哚-3-乳酸和吲哚-3-乙酸甲酯含量在乙氧氟草醚处理后较对照升高,吲哚乙酸-天冬氨酸和N-(3-吲哚乙酰基)-L-苯丙氨酸含量则降低。细胞分裂素中顺式-玉米素-D-核糖甙、双氢玉米素核苷、反式玉米素和玉米素核苷的含量在处理后增加,而6-苄氨基嘌呤、N6-异戊烯腺嘌呤、异戊烯腺嘌呤核苷和反式-玉米素-9-Β-葡萄糖苷的含量在处理后下降。1-氨基环丙烷羧酸、茉莉酸类(二氢茉莉酸、茉莉酸、茉莉酸-异亮氨酸和茉莉酸甲酯)和水杨酸类(水杨酸和水杨酸-2-O-β-葡萄糖苷)激素物质含量在乙氧氟草醚处理后均呈增加趋势。
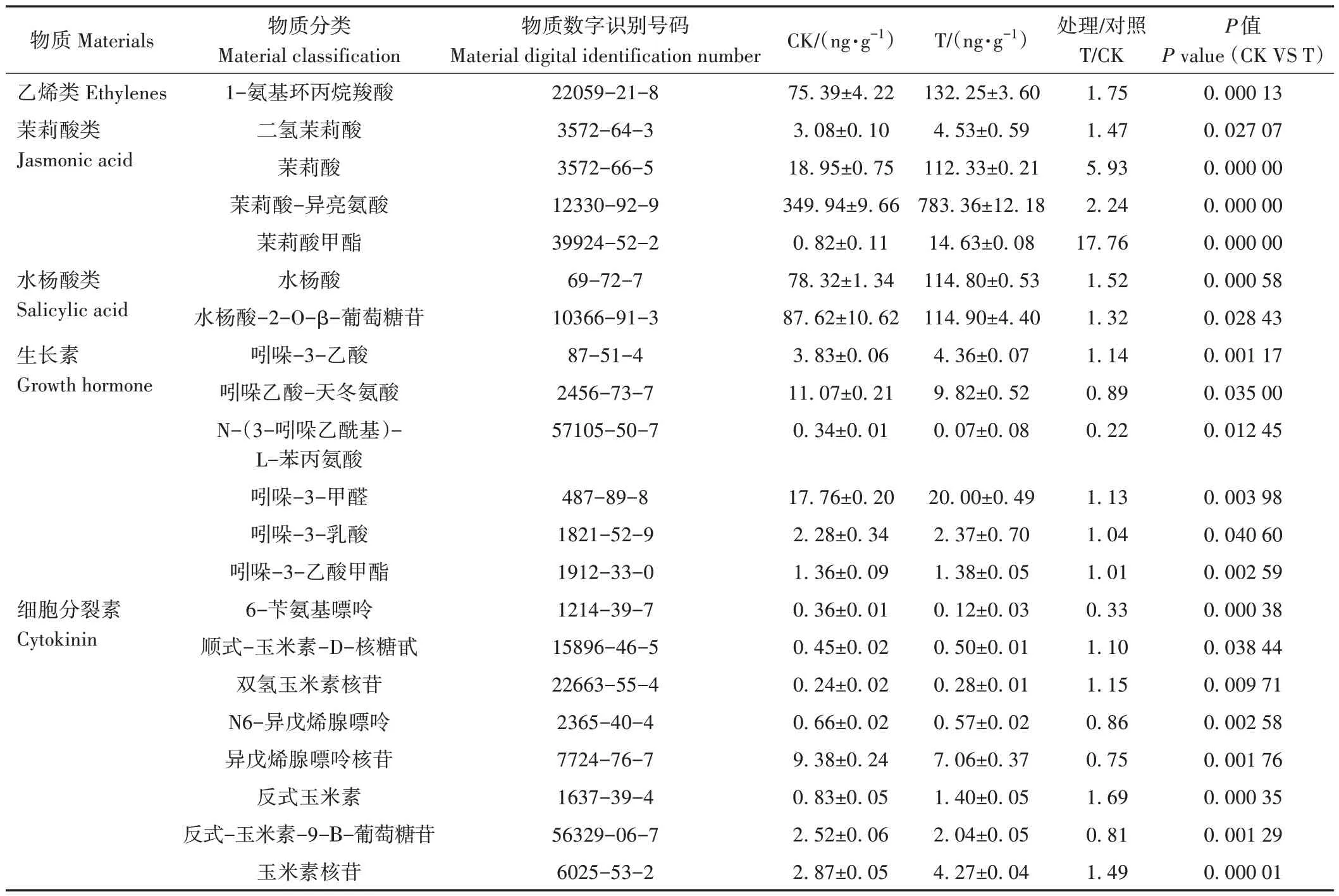
表3 水(CK)和80 μmol·L-1乙氧氟草醚(T)处理20 h后的荔枝冬梢激素含量变化Table 3 Changes of hormone concentration in litchi winter shoots after a 20-hour treatment with H2O (CK) and 80 μmol·L-1 oxyfluorfen (T)
2.5 荔枝冬梢激素信号途径相关基因相对表达量对乙氧氟草醚的响应特性
为验证转录组测序数据的可靠性,从转录组鉴定到的差异基因中选取了乙烯、茉莉酸和水杨酸途径中15 个可能参与乙氧氟草醚处理抑制荔枝冬梢过程的基因进行qRT-PCR 分析。结果显示,qRT-PCR 分析的基因表达模式与RNA-Seq 方法分析的基因表达模式相似,证实了RNA-Seq数据的可靠性。
为了研究乙烯、茉莉酸和水杨酸在乙氧氟草醚处理抑制荔枝冬梢过程中作用的分子机制,测定了2 个ACSs基因、4 个ERFs基因、2 个LOXs基因、1 个AOS基因、1 个AOC基因、2 个OPR基因、1 个EDS基因、2 个PADs基因在80 μmol·L-1乙氧氟草醚处理后的表达量变化情况。结果显示(图6),当处理时间达到10 h 时,乙氧氟草醚处理后荔枝嫩梢中的乙烯、茉莉酸和水杨酸途径基因的相对表达量与对照整体无显著差异。当处理后时间达到20 h 时,所有检测基因的相对表达量与对照相比均出现不同程度的显著上调。当处理后时间延长至36 h 时,大部分检测基因相对表达量较20 h时有不同程度的下调,但乙烯途径基因LITCHI022037和茉莉酸途径基因LITCHI003969和LITCHI011121相对表达量仍呈现上升趋势。
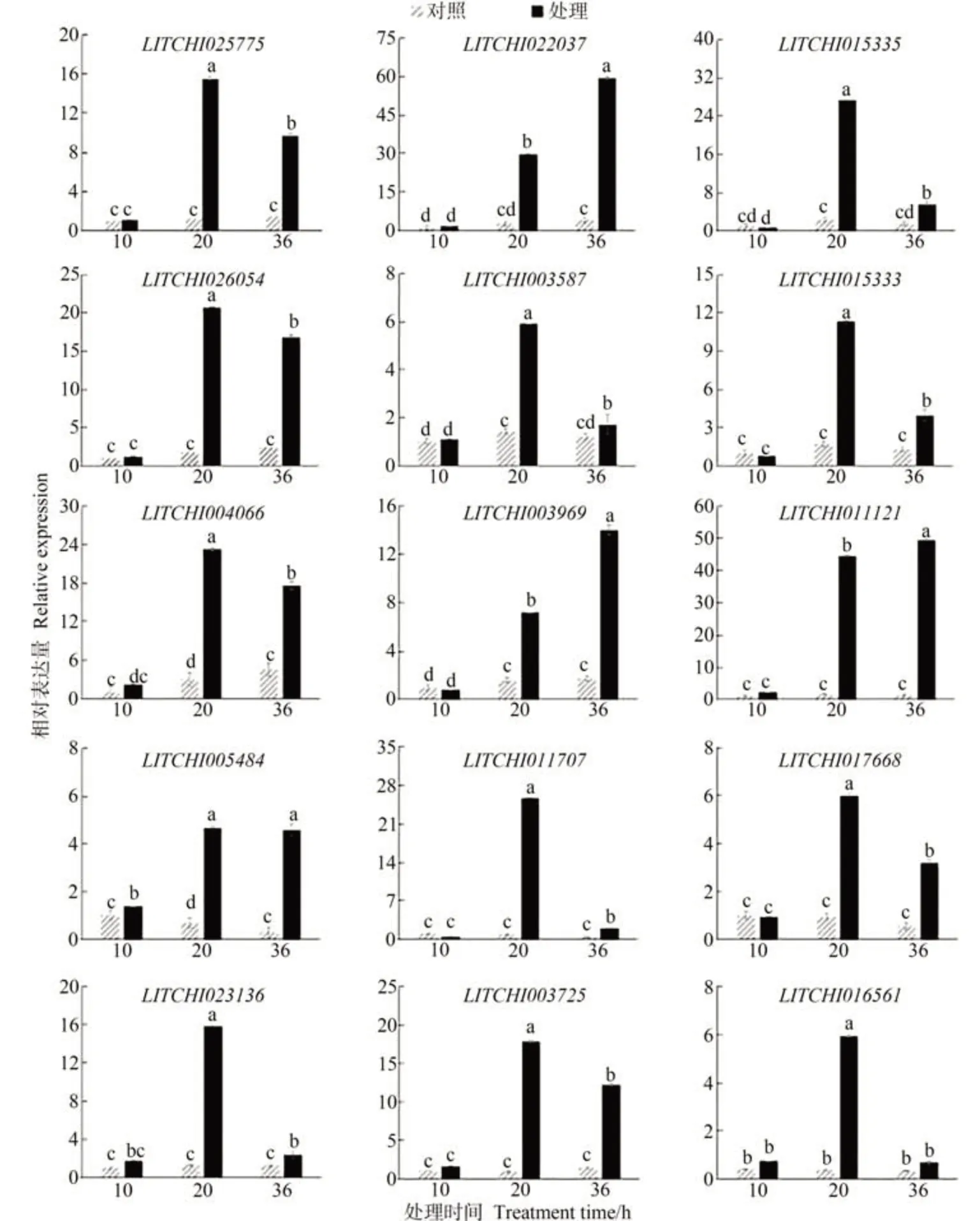
图6 荔枝嫩梢中乙烯、茉莉酸和水杨酸信号途径内各基因的相对表达量对乙氧氟草醚处理的响应特性Fig.6 The characteristics of the genes responding to oxyfluorfen treatment in ethylene, jasmonic acid and salicylic acid signaling pathways of the winter shoots in Litchi
3 讨论
二苯醚类除草剂在荔枝控梢上的应用仍不广泛,本研究验证了不同浓度乙氧氟草醚对荔枝嫩梢的杀伤效果,发现80 μmol·L-1乙氧氟草醚处理控制荔枝冬梢生长的综合效果最好,低于80 μmol·L-1则后期作用效果较弱,而更高浓度处理在前36 h 的反应时间相较于80 μmol·L-1更长。乙氧氟草醚处理后的荔枝冬梢叶片细胞质内产生1O2,这些1O2能够引起多不饱和脂肪酸的脂质过氧化作用,进而破坏生物膜的完整性[17]。台盼蓝染色结果也证明80 μmol·L-1乙氧氟草醚处理可以使荔枝嫩梢细胞膜破损,细胞出现死亡。对成熟荔枝叶片处理发现,利用80 μmol·L-1乙氧氟草醚和50 μmol·L-1百草枯处理荔枝成熟叶片72 h 后,无切口的包括完整叶片的内部组织仍然保持与对照组相似的绿色。而对有伤口的叶片,80 μmol·L-1乙氧氟草醚伤害性更弱,使用更加安全。并通过对比50 μmol·L-1百草枯的使用效果,发现80 μmol·L-1乙氧氟草醚对冬梢嫩叶和嫩梢的杀梢效果相近,且具有对成熟叶损伤部位细胞损伤较小的优点。前人在脐橙树苗和沙糖橘嫩夏梢杀梢效果的研究中也发现,乙氧氟草醚对柑橘成熟叶片和成熟期果实无药害[40-41]。以上结论为深入研究二本醚类除草剂控梢效果和开发以二苯醚类除草剂为基础的新型荔枝杀梢剂提供了一定的参考依据。
利用转录组学比较分析水处理和80 μmol·L-1乙氧氟草醚处理的荔枝冬梢样品发现,显著差异基因主要富集在植物激素信号转导、植物与病原菌互作、MAPK 信号通路等代谢途径。说明乙氧氟草醚可引起荔枝冬梢中植物激素代谢途径变化。因此,利用超高效液相色谱和串联质谱技术测定对照和乙氧氟草醚处理20 h后的荔枝冬梢样品激素或前体含量。结果显示6 大类激素中,茉莉酸、水杨酸和乙烯前体ACC 的含量在处理后呈上升趋势,说明这三种物质在荔枝冬梢中响应乙氧氟草醚处理。
为验证转录组学和激素含量测定的结果,利用qRTPCR技术对荔枝冬梢对照和不同处理时间样品进行检测,结果显示,使用80 μmol·L-1乙氧氟草醚处理荔枝嫩梢后10 h 时还未明显激活茉莉酸、乙烯和水杨酸信号途径基因,20 h 时所有关键基因相对表达量显著提高,但在处理36 h后激活效应有所降低。当处理后时间延长至36 h时,大部分基因表达量水平有不同程度的下调,仅有乙烯途径基因LITCHI022037及茉莉酸途径基因LITCHI003969和LITCHI011121相对表达量仍呈现上升趋势。水杨酸信号基因LITCHI023136、LITCHI003725和LITCHI016561相对表达量在20 h 时较10 h 明显提高,而乙烯和茉莉酸基因LITCHI025775、LITCHI026054、LITCHI015333、LITCHI003587、LITCHI015335、LITCHI004066、LITCHI005484、LITCHI011707和LITCHI017668不 仅 在20 h 时表达量明显提高,在处理36 h 仍有部分基因(LITCHI022037、LITCHI003969和LITCHI011121)的表达量较20 h升高。基因表达趋势结果与表型结果相吻合,说明这三种激素信号基因在乙氧氟草醚处理荔枝冬梢时通过胁迫响应及介导细胞程序性死亡途径参与到抑制荔枝冬梢生长的过程。其中乙稀和茉莉酸信号基因在处理36 h 时仍有显著升高的结果,与前人研究结果一致[37-38]。近年来,采用生化药物控制荔枝嫩梢的技术在不断完善,主要通过尝试不同药物、时间、计量来总结在施用过程中的适用比例[42-43]。本研究利用乙氧氟草醚处理后,荔枝嫩梢内生成的单线态氧会造成荔枝冬梢细胞膜的损伤。同时,单线态氧可能作为信号诱导乙烯、水杨酸盐和茉莉酸代谢,导致荔枝冬梢生长被抑制并最终死亡。
4 结论
本研究发现,80 μmol·L-1乙氧氟草醚处理控杀桂味荔枝冬梢效果最好。利用台盼蓝染色证明80 μmol·L-1乙氧氟草醚处理可使荔枝冬梢细胞膜受损并导致细胞死亡。与水处理相比,80 μmol·L-1乙氧氟草醚处理后的荔枝冬梢中乙烯、茉莉酸和水杨酸类激素物质的含量在处理后20 h 增加。由于荔枝生长周期长,且再生体系技术尚未成熟,一些激素信号途径关键基因并没有得到确证,这使得荔枝激素调控分子机制的研究难以顺利开展。本研究对乙氧氟草醚处理后荔枝冬梢中与单线态氧信号途径互相关联的乙烯、水杨酸盐和茉莉酸信号、代谢途径关键基因在10、20 和36 h 的相对表达量进行了检测,变化趋势与转录组学结果一致,说明植物激素及其相关基因在乙氧氟草醚控制荔枝冬梢生长中可能发挥着重要作用。
