不同生殖生长阶段高温胁迫对水稻产量和品质的影响
2023-09-07叶世河刘金龙匡佳丽莎杨洪涛刘晓龙
季 平 柳 浩 叶世河 刘金龙 匡佳丽 龙 莎杨洪涛,2 刘晓龙,2,*
(1宜春学院生命科学与资源环境学院,江西 宜春 336000;2江西省高等学校硒农业工程技术研究中心,江西 宜春 336000)
随着全球气候变暖趋势的加剧,高温热害已成为限制水稻生产的重要逆境因子之一。据统计,全球平均气温每升高1 ℃,水稻产量将下降3.2%[1]。长江中下游地区是我国水稻主产区,承担了全国约30%的水稻产量[2],该地区的水稻种植大部分时间都处于炎热的夏季,水稻育秧、移栽成活、营养生长及生殖生长都会遭受不同程度的高温胁迫[3]。高温胁迫下,水稻的种子发芽和幼苗生长均受到抑制[4-5];同时,早稻的孕穗、抽穗开花和籽粒灌浆,以及晚稻的移栽成活和分蘖也受到高温胁迫的不利影响,造成结实率和千粒重的下降,导致水稻减产[6]。
水稻在生殖生长阶段对高温胁迫最敏感,此时的高温胁迫通过多种途径影响水稻产量形成和稻米品质[7]。水稻的产量形成包括拔节孕穗、抽穗、颖花开放、胚乳形成和淀粉合成等一系列复杂的生理代谢过程,其中任何一个生理过程遭受高温胁迫都会影响产量形成[8]。水稻在拔节孕穗期遭受高温胁迫将会降低光合效率,影响干物质积累和幼穗发育[9]。抽穗开花期是水稻籽粒形成的关键时期,此时遭遇高温胁迫将会导致花粉败育、受精不良,降低穗部颖花数和结实率[10]。水稻产量的高低与籽粒充实程度密切相关,高温胁迫会导致灌浆结实期的水稻灌浆不足,降低千粒重,造成水稻减产[7,11]。此外,高温胁迫会影响结实期水稻的淀粉合成速率,改变淀粉结构,造成稻米品质下降[9,11]。因此,研究水稻在不同生殖生长阶段生长发育、产量形成和稻米品质对高温胁迫的响应规律,对于探索水稻的耐热机制具有重要意义。
前人在水稻产量形成对高温胁迫的响应机制研究中,多为关注单个生育期高温胁迫对水稻产量的影响及分子机制[12-14],如王强等[15]研究了水稻结实率和产量对不同时期高温胁迫的响应;宋有金等[16]研究了产量和根系伤流液强度对不同时期高温胁迫的响应。然而,有关不同时期高温胁迫对水稻产量损失和品质的影响,以及水稻产量结构和稻米品质对不同生殖生长阶段高温胁迫响应的研究鲜有报道。综合评估不同时段高温胁迫对水稻产量和稻米品质的影响机制有助于全面认识水稻的耐热机理。因此,本研究通过精确控制人工气候室温度,在不同生殖生长阶段模拟高温胁迫,研究高温胁迫对水稻产量结构和稻米品质的影响,旨在为水稻高温热害的产量评估和探索提高水稻品质的途径提供理论依据。
1 材料与方法
1.1 试验材料
以长江中下游地区普遍推广种植的常规籼稻品种黄华占(HHZ)和杂交籼稻品种Y两优1577(YLY1577)为试验材料,生育期均为136 d,由江西省高等学校硒农业工程技术研究中心提供。
1.2 水稻种植方式
试验于2021 年5—9 月在宜春学院现代农业研究所和江西省高等学校硒农业工程技术研究中心实验室进行,水稻的前期培养在现代农业研究所采用盆栽方式进行。5 月1 日播种,供试水稻种子用0.1%(v/v)的HgCl2充分消毒10 min,蒸馏水冲洗干净后在30 ℃恒温箱中浸种48 h,清洗干净后于培养皿中进行催芽。选取出芽一致的种子播种于装有草炭土(有机质30.91 g·kg-1、全氮2.30 g·kg-1、速效磷20.56 mg·kg-1、速效钾142.39 mg·kg-1)的育秧盘中进行育秧,育秧25 d后移栽。选择均匀一致的壮苗移栽至高28 cm、内径25 cm 的塑料桶中,塑料桶内装有稻田土,采自宜春学院现代农业研究所水稻田(有机质26.32 g·kg-1、全 氮1.93 g·kg-1、速 效 磷15.25 mg·kg-1、速 效 钾106.85 mg·kg-1),土壤经粉碎、过筛、风干并混匀后装入桶中,每桶装土10 kg。参照前人的施肥方式,秧苗移栽前施底肥4.5 g,底肥为复合肥(氮磷钾养分含量均为15%),在分蘖期(5~5.5 片叶)进行追肥,每盆施3.0 g 尿素[15]。秧苗移栽时每桶2 穴,每穴4 棵苗。移栽后于自然条件下培养,生长期按照常规栽培方式进行管理,及时拔除杂草,并严格防控病虫害。
1.3 高温处理
高温处理参照前期试验方法在江西省高等学校硒农业工程技术研究中心的KSD-15J-3 人工气候室(常州海博仪器设备有限公司)进行,采用远红外热风机(额定功率2 000 W)和自动控温系统调节室温,采用飞利浦白色LED 灯管提供光照[7]。对照组(CK)环境设置为:白天(7∶00~19∶00)32 ℃,夜晚(19∶00~7∶00)26 ℃;高温处理组在每天的9∶00~16∶00 设置38 ℃的高温胁迫,其他时间温度设置为32 ℃,温度上下波动约0.5 ℃。相对湿度设置为80%,光照时间均为12 h光/12 h 暗,光照强度为450 μmol·m-2·s-1,每天早上9∶00和下午16∶00各开门通风10 min。
水稻培养至孕穗期开始进行高温处理,对照组水稻植株从孕穗期至成熟始终置于对照环境。高温处理组设置5 个高温处理时段,分别于始穗期前7 d、始穗期(每盆有3 个分蘖穗抽出剑叶1 cm 左右)、始穗后第8 天、始穗后第15 天和始穗后第22 天移入人工气候室进行高温处理,即以始穗期计为0 d,分别在始穗前7 d至始穗期及始穗后1~7、8~14、15~21、22~28 d 5个时段进行高温处理,分别以HS1、HS2、HS3、HS4 和HS5 表示,每个时段高温处理7 d 后移入对照环境培养至成熟。高温处理时期涵盖孕穗期至灌浆后期。每个品种各处理均设置3次生物学重复,每个生物学重复种植2盆,2 个品种共计72 盆。成熟后采集水稻植株进行考种,收集籽粒用于品质理化指标的测定。
1.4 测定项目与方法
1.4.1 生长发育指标的测定 在成熟期用卷尺测定水稻植株的株高,于根部剪断,自然风干后测定有效穗数,将所有穗剪下后测定茎秆干重,每桶植株随机选取10 个穗(若不足10 个,则以实际穗数为准)测定穗长、穗重、一次枝梗和二次枝梗数。
1.4.2 产量结构指标及产量的测定 将穗脱粒、去杂质(不去空瘪粒),分为实粒和瘪粒,调查每穴的实粒数、瘪粒数、实粒重和瘪粒重,计算总粒数、总粒重、每穗粒数、结实率、千粒重、收获指数和理论产量。其中,收获指数和理论产量按以下公式计算:
1.4.3 籽粒加工和外观品质指标的测定 将稻谷置于室温风干后,按照《GB/T 17891-1999 优质稻谷》[17]的方法测定糙米率、精米率和整精米率,并取精米试样测定长宽比、垩白粒率和垩白度。
1.4.4 稻米胶稠度的测定 按照《GB/T 17891-1999优质稻谷》[17]的方法,取平衡水分后的精米样品5 g,磨碎后过100目样品筛,测定稻米胶稠度。
1.4.5 淀粉黏滞谱特征(rapid viscosity analyzer,RVA)的测定 按照《GB/T 24852-2010 大米及米粉糊化特性测定 快速粘度仪法》[18]的方法测定稻米糊化特性。经水分平衡后的精米磨碎后过100 目样品筛,用HR83-P 水分测定仪(瑞士Mettler Toledo 公司)检测样品水分含量,根据水分含量计算出黏度测定所需样本质量。准确称取样品,加入去离子水,混匀后用RVA Super 4 快速黏度分析仪(澳大利亚Newport Scientific公司)检测稻米黏度[19],并用TCW3 软件进行数据分析。
1.4.6 籽粒理化指标的测定 将不同处理的水稻籽粒脱壳后取精米,磨碎后过100 目样品筛,用于测定淀粉、直链淀粉、支链淀粉、蛋白质、蔗糖、葡萄糖和果糖含量。
淀粉含量的测定:用80%的乙醇将样品中的可溶性糖与淀粉分离,采用酸水解法分解淀粉为葡萄糖,采用蒽酮比色法测定葡萄糖含量,进一步计算淀粉含量[20]。
直链和支链淀粉含量的测定:参照Ahmed 等[21]的方法提取水稻精米中的直链和支链淀粉,用80%的乙醇提取淀粉,加入乙醚溶液,离心后保留沉淀,再加入碱溶液充分溶解后备用待测。向待测液中加入盐酸和蒸馏水,混匀后加入碘化钾-碘溶液,混匀后测定其在550、620 和743 nm 处的吸光值,计算稻米直链和支链淀粉的含量和比例。
蛋白质含量的测定:采用二辛可宁酸(bicinchoninic acid,BCA)法测定蛋白质含量[22],取0.1 g精米样品加入蒸馏水,离心后取上清液待测。酶标仪预热后加入BCA 试剂、碱溶液和酒石酸钠,再加入硫酸铜溶液,混匀后于60 ℃保温30 min,冷却后测定其在562 nm 处的吸光值,根据样品质量计算稻米蛋白质含量。
蔗糖、葡萄糖和果糖含量的测定:采用间苯二酚法测定稻米蔗糖和果糖含量[23],采用氧化酶-过氧化物酶比色法测定稻米葡萄糖含量[24]。用于测定淀粉、蛋白质、蔗糖、果糖和葡萄糖含量的试剂盒由苏州梦溪生物医药科技有限公司提供。按照说明书要求,采用iMark 酶标仪(美国赛默飞世尔科技有限公司)测定各物质的含量。
1.5 数据统计与分析
采用Excel 2016 软件进行数据的统计和分析,采用Origin 2016 软件绘图。利用SPSS 21.0 软件进行方差分析,基于单因素方差分析(one-way ANOVA)和Duncan法进行显著性差异分析,显著性差异水平为P<0.05。利用一般线性模型(general linear model,GLM)方法比较品种和处理对产量及品质的影响。
2 结果与分析
2.1 不同生殖生长阶段高温胁迫对水稻生长发育的影响
由图1 可知,两个水稻品种在对照条件和不同时期高温胁迫下的株高和茎秆干重无显著差异,YLY1577 的株高高于HHZ,说明高温胁迫对不同生殖生长阶段水稻植株的生长发育状况无明显影响。不同生殖生长阶段高温胁迫下,水稻的穗长和穗重均低于对照。HHZ 的穗长在HS1 处理下较CK 处理显著下降12.10%(P<0.05),其他处理与CK无显著差异。YLY1577 的穗长在不同时期高温处理下均较CK 差异显著(P<0.05),HS1、HS2、HS3、HS4 和HS5 处理分别较CK 处理下降了12.65%、8.27%、6.38%、6.09%和5.76%(图1-C)。两个水稻品种在不同时期高温胁迫下的穗重均较对照整体显著下降(P<0.05),其中,HHZ 在HS3 处理下的下降幅度最大,较CK 下降了23.62%;YLY1577 在HS1 处理下的下降幅度最大,较CK 下降了19.12%(图1-D)。与CK 相比,两个水稻品种的一次枝梗和二次枝梗数均在HS1处理下显著下降(P<0.05),HHZ 和YLY1577 的一次枝梗数较CK 分别下降了26.67%和22.97%(图1-E);二次枝梗数较CK分别下降了36.56%和22.50%(图1-F)。

图1 不同生殖生长阶段高温胁迫对水稻植株生长发育的影响Fig.1 Effect of heat stress at different reproductive growth stage on plants growth in rice
2.2 不同生殖生长阶段高温胁迫对水稻产量结构的影响
不同生殖生长阶段高温胁迫和对照条件下,两个水稻品种的穗数差异不显著(图2-A),HHZ 的每穗粒数在HS1 处理下较对照显著下降(P<0.05)(图2-B)。生殖生长期不同时段高温胁迫导致HHZ 和YLY1577的实粒数较对照明显下降,且均在HS2 处理下降幅度最大,HHZ 和YLY1577 在HS2 处理下的实粒数较对照分别下降了23.57%和10.18%(图2-C)。不同生殖生长阶段高温胁迫导致两个水稻品种的瘪粒数整体显著增加(P<0.05),且均在HS2 处理下达到最大值,HHZ和YLY1577 在HS2 处理下的瘪粒数较对照分别增加了67.55%和41.92%(图2-D)。两个水稻品种在不同生殖生长阶段高温胁迫下的结实率较对照均显著下降(P<0.05),且均在HS2 处理下的下降幅度最大;HHZ 在HS1、HS2、HS3、HS4 和HS5 处理下的结实 率 较CK分别下降了7.32、12.87、5.73、5.20 和5.31 个百分点,YLY1577 则分别下降了3.96、8.12、4.58、3.17 和2.76 个百分点(图2-E)。两个水稻品种的千粒重在不同生殖生长阶段高温胁迫下均较对照显著下降(P<0.05),且均在HS3 处理下降幅度最大;HHZ 和YLY1577 在HS1、HS2、HS3、HS4、HS5 处理下的千粒重较CK 分别下降了6.15%、6.15%、14.66%、12.76%、11.68%和9.86%、9.81%、14.12%、11.12%、7.77%(图2-F)。
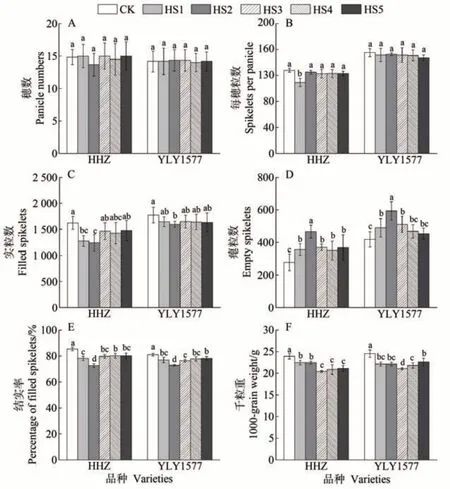
图2 不同生殖生长阶段高温胁迫对水稻产量结构的影响Fig.2 Effect of heat stress at different reproductive growth stage on yield component in rice
由图3 可知,不同生殖生长阶段高温胁迫导致两个水稻品种的产量和收获指数均较对照显著下降(P<0.05)。其中,HHZ 在HS1、HS2、HS3、HS4 和HS5处理下的收获指数较CK 分别下降了6.57、6.30、5.20、4.89 和4.43 个百分点,YLY1577 则分别下降了3.78、3.98、4.57、5.29 和4.21 个百分点(图3-B)。HHZ 在HS1、HS2、HS3、HS4 和HS5 处理下的单穴产量较CK 分别下降了26.28%、28.24%、22.97%、23.60%和19.78%,YLY1577 则分别下降了16.18%、18.68%、20.01%、17.84%和14.99%(图3-C)。

图3 不同生殖生长阶段高温胁迫对水稻产量的影响Fig.3 Effect of heat stress at different reproductive growth stage on grain yield in rice
2.3 不同生殖生长阶段高温胁迫对稻米加工和外观品质的影响
由图4 可知,高温胁迫影响了稻米的加工品质和外观品质,与CK 相比,高温胁迫下稻米出糙率、精米率和整精米率呈整体下降趋势,垩白粒率和垩白度升高。HS1和HS2处理下HHZ的出糙率较对照差异不显著(P>0.05),HS3、HS4 和HS5 处理下较对照显著下降(P<0.05)。不同生殖生长阶段高温胁迫下,YLY1577的稻米出糙率较对照均无显著差异(图4-A)。HS1、HS2 和HS5 处理下HHZ 和YLY1577 的精米率较对照下降较小,HS3和HS4处理精米率下降明显,HHZ的精米率在HS3 和HS4 处理下较对照分别下降了9.98 和8.95个百分点,YLY1577分别下降了9.05和10.27个百分点(图4-B)。HS3和HS4处理的稻米整精米率较CK下降幅度较大(图4-C)。两个品种稻米的长宽比在HS3和HS4处理下变小,但较对照均无显著差异(图4-D)。两个水稻品种的垩白粒率和垩白度在HS1 处理下较CK 均无显著差异,HS2、HS3 和HS4 处理较CK 均显著增加(P<0.05)(图4-E、F)。HHZ 在HS2、HS3 和HS4处理下的垩白粒率较CK 分别增加了6.20、7.67 和6.37 个百分点,垩白度分别增加了1.53、3.28 和2.98个百分点。YLY1577 在HS2、HS3 和HS4 处理下的垩白粒率较CK 分别增加了3.92、6.07 和6.32 个百分点,垩白度分别增加了1.62、2.37和2.22个百分点。
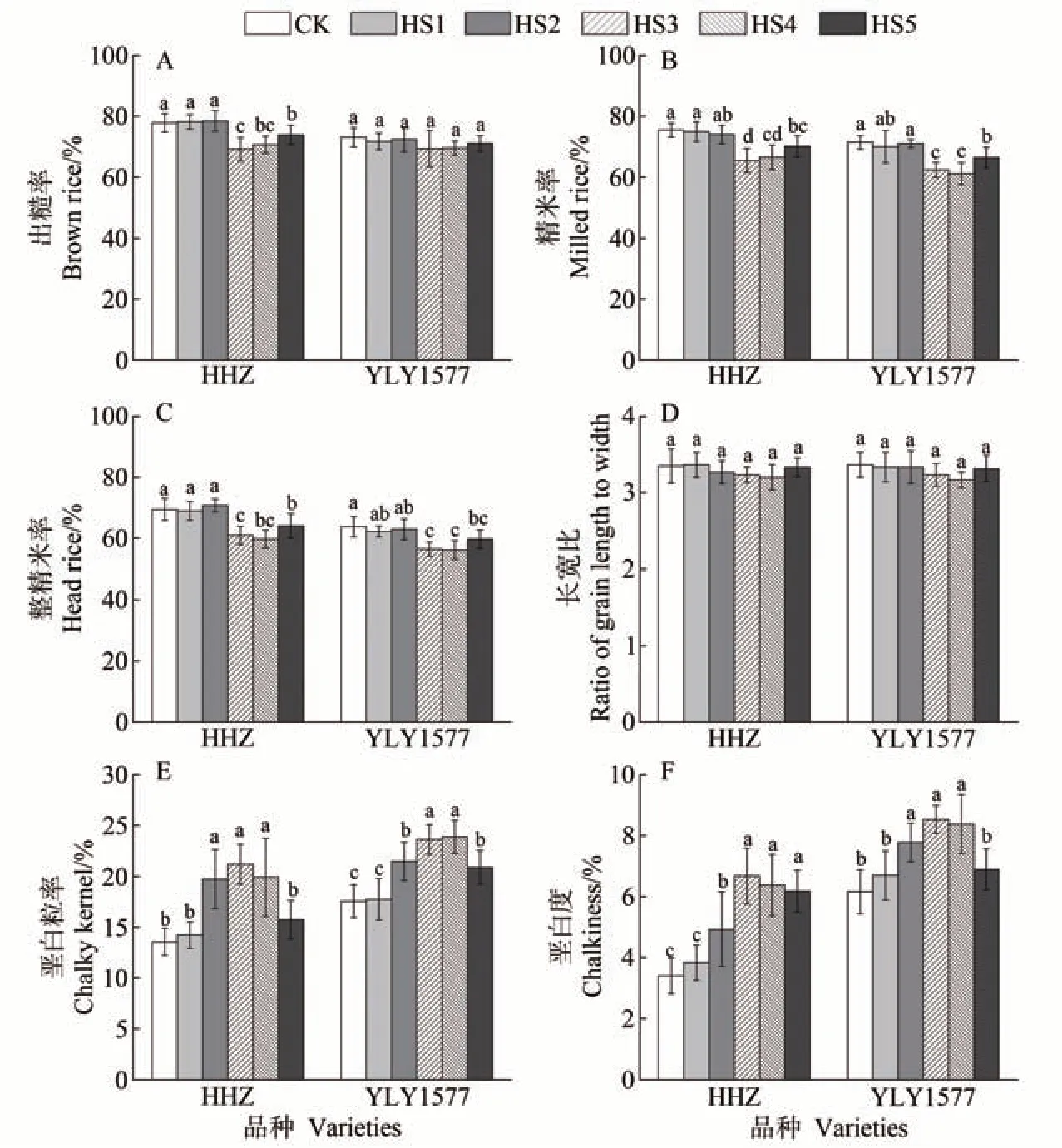
图4 不同生殖生长阶段高温胁迫对稻米加工及外观品质的影响Fig.4 Effect of heat stress at different reproductive growth period on milling and appearance quality of rice
2.4 不同生殖生长阶段高温胁迫对稻米淀粉含量和胶稠度的影响
对稻米的淀粉含量进一步分析表明,HHZ 的稻米直链淀粉含量在不同生殖生长阶段高温胁迫下较CK整体显著下降(P<0.05),以HS3 处理下降幅度最大;YLY1577的稻米直链淀粉含量在HS1和HS2处理下与CK 无显著差异,HS3~HS5 处理下显著下降(P<0.05)。HHZ 和YLY1577 的直链淀粉含量在HS3 处理下较CK分别下降了39.86%和37.18%(图5-A)。两个水稻品种的直链淀粉比例在HS3~HS5 处理下均较CK 明显下降,且在HS3 和HS4 处理下较CK 显著下降(P<0.05),其他处理较CK 无显著差异(图5-C)。HHZ 的支链淀粉含量在HS2~HS5处理下均较CK 显著下降(P<0.05)(图5-B),支链淀粉比例在各处理间差异不显著(图5-D)。YLY1577 的支链淀粉含量在HS3、HS4 处理下较CK显著下降(P<0.05)(图5-B),支链淀粉比例在各处理间也较CK 明显下降(图5-D)。HHZ 和YLY1577 的总淀粉含量在HS2~HS5 处理下均较CK 整体显著下降(P<0.05),且均以HS3 处理降幅最大,HHX 和YLY1577 的总淀粉含量在HS3 处理下分别较CK 下降了18.70%和14.09%(图5-E)。两个水稻品种的稻米胶稠度在HS2~HS5 处理下较对照整体显著下降,以HS3处理降幅最大(图5-F)。

图5 不同生殖生长阶段高温胁迫对稻米淀粉含量和胶稠度的影响Fig.5 Effect of heat stress at different reproductive growth stage on starch content and gel consistence in rice
2.5 不同生殖生长阶段高温胁迫对稻米RVA特征谱的影响
由图6 可知,生殖生长期高温胁迫影响了稻米淀粉黏滞性特征,孕穗期高温胁迫对淀粉最终黏度、崩解值、消减值和糊化温度等指标无明显影响。始穗后高温胁迫导致稻米淀粉最终黏度、消减值和糊化温度升高,崩解值下降,不同时段高温胁迫对不同水稻品种稻米淀粉RVA 特征谱的影响具有差异性。HHZ 的淀粉最终黏度在HS4 处理下最高,较对照显著增加了15.15%;YLY1577的淀粉最终黏度在HS3处理下最高,较对照显著增加了17.76%(图6-A)。两个水稻品种的淀粉崩解值在不同时段高温胁迫下的变化趋势基本一致,HHZ 在HS3 和HS4 处理较对照下降幅度较大,分别显著下降了13.31%和13.54%,但两者间差异不显著;YLY1577 在HS3 处理下降幅度最大,较CK 显著下降了17.38%(图6-B)。HHZ 的淀粉消减值在HS3 和HS4 处理下较高,分别较对照显著增加了20.17%和20.30%;YLY1577 在HS3 处理下的淀粉消减值最高,较对照显著增加了20.75%(图6-C)。两个水稻品种的淀粉糊化温度在高温胁迫下的变化趋势基本一致,始穗后高温胁迫导致稻米糊化温度增加,且均以HS3和HS4处理增加幅度较大(图6-D)。

图6 不同生殖生长阶段高温胁迫对稻米淀粉RVA特征谱的影响Fig.6 Effect of heat stress at different reproductive growth period on starch RVA characteristics in rice
2.6 不同生殖生长阶段高温胁迫对稻米蛋白质和糖类物质含量的影响
由图7 可知,孕穗期高温胁迫下两个水稻品种的稻米蛋白质含量较对照差异不显著,始穗后高温胁迫导致两个水稻品种的稻米蛋白质含量整体明显增加。与CK 相比,HHZ 的蛋白质含量在HS3 处理下的增幅最大,较对照显著增加了19.74%;YLY1577 的蛋白质含量在HS4 处理较对照增幅最大,显著增加了13.34%,HS3 和HS4 处理间差异不显著(图7-A)。与CK相比,HHZ的蔗糖含量在HS1和HS2处理下显著下降;YLY1577 的蔗糖含量在HS1 处理下显著下降,HS4处理下显著增加(图7-B)。两个水稻品种的稻米葡萄糖含量在不同时期高温处理下较对照均显著增加,且在HS3 处理下增加幅度最大,较CK 分别增加了15.92%和18.83%(图7-C)。不同时期高温处理下两个水稻品种的稻米果糖含量较对照均显著增加,且在HS3~HS5处理下增加较明显(图7-D)。
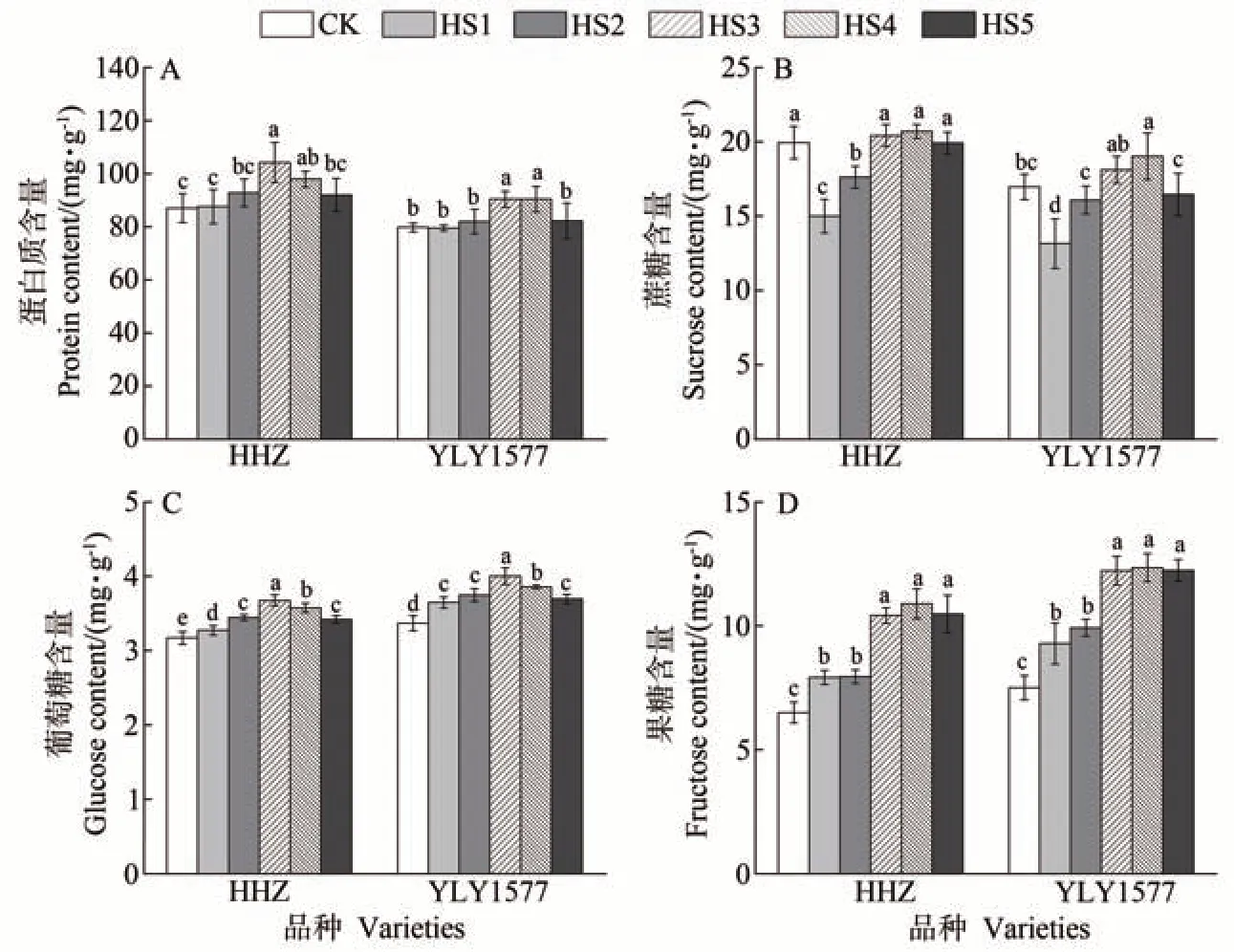
图7 不同生殖生长阶段高温胁迫对稻米蛋白质和糖类物质含量的影响Fig.7 Effect of heat stress at different reproductive growth period on protein and carbohydrate content in rice
2.7 水稻产量和品质指标的方差分析
由表1 可知,品种和处理对水稻的收获指数、产量、淀粉含量、稻米外观和食味品质指标影响极显著(P<0.01),品种和处理互作对水稻产量形成和稻米品质指标的影响不显著(P>0.05),说明高温胁迫对水稻产量形成和品质的影响具有协同性,水稻产量和品质对高温胁迫的响应因品种而异。
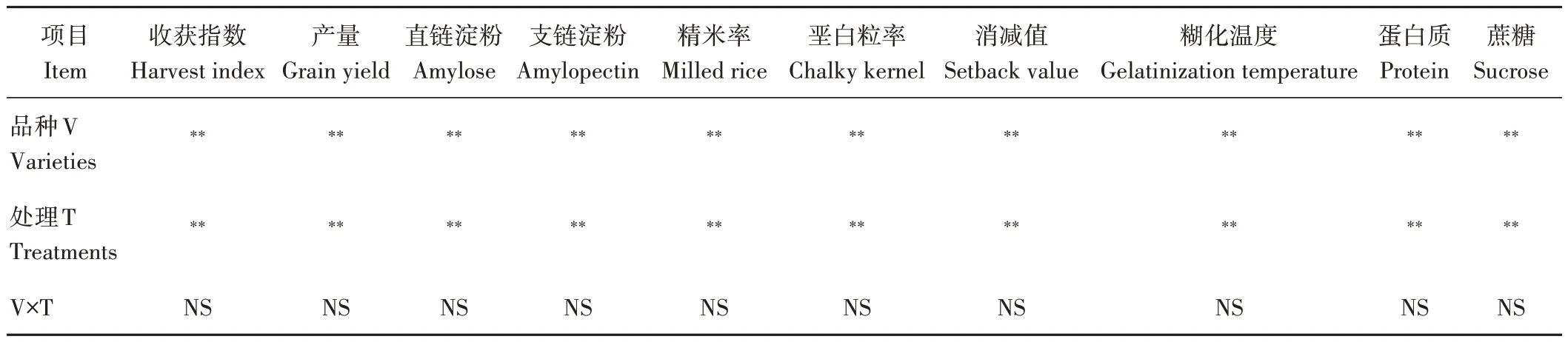
表1 水稻产量和品质指标的方差分析Table 1 Variance analysis of yield and quality indexes in rice
3 讨论
高温胁迫对不同生长发育阶段的水稻生长发育均有影响,特别是生殖生长期的高温胁迫可直接影响水稻产量和稻米品质[25]。本研究结果表明,孕穗期至灌浆后期不同时段高温胁迫均导致水稻产量显著下降,黄华占和Y两优1577分别在始穗后1~7和8~14 d遭遇高温胁迫时的产量损失最大,表明抽穗开花期和灌浆初期是高温胁迫导致水稻减产的重要时段,这与前人研究结果一致[5-16,26-27]。孕穗期高温胁迫对稻米外观品质、加工品质和淀粉含量无明显影响,始穗后不同时段高温胁迫对稻米品质均产生不同程度影响,且在始穗后8~14 d 高温胁迫下变化最为明显,说明该时段是高温胁迫影响稻米品质的关键时期[28-29],这与前人研究结果基本一致[30]。
高温胁迫显著影响水稻的产量结构指标[7-9,14]。本试验中,不同生殖生长阶段高温胁迫对水稻生长指标和穗数影响不大,这主要与水稻在营养生长期已达到稳定的穗数有关。然而,孕穗期高温胁迫导致一次和二次枝梗数较对照显著下降,穗长较其他处理也明显下降,说明高温胁迫对水稻枝梗分化影响明显,可能与高温胁迫影响枝梗分化进程有关[31-32]。不同生殖生长阶段高温胁迫下,结实率和千粒重分别在始穗后1~7 和8~14 d 高温处理下的下降幅度最为明显,说明抽穗开花期高温胁迫主要影响籽粒结实过程,而灌浆初期高温胁迫主要影响淀粉合成过程,收获指数和谷物产量与穗重呈现相似的变化趋势,进一步说明高温胁迫对水稻产量的影响主要在于降低结实率和千粒重,这与前人研究结果基本一致[7,9,11]。因此,当水稻在孕穗期遭遇高温天气时,幼穗发育首先受到抑制,导致枝梗数下降;若高温天气持续进行,将降低结实率;长期的高温天气还会继续影响籽粒灌浆进程,使千粒重下降,最终导致产量下降。始穗后1~7 d 和8~14 d 高温胁迫下水稻产量损失最大,这与前人研究结果基本一致[15-16]。
直链淀粉含量、胶稠度和淀粉RVA 特征谱等指标是衡量稻米食味蒸煮品质的重要指标[33],这些指标易受高温影响,进而影响稻米品质[19]。本试验中,高温胁迫导致总淀粉、直链淀粉和支链淀粉含量整体显著下降,始穗后8~14 和15~21 d 高温处理对直链和支链淀粉含量影响更明显,说明该时段高温是影响稻米淀粉合成的关键时期[34]。淀粉胶稠度在始穗后8~14 d下降明显,这可能是由于高温胁迫降低籽粒实际直链淀粉含量,增加稻米流胶长度,进而降低胶稠度[35]。此外,在本试验中,不同生殖生长阶段高温胁迫影响了淀粉的RVA特征谱,孕穗期高温胁迫对稻米RVA特征谱影响较小,始穗后8~21 d是稻米淀粉RVA 特征谱对高温胁迫响应的敏感时期,这与前人研究结果基本一致[19],此时的高温胁迫导致淀粉最终黏度、消减值和糊化温度升高,崩解值降低,进而影响稻米食味品质。抽穗灌浆期不同时段高温胁迫导致稻米蛋白质含量升高,这与多数前人研究结果相一致[36-38],可能是由于此时的高温胁迫影响籽粒中的三羧酸循环过程,增加了蛋白质含量[13]。
高温胁迫会影响蔗糖-淀粉代谢途径中的酶活性,影响稻米中蔗糖、葡萄糖和果糖等物质的含量[39]。本研究结果表明,孕穗期和始穗1~7 d 高温胁迫导致稻米蔗糖含量下降,可能与高温胁迫影响该时期的光合效率、抑制光合产物积累有关[40]。始穗后第8 天水稻处于灌浆初期,此时的高温胁迫影响蔗糖-淀粉代谢途径灌浆酶活性,导致籽粒蔗糖、葡萄糖和果糖含量增加,影响淀粉生成,这与前人研究结果基本一致[41]。始穗后8~21 d 高温胁迫下,稻米蔗糖含量较高,葡萄糖和果糖含量增加,此时淀粉糊化温度和消减值的增加可能与稻米内较高的糖含量有关,表明稻米糖含量的变化可能是高温胁迫影响淀粉粘滞性的一个重要因素[42],以上研究结果与前人研究结论相吻合[30,39,42]。本试验中,杂交籼稻品种Y 两优1577 的产量高于黄华占,但稻米品质和淀粉含量略低于黄华占,在耐热型水稻品种选育中应着重考虑其对抽穗开花期和灌浆初期高温胁迫的抗性,并综合考虑稻米品质等指标。
4 结论
孕穗期高温胁迫主要通过抑制穗部发育影响产量形成,对稻米品质影响较小。始穗后1~7 和8~14 d 高温胁迫是导致水稻减产的关键时期,主要通过降低结实率和千粒重影响水稻产量。始穗后8~21 d 是稻米品质对高温胁迫响应的敏感时期,该时段高温胁迫会影响稻米外观和加工品质,降低食味品质。水稻在始穗后8~14 d 遭遇高温胁迫会导致产量和品质的双重损失,在生产中可通过调整播期、水肥管理和喷施植物生长调节剂等措施缓解高温胁迫,水稻品种在始穗后8~14 d的产量和稻米品质变化及其对高温胁迫的响应可作为选育耐高温优质水稻品种的参考方向。
