三色堇酪氨酸脱羧酶基因表达的位置效应
2023-09-07刘国锋张文娥
饶 英 林 颖 王 健 刘国锋 张文娥 彭 婷,*
(1贵州大学农学院/山地植物资源保护与种质创新教育部重点实验室,贵州 贵阳 550025;2海南省热带特色花木资源生物学重点实验室/海南大学林学院,海南 海口 570228;3广州市林业和园林科学研究院,广东 广州 510405)
三色堇(Viola×wittrockiana)为堇菜科(Violaceae)堇菜属(Viola)植物,是世界范围内重要的冬春季花坛花卉,具有典型的花瓣色斑性状,是观赏植物花色、花斑研究的理想材料。目前国内外对三色堇花色、花斑的研究主要集中在花色素成分鉴定[1-2]及花青素合成相关基因的分离与表达分析等方面[3-4]。
酪氨酸脱羧酶(tyrosine decarboxylase, TyDC)是连接植物初生代谢与次生代谢的初始酶[5],可将底物酪氨酸脱羧形成酪胺[6-8]。在天然药源植物红景天(Rhodiola rosea)和地黄(Rehmannia glutinosa)中,TyDC对红景天苷和麦角甾苷的合成分别起促进作用[9-11]。近期的一些研究认为TyDC 可能与花色形成有关。如Zhou 等[12]研究发现拟南芥TyDC 的作用底物酪氨酸可以诱导花青素的合成,对酪氨酸降解缺陷突变体fah进一步研究发现,在突变体中,酪氨酸对花青素的诱导效果显著低于野生型,且突变体中花青素合成的结构基因和调控基因表达量的提升幅度显著低于野生型,说明酪氨酸的降解产物诱导了花青素的积累。本课题组前期在三色堇品种黄色带花斑中的研究发现,VwTyDC基因在花瓣中的表达量显著高于根、茎和叶,并且在色斑区的表达量高于非色斑区[13]。Cui等[2]用酪氨酸对三色堇非色斑区进行处理,发现非色斑区出现了条纹状的色斑,通过病毒介导的基因沉默(virusinduced gene silencing, VIGS)技术沉默VwTyDC基因后,三色堇色斑区的颜色明显变浅,说明VwTyDC基因可能与花青素的合成有关。
本课题组在转录组中发现了一个新基因并命名VwTyDC-like,该基因在三色堇花瓣不同区域存在差异表达。为进一步探究VwTyDC-like基因在三色堇中的精细表达模式,本研究克隆获得VwTyDC-like基因的cDNA 及gDNA 序列,并与VwTyDC进行对比分析。同时,通过实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)技术分析VwTyDC和VwTyDC-like基因在三色堇不同组织器官、花瓣发育不同时期、不同花色品种及其唇瓣不同区域的表达模式,以期为探究VwTyDC与VwTyDC-like基因在三色堇中的生物学功能奠定基础,为三色堇新品种培育及植物花色形成机理研究提供新的思路和途径。
1 材料与方法
1.1 试验材料
以三色堇超级宾哥(Matrix)系列的不同花色品种,包括黄色(Yellow)、黄色带花斑(Yellow Blotch)、纯蓝色(True Blue)、蓝色带花斑(Blue Blotch)、白色(White)和白色带花斑(White Blotch),帝国巨人Ⅱ(Majestica GiantsⅡ)系列的淡紫色渐变(Lavender Shades)和得大(Delta)系列的紫罗兰白双色(Violet & White)(图1)为供试材料,所有种子均购于美国泛美种子公司。于2021年8月上旬将不同品种的三色堇种子播种在广州市观赏植物种质资源圃,采用常规栽培管理措施,待2021 年10 月上旬三色堇进入始花期,分区域和发育时期采集不同品种的花瓣进行后续试验。

图1 不同花色的三色堇品种Fig.1 Pansy cultivars with different flower colors
1.2 试验方法
1.2.1 样品采集 样品采集分为四组,第一组:采集黄色带花斑品种的根、茎、叶、花芽、花、幼果和种子用于不同组织器官的表达分析;第二组:参照Li等[1]的方法将三色堇的花发育过程划分为7个时期(S1~S7),采集黄色带花斑品种7 个时期花瓣中心色斑区(a)与非色斑区(b)的样品,用于不同花发育时期的表达分析;第三组:将白色带花斑和白色2个品种的唇瓣由内向外划分为5个区域(区域1~5)进行采样(图1),用于唇瓣不同区域的表达分析;第四组:收集5 个不同花色品种(黄色、纯蓝色、蓝色带花斑、粉紫色渐变和紫罗兰白双色)花发育S7 期中心色斑区(无斑品种为中心色斑对应区,a)、非色斑区(无斑品种为非色斑对应区,b)及边缘色斑区(仅紫罗兰白双色品种有此区域,c)的样品,用于不同花色三色堇品种在S7期的表达分析(图1)。每个样品设置3 个生物学重复,于液氮中速冻后,-80 ℃条件下保存备用。
1.2.2 植物核酸提取和cDNA合成 以三色堇的嫩叶或花瓣为材料,采用改良溴化十六烷基三甲胺(cetyl trimethylammonium bromide, CTAB)法提取总DNA,利用EASYspin植物RNA快速提取试剂盒(艾德莱,北京)提取不同品种、时期及组织品种样本总RNA,将检测合格的RNA 样品按照Prime ScriptTMRTreagent kit with gDNA Eraser(Perfect Real Time)试剂盒(TaKaRa,大连)说明书进行反转录,反转录后的cDNA 样品置于-20 ℃冰箱保存备用。
1.2.3VwTyDC-like基因克隆和结构分析 基于黄色带花斑品种转录组数据库筛选到1 个VwTyDC-like(Unigene0055670),结合分离VwTyDC-like基因启动子时得到的序列,利用Prime 5.0 软件设计特异引物(表1),分别扩增cDNA 及gDNA 序列。将PCR 产物用胶回收试剂盒(艾德莱,北京)进行回收后,连接到pMD19-T 载体,转化大肠杆菌DH5α 感受态细胞,挑选单菌落经PCR 检测后,送至上海生工生物公司测序。利用在线网站GSDS(http://gsds.gao-lab.org/)分析该基因的外显子-内含子结构,并利用Exon-Intron Graphic Maker 在线网站(http://www.wormweb.org/exonintron)绘制基因构图。
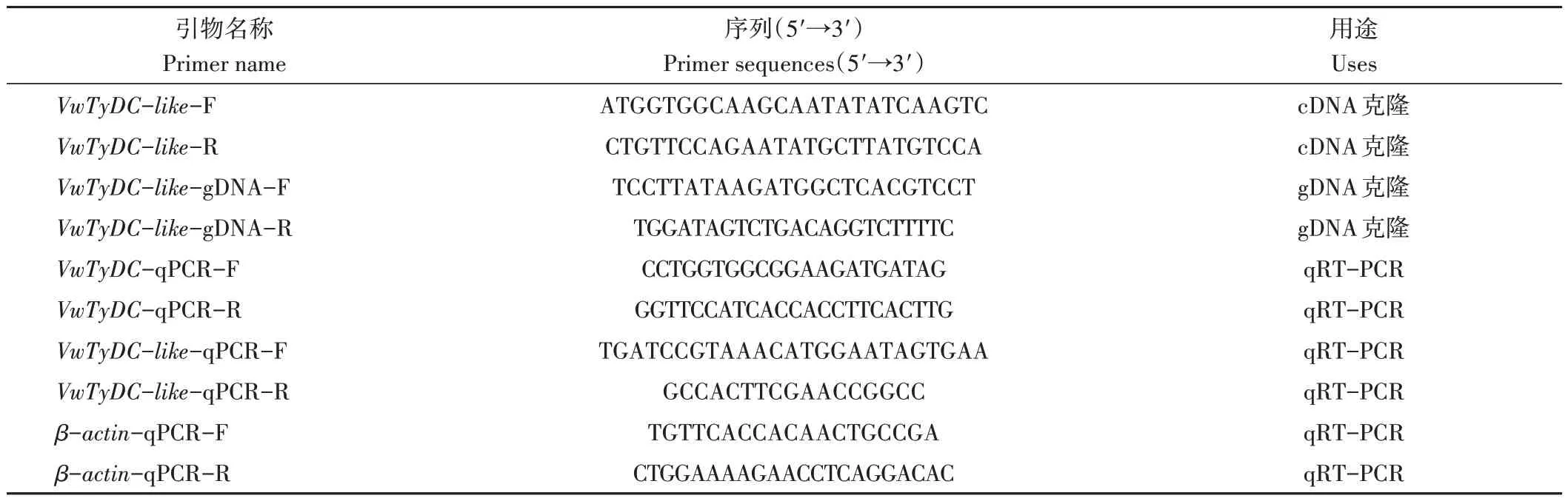
表1 本研究所用引物Table 1 Primer sequences used in this study
1.2.4 生物信息学分析 利用ProtParam 在线分析软件(https://web. expasy. org/protparam/)分析VwTyDClike蛋白的理化性质,VwTyDC-like 蛋白的二级结构和三级结构预测分别使用在线工具SOPMA(https://npsa-prabi. ibcp. fr/cgi-bin/secpred_sopma. pl)和SWISSMODEL(https://swissmodel. xpasy. org/)完成。将获得的VwTyDC-like 序列在NCBI 数据库中(https://blast.ncbi. nlm. nih. gov/Blast. cgi)进行比对,下载其他物种的同源序列并利用DNAMAN6.0 软件进行氨基酸序列多重比对。利用MEGA 11 软件采用最大似然法(maximum likelihood,ML)构建系统发育树,bootstrap值设为1 000。
1.2.5 基因表达分析 以反转录cDNA 为模板,按照TB Green®Premix Ex Taq™ II(Tli RNaseH Plus)试剂盒(TaKaRa,大连)操作说明,利用实时荧光定量PCR(quantitative real-tive PCR, qRT-PCR)技术分析VwTyDC-like和VwTyDC在三色堇不同组织器官、花发育的不同时期、不同品种中的表达模式。利用Primer 5.0 设计引物(表1),以三色堇β-actin作为内参基因[15],以2-ΔΔCT法计算目的基因的相对表达量。每个样本均设置3 个生物学重复,3 次技术重复。使用Excel2016 和SPSS19.0 软件对数据进行分析,利用Duncan 新复极差法做显著性分析(P<0.05),并采用Origin 2019b软件作图。
2 结果与分析
2.1 VwTyDC-like基因的克隆与序列分析
从前期获得的黄色带花斑三色堇转录组数据库中筛选到1 个在花瓣不同区域存在差异表达的Unigene。以黄色带花斑品种花瓣的cDNA 为模板,通过设计特异引物进行PCR 扩增,得到1 条特异条带(图2-A)。测序后获得的编码序列(coding sequence, CDS)序列长1 497 bp,编码498个氨基酸。该基因序列与VwTyDC高度相似,命名为VwTyDC-like基因,GenBank 登录号为ON866521。同时,以黄色、纯蓝色、白色3个无斑品种花瓣的cDNA 为模板通过PCR 扩增VwTyDC-like和VwTyDC的核苷酸序列。结果显示,3 个品种VwTyDClike基因的核苷酸序列与黄色带花斑品种完全一致;纯蓝色、白色品种VwTyDC基因的核苷酸序列与黄色带花斑也完全一致,黄色品种的核苷酸序列与黄色带花斑品种相比,仅有1 个碱基的差异,但编码氨基酸序列一致。以黄色带花斑叶片总DNA为模板,扩增得到单一条带(图2-B),长度为2 340 bp(包含部分启动子序列),GenBank 登录号为ON866520。基因结构分析发现,VwTyDC-like基因包含2个外显子和1个内含子(图2-C)。
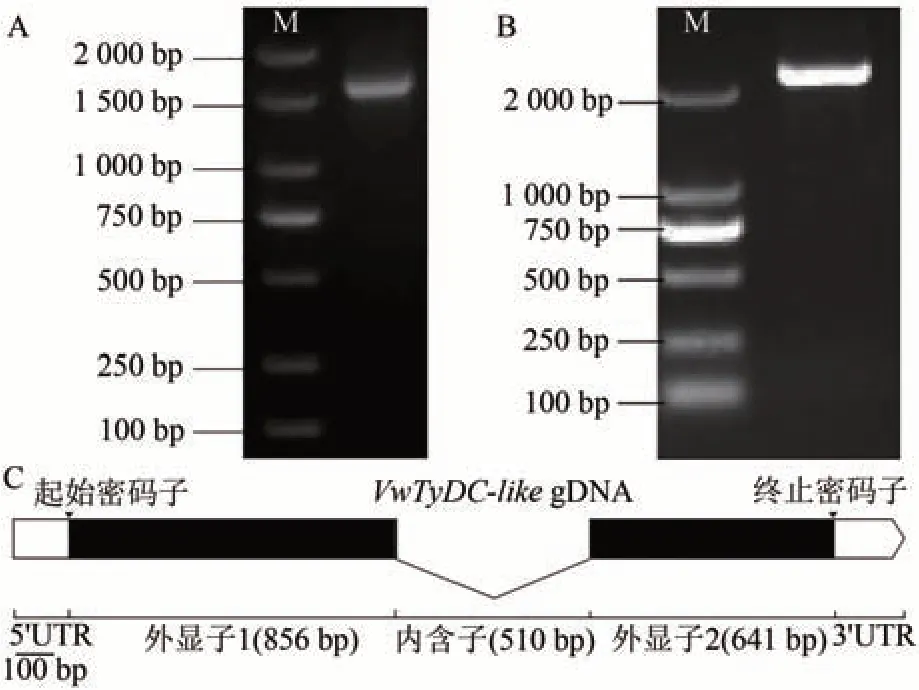
图2 VwTyDC-like基因克隆与结构分析Fig.2 Cloning of VwTyDC-like and exon-intron structure analysis
2.2 VwTyDC-like的序列分析
利用ExPASy中的ProtParam tool分析VwTyDC-like蛋白的理化性质,结果表明该蛋白的相对分子量为54.94 kDa,理论等电点为5.84,脂肪系数86.97,总平均亲水性为-0.052。利用ProtScale 程序分析VwTyDC-like 蛋白亲水/疏水性,结果显示其属于亲水性蛋白,二级结构与三级结构预测结果显示2 个蛋白主要由α 螺旋和无规则卷曲组成。用DNAMAN 对来自不同物种的TyDC 蛋白进行序列比对,发现VwTyDC-like与VwTyDC(ASW27173.1)氨基酸序列有37 个位点存在差异,与狗尾草(Setaria viridis)、橡胶树(Hevea brasiliensis)和毛果杨(Populus trichocarpa)等物种的同源性为66.18%(图3)。使用MEGA 11软件对三色堇及其他物种的TyDC蛋白构建系统进化树,结果显示,VwTyDC-like 蛋白与三色堇VwTyDC 蛋白进化关系最近,与橡胶树TyDC 蛋白亲缘关系相对较近,与葡萄(Vitis riparia)TyDC 蛋白亲缘关系相对较远,与单子叶植物狗尾草和玉米(Zea mays)TyDC 蛋白亲缘关系最远(图4)。

图3 VwTyDC-like与其他物种TyDCs蛋白序列比对Fig.3 Protein sequence alignment of VwTyDC-like and other TyDCs from different plants
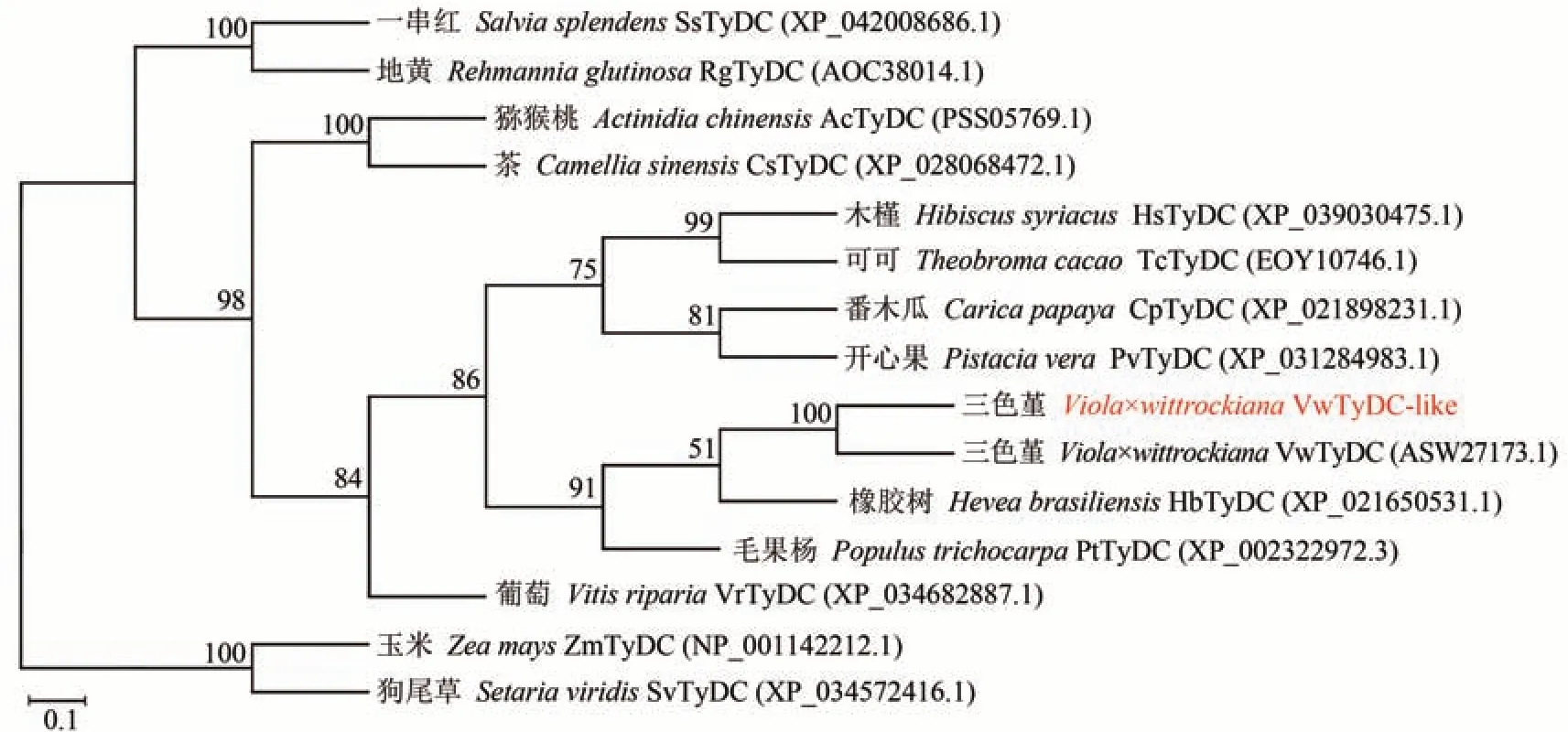
图4 三色堇与其他物种TyDC的进化树分析Fig.4 Phylogenetic tree of VwTyDC-like and VwTyDC with other TyDCs from different plants
2.3 VwTyDC-like 与VwTyDC 在不同组织及不同花发育时期的表达模式
qRT-PCR 结果显示,VwTyDC-like基因在超级宾哥系列黄色带花斑品种中的表达具有组织特异性,在花中的表达量显著高于其他组织(图5-A)。在花发育的S7 期,VwTyDC-like在花瓣基部的中心色斑区(a)的表达量显著高于非色斑区(b),并达到峰值,此前S1~S6期该基因在中心色斑区(a)与非色斑区(b)的表达量无显著差异(图5-C)。进一步分析显示,VwTyDC与VwTyDC-like基因呈现出相似的表达模式(图5)。
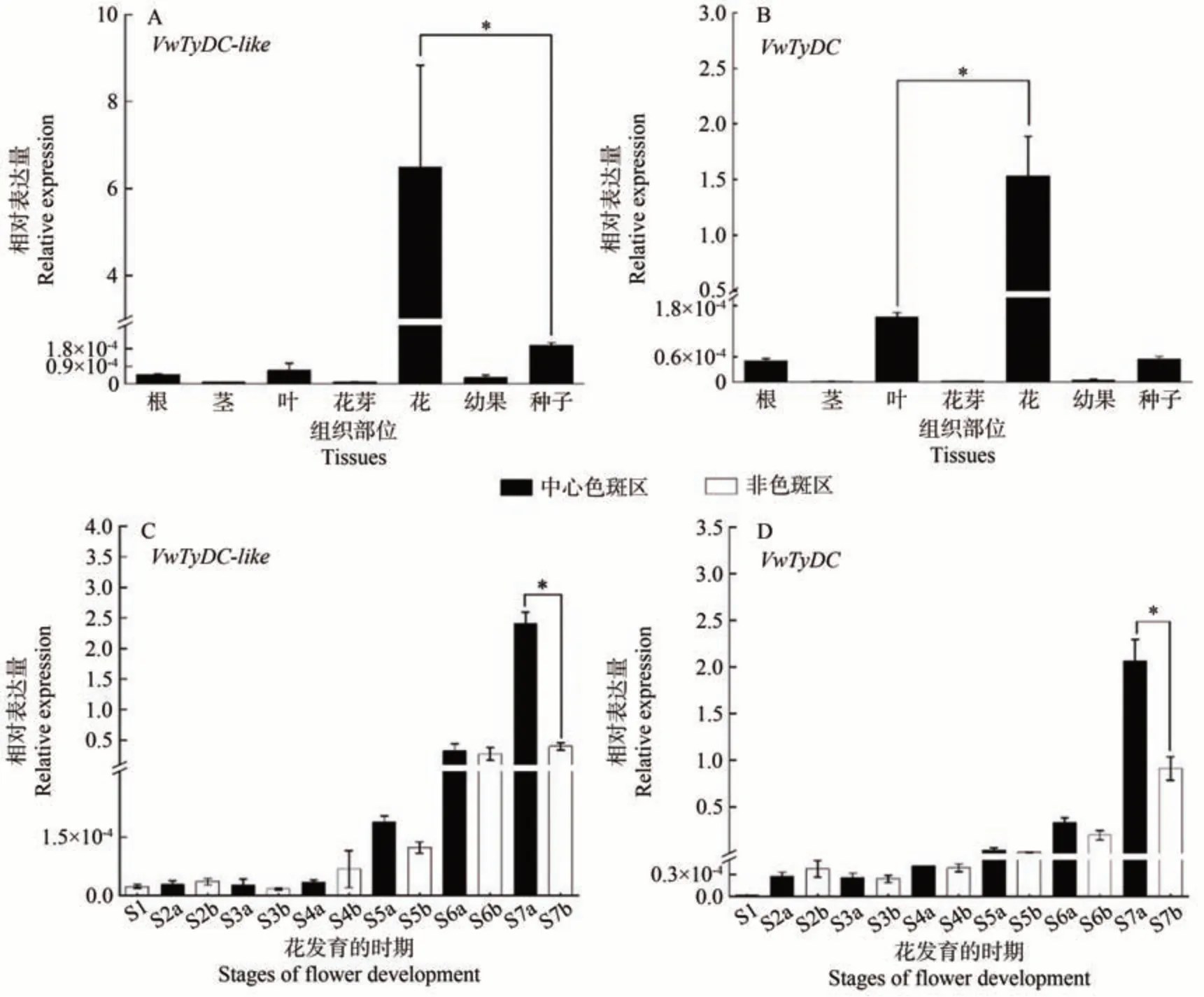
图5 VwTyDC-like与VwTyDC在黄色带花斑品种不同组织(A、B)和花发育的不同时期的表达分析(C、D)Fig.5 Expression analysis of VwTyDC-like and VwTyDC in different tissues (A, B) and at different flower developmental stages in Yellow Blotch cultivar (C, D)
2.4 VwTyDC-like与VwTyDC在花瓣中的表达模式
本研究利用得大系列的紫罗兰白双色品种分析了2 个基因的表达模式,该品种的色斑分布比较特殊,除花瓣基部有较大块的色斑以外,下方三枚花瓣的边缘也有小面积的色斑(图1-H)。qRT-PCR 分析结果表明,在花发育的S7期,2个基因在中心色斑区(a)的表达量显著高于边缘色斑区(c)和非色斑区(b);此外,非色斑区(b)的表达量高于边缘色斑区(c)(图6-A、B),推测VwTyDC-like与VwTyDC基因在花瓣中的表达可能具有位置效应。
为进一步明确VwTyDC-like与VwTyDC在花瓣中的表达分布是否与位置有关,本研究将白色带花斑和白色品种在S7 期的唇瓣划分成5 个区域,即从花瓣基部到花瓣边缘分为区域1~5(图7-A),采用qRT-PCR分析VwTyDC-like与VwTyDC在唇瓣不同区域的表达模式。结果显示,2 个基因明显呈现出从基部到边缘表达量逐渐下降的趋势,在唇瓣基部(区域1,无色斑)的表达量均显著高于其他位置(区域2~5),且在唇瓣边缘(区域5)几乎检测不到2 个基因的表达(图7),说明VwTyDC-like与VwTyDC在花瓣中的表达分布的确与位置有关。同时,VwTyDC-like基因的表达量远高于VwTyDC,达到了后者的7~13倍(图6、7)。
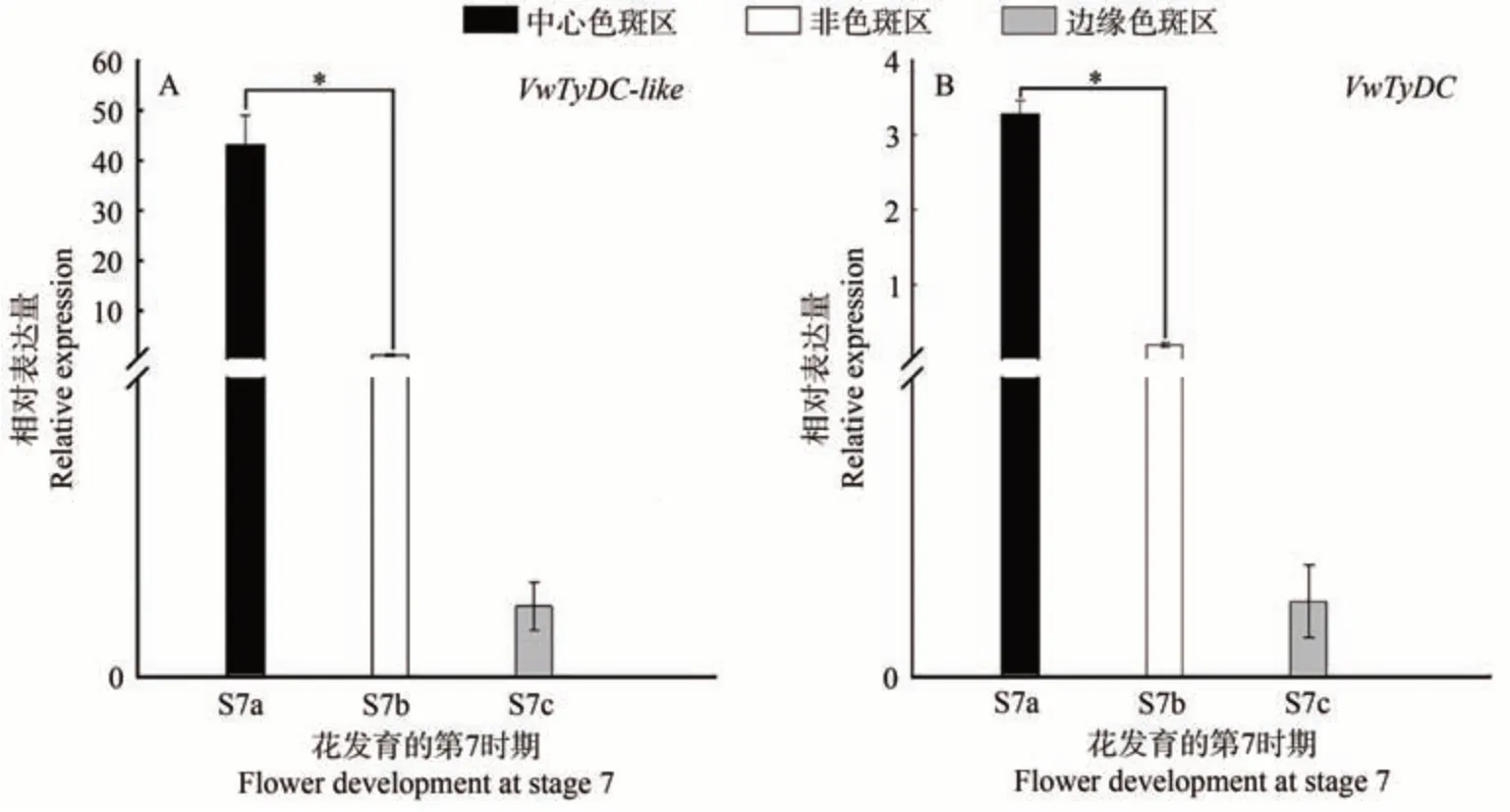
图6 VwTyDC-like与VwTyDC基因在紫罗兰白双色品种中心色斑区(a)、非色斑区(b)和边缘色斑区(c)的表达分析Fig.6 Expression analysis of VwTyDC-like (A) and VwTyDC (B) in the central blotch areas (a)、 non-blotch areas (b) and marginal blotch areas (c) of pansy petals in Violet & White cultivar

图7 白色带花斑和白色品种唇瓣不同区域(A)中VwTyDC-like (B)与VwTyDC (C)的表达分析Fig.7 Expression analysis of VwTyDC-like (B) and VwTyDC (C) in different areas of labella (A) in White Blotch and White cultivars
2.5 VwTyDC-like 与VwTyDC 在不同花色品种中的位置效应
为了解VwTyDC-like与VwTyDC基因表达的位置效应在其他三色堇品种中是否广泛存在,本研究还检测了二者在不同系列不同花色三色堇品种中的表达情况,包括超级宾哥系列黄色、纯蓝色、蓝色带花斑和帝国巨人II 系列的淡紫色渐变品种。结果发现,在有斑品种淡紫色渐变和蓝色带花斑中,VwTyDC-like与VwTyDC基因在花发育的S7 期,中心色斑区(a)的表达量显著高于非色斑区(b)。而且,在无斑品种黄色和纯蓝色中,VwTyDC-like与VwTyDC基因的表达量在中心色斑对应区(a)同样显著高于非色斑对应区(b),无斑品种表现出与有斑品种相似的表达特点(图8)。说明VwTyDC-like与VwTyDC基因表达的位置效应在其他三色堇品种中同样存在。同时,VwTyDC-like在不同品种中的表达量明显高于VwTyDC,在淡紫色渐变品种中,VwTyDC-like的表达量比VwTyDC高252倍(图8)。
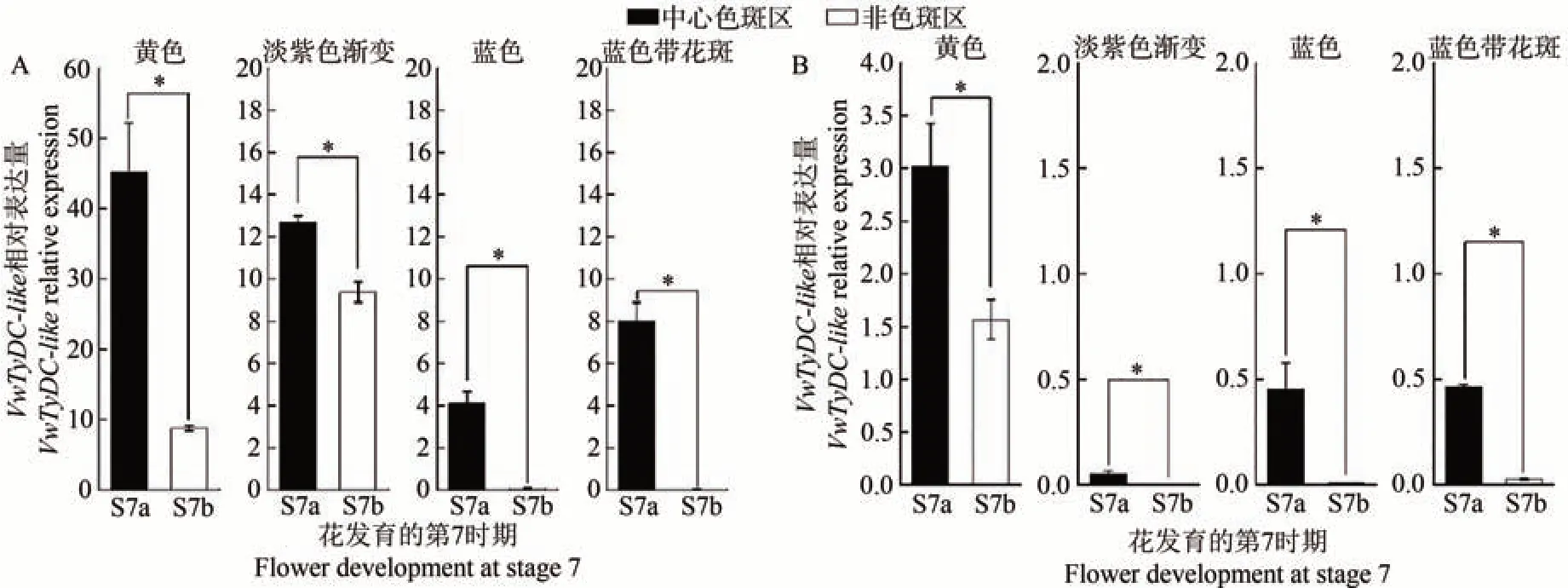
图8 VwTyDC-like(A)与VwTyDC(B)基因在不同花色品种不同区域的表达分析Fig.8 Expression analysis of VwTyDC-like (A) and VwTyDC (B) in the different areas of pansy petals with or without blotches in the pansy cultivars with different flower colors
3 讨论
目前有关植物TyDC基因的研究较少,主要集中在逆境胁迫应答[14-22]和酪氨酸衍生物合成路径中活性分子的生物合成等方面[5-11,23-24]。本研究从三色堇中分离了一个新的酪氨酸脱羧酶基因(VwTyDC-like),并将该基因与之前报道的VwTyDC进行了精细表达模式的比较分析。
3.1 VwTyDC-like与VwTyDC的同源性
VwTyDC-like与VwTyDC基因具有相似的核苷酸序列,氨基酸序列一致性为92.37%。蛋白质二级结构与三级结构预测结果显示2 个蛋白主要由α 螺旋和无规则卷曲组成。同时,qRT-PCR 分析结果显示,2个基因在三色堇不同组织器官、花发育不同时期、唇瓣不同区域、不同花色品种中呈现出高度一致的表达模式,在花中表达量最高;在花发育不同时期,表达差异仅在花发育的后期(S7)出现;无论品种有无色斑存在,在不同花色品种及其唇瓣的不同区域,表达量均由花瓣基部向外递减。VwTyDC-like与VwTyDC基因有着高度相似的氨基酸序列及表达模式,说明二者为同源基因,可能具有相似的生物学功能。值得注意的是,VwTyDC-like的表达量明显高于VwTyDC(图5~8),暗示VwTyDClike可能在同源基因发挥功能的过程中具有更大作用。
3.2 VwTyDC-like与VwTyDC基因表达的位置效应
本研究对超级宾哥系列黄色带花斑品种VwTyDC-like与VwTyDC基因的表达模式进行了分析。结果显示,在有斑品种黄色带花斑中,VwTyDC-like与VwTyDC在花发育S7 期的中心色斑区(a)的表达量显著高于非色斑区(b)(图5),这与前人的研究结果一致[13]。为探究VwTyDC-like与VwTyDC基因在有斑品种中心色斑区与非色斑区的表达差异是否源于位置效应,本研究还检测了在花瓣基部与边缘都有色斑分布的品种紫罗兰白双色VwTyDC-like与VwTyDC基因的表达水平。结果发现,2 个基因在非色斑区的表达量虽然低于基部的中心色斑区,但高于边缘色斑区(图6)。
进一步将白色与白色带花斑品种的唇瓣划分为5个区域进行分析,结果显示,在有斑品种和无斑品种中,VwTyDC-like与VwTyDC基因在唇瓣中的表达水平均由内向外依次降低(图7),说明它们的表达存在明显的位置效应。为验证VwTyDC-like与VwTyDC基因表达的位置效应在其他三色堇品种中是否同样存在,分析了2个基因在三色堇不同系列不同色系品种中的表达模式,结果发现在不同花色品种中,不论三色堇花瓣是否带有花斑,二者在中心色斑区或其对应区的表达量均显著高于非色斑区或其对应区(图8),进一步说明VwTyDC-like与VwTyDC基因表达的位置效应在三色堇品种中广泛存在。至于VwTyDC-like与VwTyDC基因在花瓣上表达具有位置效应的原因,仍有待进一步研究。
3.3 VwTyDC-like 和VwTyDC 基因与三色堇色斑形成的可能关系
已有研究表明,植物花斑形成的本质是花色素合成及调控基因在花器官(特别是花瓣或花萼)的特定区域差异表达的结果[25-27]。前人对拟南芥进行研究发现,TyDC的底物酪氨酸以一种剂量依赖型的方式有效诱导花青素的生物合成,主要通过上调花青素合成途径中的下游基因如DFR、LODX和UF3GT的表达量以及MYB-bHLH-WD40转录复合体中PAP1/MYB75、PAP2、GL3、EGL3、TTG1基因的表达促进花青素的积累[12]。Ma等[28]对茶叶品种Huabai 1的研究发现,酪氨酸与类黄酮化合物的积累有关。Cui等[2]用酪氨酸处理三色堇花瓣非色斑区,发现非色斑区出现了条纹状色斑,通过VIGS技术沉默VwTyDC基因后,三色堇花瓣的色斑颜色明显变浅,这些结果说明酪氨酸和VwTyDC基因确实与三色堇花青素的合成有关联。本研究通过基因表达模式分析,发现VwTyDC-like与VwTyDC的表达模式与三色堇花瓣中花斑的分布可能没有直接对应关系。由此推测在三色堇花斑形成过程中,酪氨酸脱羧酶基因还需要和其他因子联合发挥作用,而无斑品种缺乏这些相关因子。有研究发现,三色堇花斑由S和K两个基因控制[29],酪氨酸脱羧酶基因可能是其中一个基因,需要和另一个基因共同发挥作用。但是,另一个基因尚未被克隆,二者如何联合发挥作用还有待进一步研究。
4 结论
本研究从三色堇花瓣中克隆到一个VwTyDC-like基因,与VwTyDC同源性最高。VwTyDC-like与VwTyDC基因在三色堇不同组织器官、花发育不同时期、不同花色品种及其唇瓣不同区域表现出高度一致的表达模式,但VwTyDC-like基因的表达水平普遍高于VwTyDC。VwTyDC-like与VwTyDC基因的表达均具有组织特异性,在花瓣中的表达量远远高于其他组织器官,且在花发育后期(S7)表达量最高。无论三色堇花瓣是否带有花斑,VwTyDC-like与VwTyDC基因的表达量均由花瓣基部向外逐渐降低。本研究结果表明VwTyDC-like与VwTyDC基因在花瓣中的表达具有明显的位置效应。
