不同蓝莓品种对低温处理的生理响应及抗寒性评价
2023-09-04王兴东杨玉春刘有春
魏 鑫,王 升,王兴东,杨玉春,刘有春,刘 成
(辽宁省果树科学研究所,辽宁 营口 115009)
蓝莓学名越橘,属于杜鹃花科(Ericaceae)越橘属(Vaccinium),果实中富含花青素等营养保健功能成分,是具有很高经济价值和广阔开发前景的新兴小浆果果树,目前成为全球发展最快的果树树种[1-3]。由于我国现在栽培的多数蓝莓品种来源于美国、加拿大等地,其适宜栽培区域气候条件较为温和,即使冬季温度较低但空气湿度相对较高。我国东北地区冬季严寒、气候干燥,种植蓝莓时,冬季需进行防寒保护才能安全越冬,不仅增加了生产成本与工作量,而且在防寒过程中树体易受损伤,造成减产。蓝莓的抗寒性问题已成为限制该产业持续发展的主要因素之一[4]。因此,鉴定、评价蓝莓品种的抗寒性与分析抗寒机制对于抗寒品种的筛选和挖掘利用、栽培区域的合理布局与规划设计、防寒措施的制定等均具有重要意义。
目前蓝莓抗寒性研究主要从品种类型的抗寒性差异,质膜透性、组织结构、组织含水量、膜脂过氧化、保护酶系统等抗寒生理机制以及功能基因挖掘等方面进行的[5]。研究表明,随着温度的降低,抗寒性强的品种相对电导率的变化相对平缓,枝条中果糖和葡萄糖浓度与抗寒性水平呈显著正相关,丙二醛(MDA)含量与抗寒性呈负相关[4,6-8]。休眠期蓝莓花芽的电解质渗出率、渗透调节物质含量以及过氧化物酶(POD)活性总体上呈现规律性变化[9]。脯氨酸的积累有助于增强蓝莓抵御严寒的能力,保护酶活性、可溶性糖和脱落酸含量与越冬过程中抗寒能力的提高具有密切关系[10]。植物的抗寒机制非常复杂,将多种抗寒指标结合起来系统的分析和评价,才能更客观地反映蓝莓的抗寒性。而目前围绕蓝莓的抗寒性研究大多是初步探讨,并未将各项抗寒指标建立有效的联系和进行综合分析。因此,以抗寒性存在差异的不同品种蓝莓为试材,通过田间调查和人工模拟低温环境相结合的方法开展抗寒性研究,通过对质膜透性、MDA 含量、保护酶活性、渗透调节物质含量、纤维素含量、总含水量、自由水及束缚水含量等抗寒相关指标的综合分析和评价,将各项抗寒指标建立有效的联系,揭示受低温胁迫后各项指标的协同作用,以期明确蓝莓树体受冻后的生理响应机制和抗寒品种的抗性机制。
1 材料和方法
1.1 试验材料
供试品种5 个,分别为7 年生矮丛品种美登、半高丛品种北陆、北高丛品种蓝丰、南高丛品种密斯梯和兔眼品种精华。供试材料定植于辽宁省果树科学研究所蓝莓资源圃塑料大棚内,采取常规管理措施。
1.2 试验方法
1.2.1 取样与材料处理 于2016 年1 月10 日选取经过抗寒锻炼的5个品种植株上不同方向且完全木质化直径为0.3~0.5 cm 的1 年生枝条。将枝条分成三部分,一部分枝条用于不同低温处理,处理为-15、-20、-25、-30、-35、-40 ℃及对照(4 ℃冰箱冷藏保存),冷冻时温度下降幅度和解冻时的温度回升幅度都为5 ℃∕h,冷冻到达处理所需温度后,维持24 h,然后解冻。测定相对电导率、MDA 含量、保护酶活性和游离脯氨酸含量。一部分枝条于120 ℃烘箱杀青20 min 后,80 ℃烘干至恒定质量,粉碎成粉末后用于测定可溶性总糖、还原糖、淀粉、可溶性蛋白、糖组分以及纤维素含量。一部分枝条用于测定枝条总含水量、自由水含量并计算束缚水含量、自由水含量∕总含水量和束缚水含量∕总含水量。每处理重复3次,每次重复10个枝条。
1.2.2 测定指标与方法 相对电导率采用电导率仪法测定[11];MDA 含量采用TBA 显色法测定[12];过氧化物酶(POD)活性采用愈创木酚法测定[12];超氧化物酶(SOD)活性采用氮蓝四唑光还原法测定[12];游离脯氨酸含量采用硝基水杨酸法测定[12];可溶性糖、还原糖、淀粉、纤维素含量采用蒽酮比色法测定[12-13];可溶性蛋白含量采用考马斯亮蓝比色法测定[12];糖组分与含量采用高效液相色谱法测定[14-15];总含水量、自由水和束缚水含量采用烘干称重法和阿贝折射仪测定[12]。
1.3 统计分析
采用Excel 2007 软件进行数据预处理,采用Origin 2018制图,应用SPSS 20.0软件进行单因素方差分析和主成分分析,采用Duncan’s法检验差异显著性。
2 结果与分析
2.1 低温处理对5个蓝莓品种枝条相对电导率的影响
在试验低温范围内,5 个品种蓝莓相对电导率随处理温度的降低逐渐升高并呈S形变化。不同品种相对电导率随温度降低变化的速率有所不同。美登、北陆和蓝丰品种相对电导率随温度的降低变化较为平缓。密斯梯和精华2 个品种在-25 ℃后相对电导率明显提高。-40 ℃时,美登、北陆、蓝丰、密斯梯和精华5 个品种相对电导率较4 ℃时分别增加了1.02、1.20、1.17、2.56、2.45倍(图1A)。

图1 低温处理对5个蓝莓品种枝条相对电导率的影响Fig.1 Effect of different low temperature on relative electrical conductivity in five varieties of blueberry shoots
4 ℃条件下,5 个品种枝条相对电导率最低,密斯梯相对电导率显著高于精华、美登,与蓝丰、北陆差异不显著;密斯梯、蓝丰和北陆间差异不显著。-40 ℃处理下,5 个品种枝条相对电导率均最高,密斯梯相对电导率显著高于其他4 个品种,精华显著高于蓝丰、美登和北陆,蓝丰与北陆间差异不显著,但均显著高于美登(图1B)。
2.2 低温处理对5个蓝莓品种枝条MDA含量的影响
在试验低温范围内,5 个品种MDA 含量随处理温度的降低逐渐升高并呈S 形变化。4 ℃时美登枝条MDA 含量显著低于其他温度,-40 ℃时显著高于其他温度;4 ℃时北陆、蓝丰枝条MDA 含量与-15 ℃差异不显著,但显著低于其他温度,-40 ℃时显著高于其他温度;4 ℃时密斯梯枝条MDA 含量显著低于其他温度,-40 ℃时显著高于其他温度,-20 ℃、-25 ℃和-30 ℃之间差异不显著;4 ℃时,精华枝条MDA 含量与-15 ℃差异不显著,但显著低于其他温度,由-20 ℃开始,随温度的降低,枝条MDA 含量显著升高(图2A)。
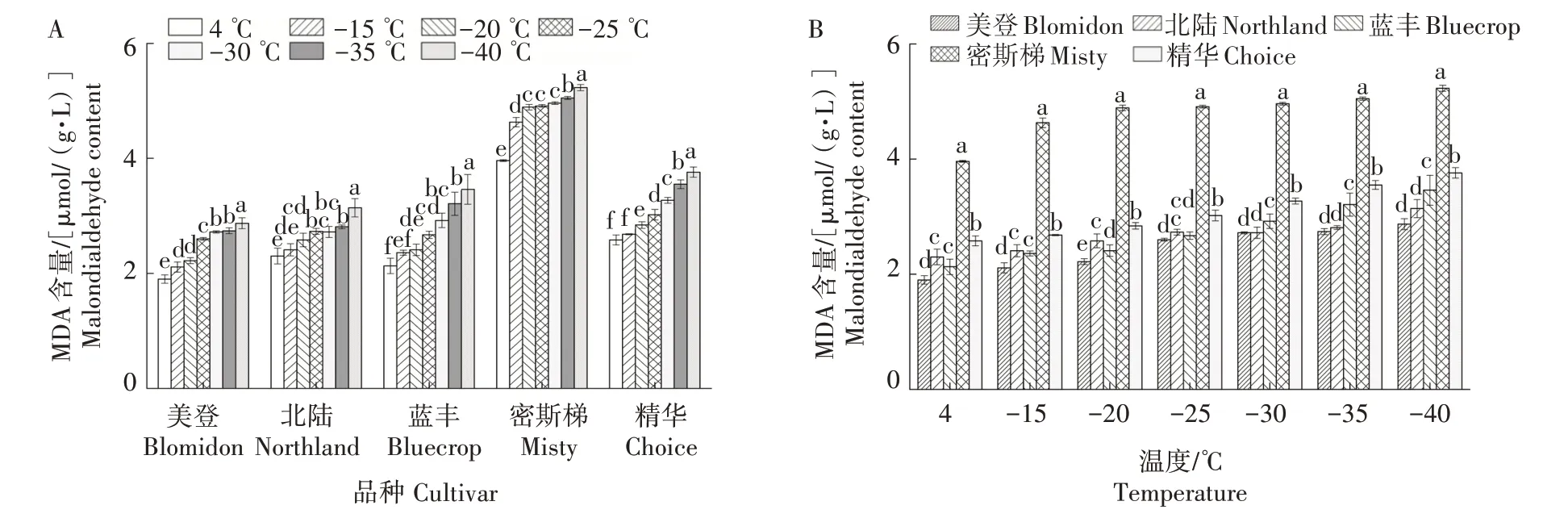
图2 低温处理对5个蓝莓品种枝条MDA含量的影响Fig.2 Effect of different low temperature on malondialdehyde content in five varieties of blueberry shoots
在各温度处理条件下,密斯梯MDA含量均显著高于其他4 个品种,精华显著高于其他3 个品种。4 ℃和-15 ℃时,北陆与蓝丰间差异不显著但均显著高于美登;-30、-35、40 ℃时,蓝丰显著高于北陆和美登,美登和北陆间差异未达显著水平(图2B)。
2.3 低温处理对5个蓝莓品种枝条POD活性的影响
5个品种POD活性在试验温度范围内均呈单峰变化趋势,但出峰温度有所不同。美登、北陆和蓝丰3 个品种在-30 ℃时,POD 活性达到最高,显著高于其他温度。密斯梯和精华2 个品种在-25 ℃时,POD 活性达到最高,显著高于其他温度。不同温度下,美登枝条POD 活性由高到低依次为-30、-25、-20、-35、-40、-15、4 ℃,各温度间差异均达显著水平;不同温度下,北陆蓝莓枝条POD 活性由高到低依次为-30、-25、-35、-20、-40、-15、4 ℃,除-35 ℃和-25 ℃间差异不显著外,其余温度间差异均达显著水平;4 ℃和-40 ℃时,蓝丰枝条POD 活性显著低于其他温度,两者间差异不显著;-25、-20、-30 ℃时,密斯梯枝条POD活性显著高于其他温度,-25 ℃显著高于-20 ℃和-30 ℃,-20 ℃显著高于-30 ℃;精华枝条POD 活性由高到低依次为-25、-35、-20、-30、-15、-40、4 ℃,各温度间差异均达到显著水平(图3A)。

图3 低温处理对5个蓝莓品种枝条过氧化物酶活性的影响Fig.3 Effect of different low temperature on peroxidase activity in five varieties of blueberry shoots
在各温度条件下,美登POD 活性显著高于其他4个品种,北陆和蓝丰均显著高于密斯梯和精华;除在-30 ℃时北陆和蓝丰间差异不显著外,其他低温条件下北陆均显著高于蓝丰;在-25 ℃条件下,密斯梯和精华间差异不显著,其他低温条件下精华均显著高于密斯梯(图3B)。
2.4 低温处理对5个蓝莓品种枝条SOD活性的影响
在试验温度范围内,5 个品种SOD 活性均呈单峰变化趋势,但出峰温度有所不同。美登、北陆和蓝丰3 个品种在-30 ℃时,SOD 活性达到最高,显著高于其他温度。密斯梯和精华2 个品种在-25 ℃时SOD 活性达到最高,显著高于其他温度。4 ℃时,除密斯梯枝条SOD 活性与-40 ℃差异未达显著水平外,其余4 个品种均为显著低于其他温度。-40 ℃时,美登、蓝丰枝条SOD 活性与-15 ℃差异不显著,但显著低于-20、-25、-30、-35 ℃。-40 ℃时,北陆、密斯梯和精华3 个品种枝条SOD 活性显著低于-15、-20、-25、-30、-35 ℃(图4A)。

图4 低温处理对5个蓝莓品种枝条SOD活性的影响Fig.4 Effect of different low temperature on superoxide dismutase activity in five varieties of blueberry shoots
4 ℃条件下,美登、北陆和蓝丰显著高于密斯梯和精华,美登、北陆、蓝丰以及密斯梯和精华间差异不显著;-25 ℃条件下,美登显著高于北陆、密斯梯和精华,与蓝丰间差异不显著,蓝丰显著高于密斯梯和精华,与北陆间差异不显著;-30 ℃条件下,美登显著高于其他4 个品种,北陆和蓝丰显著高于密斯梯和精华,北陆、蓝丰以及密斯梯和精华间差异不显著;-40 ℃条件下,美登显著高于北陆、密斯梯和精华,与蓝丰间差异不显著,蓝丰显著高于密斯梯和精华,与北陆间差异不显著(图4B)。
2.5 低温处理对5个蓝莓品种枝条渗透调节物质及纤维素含量的影响
在试验低温范围内,5 个品种游离脯氨酸含量随处理温度的降低呈逐渐上升的变化趋势。4 ℃时,美登枝条游离脯氨酸含量与-15 ℃和-20 ℃差异不显著,但显著低于其他温度;北陆枝条游离脯氨酸含量与-15 ℃差异不显著,但显著低于其他温度;蓝丰、密斯梯和精华3 个品种枝条游离脯氨酸含量均显著低于其他温度。-40 ℃时,5个品种枝条游离脯氨酸含量均显著高于其他温度(图5A)。

图5 低温处理对5个蓝莓品种枝条游离脯氨酸含量的影响Fig.5 Effect of different low temperature on free proline content in five varieties of blueberry shoots
4 ℃条件下,美登枝条游离脯氨酸含量显著高于另外4 个品种,蓝丰、北陆显著高于密斯梯,但与精华差异不显著;-40 ℃条件下,北陆枝条游离脯氨酸含量显著低于美登,显著高于蓝丰、密斯梯和精华,蓝丰、精华和密斯梯间差异未达显著水平(图5B)。
5 个品种可溶性总糖、还原糖含量由高到低依次为北陆、美登、蓝丰、精华和密斯梯,5个品种的可溶性总糖、还原糖含量差异均达显著水平。北陆品种淀粉含量显著高于另外4 个品种,美登显著高于蓝丰、密斯梯和精华,蓝丰和精华间差异未达显著水平,但均显著高于密斯梯。北陆、美登2个品种的可溶性蛋白含量显著高于其他3 个品种,北陆和美登间差异未达显著水平;蓝丰与精华间差异未达显著水平,但均显著高于密斯梯。美登、北陆、蓝丰3个品种的果糖含量差异未达显著水平,但显著高于密斯梯和精华,密斯梯和精华间差异达显著水平。北陆品种葡萄糖含量显著高于密斯梯和精华,与美登和蓝丰间差异未达显著水平;美登和蓝丰显著高于密斯梯,与精华差异未达显著水平;密斯梯和精华间差异未达显著水平。美登品种蔗糖含量显著高于其他4 个品种,密斯梯显著低于其他4 个品种,北陆显著高于蓝丰和精华,而蓝丰和精华间差异未达显著水平。北陆纤维素含量显著高于其他4个品种;美登显著高于密斯梯和精华,与蓝丰差异不显著;蓝丰显著高于精华,与密斯梯差异不显著;密斯梯与精华间差异不显著(表1)。
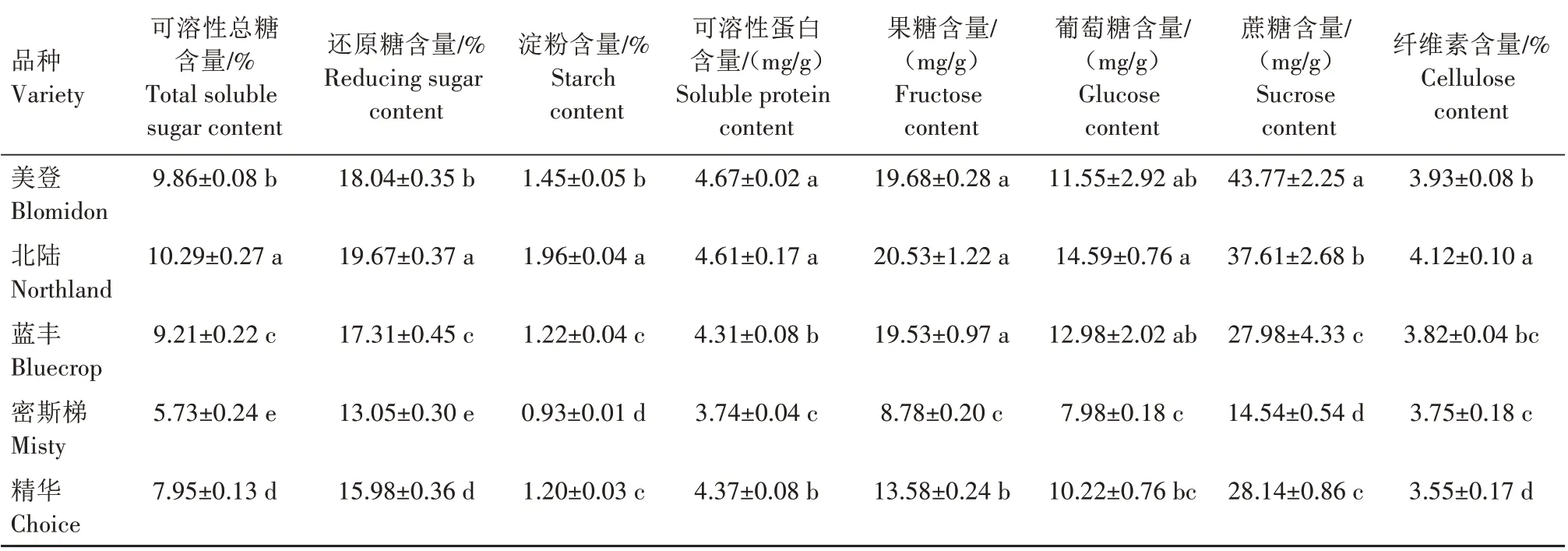
表1 5个蓝莓品种枝条渗透调节物质及纤维素含量差异Tab.1 Difference of osmotic regulator and cellulose content in five varieties of blueberry shoots
2.6 5个蓝莓品种枝条含水量的差异
密斯梯品种枝条总含水量显著高于另4 个品种,精华、蓝丰和北陆之间差异不显著但均显著高于美登;密斯梯品种枝条自由水含量显著高于另4个品种,蓝丰和精华2 个品种显著高于美登和北陆,美登、北陆间差异未达显著水平;北陆、蓝丰和精华束缚水含量显著高于美登和密斯梯,美登显著高于密斯梯;密斯梯自由水含量∕总含水量显著高于另外4 个品种,4 个品种之间差异未达显著水平;密斯梯束缚水含量∕总含水量显著低于另外4 个品种,4个品种之间差异未达显著水平(图6)。
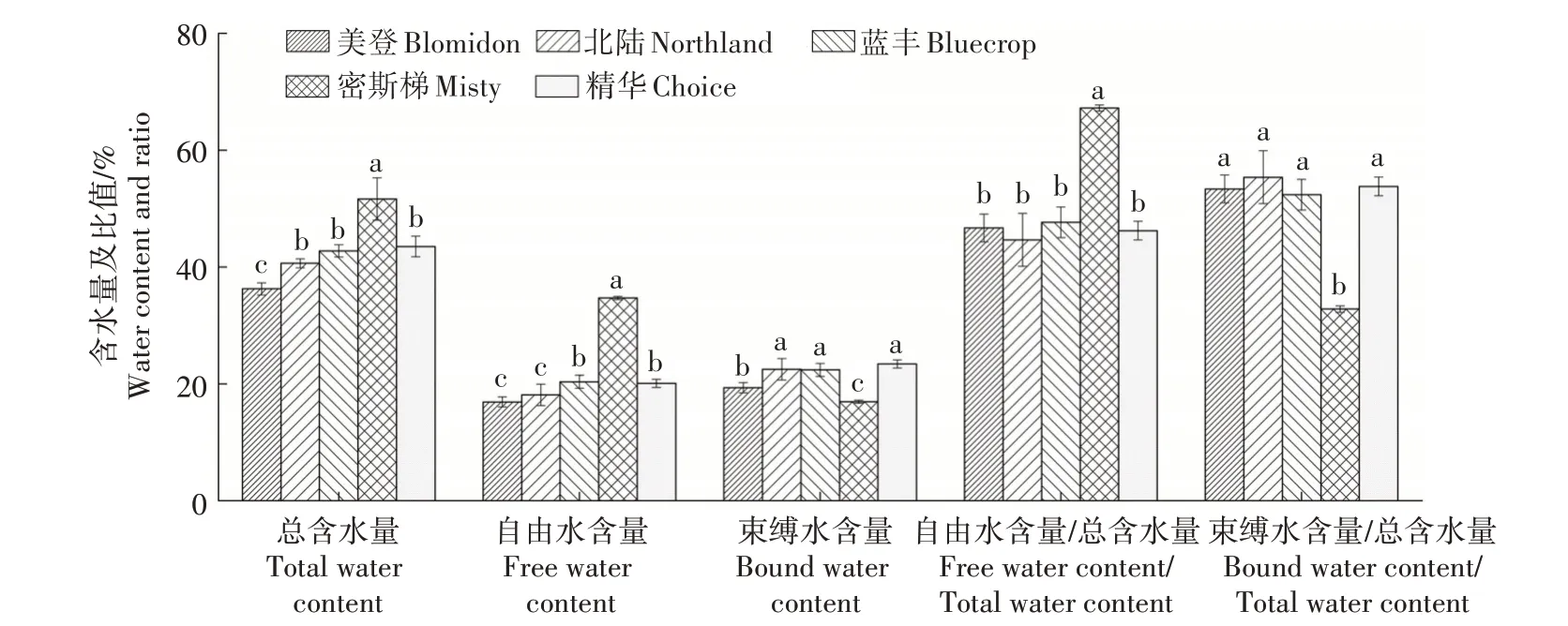
图6 5个蓝莓品种枝条含水量的差异Fig.6 Difference of water content in five varieties of blueberry shoots
2.7 蓝莓抗寒性的综合分析
对美登、北陆、蓝丰、密斯梯和精华5 个蓝莓品种的18 个抗寒生理指标进行主成分分析。前3 个主成分方差累计贡献率达97.632%(表2)。表明这3个主成分可以反映绝大部分原始信息,因此,可以用这3个主成分对5 个蓝莓品种的抗寒性进行综合分析。
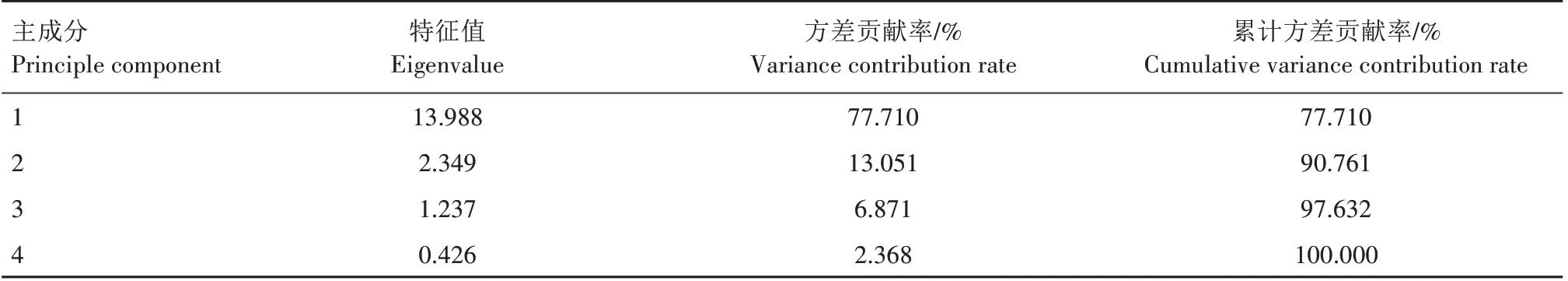
表2 不同低温处理蓝莓抗逆生理指标的特征值及方差贡献率Tab.2 Eigenvalues and variance contribution rates of physiological indexes of stress resistance for blueberry under different low temperature

表3 主成分在各抗逆指标上的载荷矩阵Tab.3 The loading matrix of principle components on each resistance index
前3个主成分的各指标在蓝莓抗寒能力中所起的作用有所不同。第1 主成分的特征值为13.988,方差贡献率为77.710%,SOD 活性、相对电导率和MDA、游离脯氨酸、可溶性蛋白、淀粉、纤维素、还原糖、可溶性总糖、果糖、葡萄糖、蔗糖、总含水量、自由水含量以及自由水含量∕总含水量、束缚水含量∕总含水量载荷值较大,其中MDA含量、相对电导率、总含水量、自由水含量和自由水含量∕总含水量为负向,其余为正向。若选择载荷值绝对值为0.95 以上,则相对电导率和MDA、可溶性蛋白、还原糖、可溶性总糖、果糖、自由水含量可以作为第1主成分的代表特征向量;第2 主成分的特征值为2.349,方差贡献率为13.051%,POD 活性、束缚水含量载荷值较大,POD 活性为正向,束缚水含量为负向,载荷值在0.78 以上;第3 主成分的特征值为1.237,方差贡献率为6.871%,具有较大载荷值的指标为淀粉和纤维素含量,载荷值在0.40以上(表2—3)。
对18个抗寒生理指标进行降维处理,使数据标准化并保留3 个主成分,用f1、f2 和f3 分别代表前3个主成分,Zx1—Zx18 分别代表MDA 含量等18 个指标消除量纲得到标准化的数据,利用主成分的特征值和因子载荷计算各综合指标系数,得到3 个主成分分值的表达式,如下:
主成分值1:f1=-0.26×Zx1+0.15×Zx2+0.24×Zx3-0.26×Zx4+0.21×Zx5+0.26×Zx6+0.22×Zx7+0.2×Zx8+0.26×Zx9+0.27×Zx10+0.26×Zx11+0.23×Zx12+0.25×Zx13-0.25×Zx14-0.25×Zx15+0.14×Zx16-0.24×Zx17+0.24×Zx18;
主成分值2:f2=0.08×Zx1+0.51×Zx2+0.02×Zx3-0.07×Zx4+0.41×Zx5+0×Zx6-0.06×Zx7+0.14×Zx8-0.07×Zx9-0.02×Zx10-0.01×Zx11-0.18×Zx12+0.16×Zx13-0.13×Zx14+0.11×Zx15-0.54×Zx16+0.24×Zx17-0.24×Zx18;
主成分值3:f3=0.19×Zx1-0.27×Zx2+0.13×Zx3-0.06×Zx4-0.09×Zx5-0.15×Zx6+0.38×Zx7+0.56×Zx8+0.17×Zx9+0.07×Zx10+0.12×Zx11+0.33×Zx12-0.12×Zx13+0.24×Zx14+0.23×Zx15-0.14×Zx16+0.21×Zx17-0.21×Zx18。
以所选取的第1—3 主成分的特征值和主成分值构建综合评价模型:F=(13.988×f1+2.349×f2+1.237×f3)∕(13.988+2.349+1.237),其中,F 为综合评价指标。计算得出5个品种不同低温处理的综合评价值F,根据F 值的大小判断抗寒性强弱,得分值越高则抗寒性越强(表4)。

表4 蓝莓不同低温处理下综合评价变量及F值Tab.4 The evaluation variables and F value of blueberry under different low temperature
5 个品种f1 由高到低依次为北陆、美登、蓝丰、精华和密斯梯,f2 由高到低依次为美登、密斯梯、北陆、蓝丰和精华,f3 由高到低依次为北陆、密斯梯、蓝丰、美登和精华;综合评价F值由高到低依次为美登、北陆、蓝丰、精华和密斯梯。由此可见,5个蓝莓品种的抗寒性由强到弱依次为美登>北陆>蓝丰>精华>密斯梯(表4)。
3 结论与讨论
3.1 不同蓝莓品种低温处理下质膜透性间的差异
在低温处理过程中,活性氧自由基积累,电解质外渗,MDA 大量积累,造成细胞膜系统的损伤[16-17]。本研究发现,在试验低温范围内,5 个品种枝条相对电导率和MDA 含量随处理温度的降低而呈逐渐升高的趋势。密斯梯和精华相对电导率和MDA 含量明显升高,美登、北陆和蓝丰相对电导率和MDA 含量升高的速度相对平缓,说明美登、北陆和蓝丰3个品种细胞膜系统受害程度低于密斯梯和精华。
3.2 不同蓝莓品种低温处理下保护酶活性间的差异
植物体内酶活性的变化能够反映植物在低温处理后受伤的程度,POD 和SOD 是植物体内重要的保护酶,与抗寒性有着密切的关系[18-19]。刘兴禄等[20]研究发现,5 个苹果砧木枝条的POD 和SOD 活性随着温度的降低呈先升后降的规律,抗寒性较强的品种受低温处理时酶活性增加幅度大。本研究发现,在试验低温范围内,5 个品种枝条POD、SOD 活性随处理温度降低呈单峰变化趋势,说明轻度低温处理下,参试品种通过提高酶活性来减轻低温处理的伤害。随着胁迫程度的增加,活性氧积累超过保护酶系统的清除能力,进而对抗氧化酶系统造成伤害导致其活性下降。密斯梯枝条酶活性的变化较另4个品种更为平稳,美登枝条酶活性的变化较另4 个品种更为迅速,且活性较高,可以快速清除超氧阴离子自由基和过氧化氢等活性氧,进而来维持细胞的稳定。
3.3 不同蓝莓品种渗透调节物质及纤维素含量间的差异
渗透调节作用是植物抵御逆境的重要生理机制,植物通过合成和积累脯氨酸、可溶性糖、可溶性蛋白等渗透调节物质提高细胞的渗透浓度,降低水势,增强植株的保水能力,从而提高抗寒性[21-22]。本研究发现,美登、北陆和蓝丰3个品种具有较高的可溶性总糖、还原糖、葡萄糖、果糖、蔗糖、淀粉、纤维素和可溶性蛋白含量,当受到低温处理时,其体内较高的渗透调节物质含量促使细胞渗透势下降,加强细胞吸水和保水能力,保证原生质不会脱水凝固,进而增强树体对低温的抗性。
3.4 不同蓝莓品种组织含水量间的差异
植物在遭受低温处理时,组织发生结冰,造成细胞死亡,因此,避免细胞内结冰是植物抗寒性的重要机制,植物的抗寒能力与体内的水分变化息息相关。束缚水有助于保持原生质胶体及大分子结构的稳定性,有利于植物抗寒性的增强[23-24],自由水占总含水量百分比越大则代谢越旺盛,抗性降低[25]。抗寒性强的桃品种可大幅提高束缚水与自由水比值以应对低温胁迫[26]。番茄能通过降低自由水含量、增加束缚水含量来降低自身代谢活性,减轻低温伤害[27]。美登由于具有较低的总含水量、自由水含量和自由水含量∕总含水量进而增强树体的抗寒性。而密斯梯由于具有较高的总含水量、自由水含量和自由水含量∕总含水量以及较低的束缚水含量∕总含水量,树体的抗寒性较差。
3.5 主成分分析综合评价抗寒性
果树的抗寒性是受多种因素共同影响的复杂生理生化过程,使用单一指标评价样本抗寒性时,难以全面客观地反映植物抗寒性的强弱,所以应进行抗寒性的综合评价[26,28-29]。主成分分析是一种可以综合多个指标的贡献值来判断抗寒性的综合评价方法,能够可靠地反映植物的抗寒性[30]。因此,在本研究中运用主成分分析的方法结合18 个指标对5 个不同类型的蓝莓品种进行抗寒性的综合评价。可以将相对电导率,MDA、可溶性蛋白、还原糖、可溶性总糖、果糖、自由水、束缚水含量和POD 活性作为评价蓝莓品种抗寒性强弱的指标。通过综合分析可确定供试5个蓝莓品种的抗寒性由强到弱依次为美登、北陆、蓝丰、精华和密斯梯。
综上所述,可以将相对电导率,MDA、蛋白质、还原糖、可溶性总糖、果糖、自由水、束缚水含量和POD 活性作为评价蓝莓品种抗寒性强弱的指标。综合分析可确定供试5个蓝莓品种的抗寒性由强到弱依次为美登、北陆、蓝丰、精华和密斯梯。美登抗寒性较强的原因是具有较高的POD 和SOD 活性,渗透调节物质含量以及较低的总含水量、自由水含量和自由水含量∕总含水量。
