植物群落邻体间的系统发育关系是否影响邻体效应?
2023-08-30王丽萍乌俊杰陈凤仙于小莉孙继文
王丽萍,乌俊杰,陈凤仙,冯 哲,于小莉,孙继文
(大理大学农学与生物科学学院,云南 大理 671003)
邻体效应是维持植物群落构建的主要驱动机制之一[1],当相邻植物个体发生相互作用时,竞争抑制和促进通常同时存在[2]。邻体竞争会强烈影响植物生长与种群更新,即相对某植物个体而言,其周围同种或异种邻体为争夺资源而对目标物种产生不利影响[3]。最近的研究表明,密度制约存在于亲缘关系较近的异种个体之间[4—5],即系统发育密度制约,是密度制约的延伸[6—8]。不同的异种植物对目标植物的影响不相同,这种差异源于邻体与目标种在谱系上的距离[6,9]。
当目标物种与邻体的亲缘关系较近,对其生长、存活或更新等不利时,为系统发育负密度制约(Phylogenetic negative density dependence, PNDD)[6]。研究表明,系统发育关系较近的物种拥有更多共同的病原体和昆虫食草动物[5,10—11],也更容易杀死亲缘关系较近的种子、幼苗和幼树[6,12],以至于近缘物种之间难以共存[5]。
在少数研究中,系统发育关系较近的邻体间表现另一种共存模式,即近缘异种邻体有利于目标物种生长,而远亲邻体也可能会抑制目标植物的生长从而不利于存活,表现为系统发育正密度制约(Phylogenetic positive density dependence, PPDD)[9]。生境过滤可能导致系统发育相似性和幼苗存活率之间存在明显的正相关关系,密切相关的植物通常具有相似的生境条件需求[13—14],即环境偏好相似[9],从而更可能聚集在一起,导致群落谱系聚集[15]。当生境异质性得到控制时,密切相关的异种邻体的幼苗存活率也更高[16]。这进一步反映出系统发育距离较近的物种可能在空间上更聚集并局部适应共同的生物因素或非生物环境[17]。
然而,系统发育方法的应用可能会受到限制,邻体效应的机制可能取决于植物的其他特征,而不是物种本身的系统发育相关性,即邻体间无系统发育密度制约,邻体间的相互作用强度主要是由层次结构驱动[18]。由于研究系统发育相关性在驱动竞争结果中受到分类群宽度和生态环境影响的限制,使研究变得模糊。因此,Mahon 等[19]采用回归和模型选择的方法研究系统发育相关性对不同生态环境下竞争的重要性,表明功能相似,非系统发育相关性,可以解释竞争相关的群落构建,系统发育距离的远近对邻体效应没有影响。
关于邻体间系统发育关系对邻体效应的影响研究较多,但至今无统一定论。因此,本文通过文献检索、筛选、整合有关邻体间系统发育关系对邻体效应影响的文献,并分析所筛选文献中不同结果的原因,旨在明确系统发育距离对邻体效应的影响表现不一的生态学机制,进而为研究森林群落中邻体间系统发育距离对邻体效应的影响,揭示群落构建和物种共存过程的影响机制提供科学依据。
1 数据来源与研究方法
1.1 文献检索
在Web of Science、Google Scholar 和CNKI 等3个数据库搜索邻体间系统发育关系对邻体效应的影响的文献,检索时间为1980.01.01~2022.05.01。
在Web of Science 上利用标题、摘要和全文进行检索,关键词均为: (‘phylogen*’ or ‘relat*’) and(‘density depend*’ or ‘density-depend*’or ‘neighbour*’or ‘neighbor*’ or ‘competition’ or ‘survival’ or ‘growth’or ‘coexist*’ or ‘Janzen-Connell’ or ‘Janzen Connell’ or‘J-C’ or ‘mortality’ or ‘recruit*’) and (‘plant*’ or ‘tree*’or ‘forest*’ or ‘seed*’) ,检索到4230 篇。
在 Google Scholar 上检索的关键词:(‘phylogenetic’ or ‘phylogeny’ or ‘phylogenetically’ or‘phylogenetics’ or ‘relation’ or ‘relationship’ or ‘relative’or ‘relatedness’) and (‘density dependence’ or‘density dependent’ or ‘density-dependent’ or ‘densitydependence’ or ‘neighbourhood’ or ‘neighborhood’ or‘neighbor’ or ‘competition’ or ‘survival’ or ‘growth’ or‘coexist’ or ‘coexistent’ or ‘Janzen-Connell’ or ‘Janzen Connell’ or ‘J-C’ or ‘mortality’ or ‘recruitment’) and(‘plant’ or ‘tree’ or ‘forest’ or ‘seed’)。共检索到88 篇。
在CNKI 上检索式为:SU=(‘系统发育’+‘谱系’)AND SU=(‘密度制约’+‘邻体’+‘竞争’+‘存活’+‘生长’+‘Janzen-Connell’+‘Janzen Connell’+‘J-C’+‘共存’+‘死亡’+‘更新’),检索到75 篇。
总共检索到4393 篇文献,删除重复后得到4388篇(图1)。
1.2 文献筛选
在检索到的文献中,依次通过标题、摘要及全文筛选出包含邻体间系统发育关系对邻体效应影响的文献35 篇(图1)。根据文章模型中系统发育指数的参数值及显著性来判定邻体间系统发育是否会显著地影响邻体效应。筛选的文献缺少对照组,数据为单组率分析。
1.3 数据提取
利用Plot Digitizer (Web Plot Digitizer - Extract data from plots, images, and maps (automeris.io))工具提取文献中的数据。提取每一项研究中系统发育的距离对邻体间相互作用的影响强度,并且记录作者、年份、研究地点和位置、年均温度、年均降水量、研究物种、生活史阶段、研究时间、预测类型(物种的死亡、存活、竞争、共存、发芽)、预测因子(系统发育的距离、物种相关性、系统发育多样性等)。
1.4 系统发育指数类型
文献中包含的系统发育距离指数有10 种(表1)。
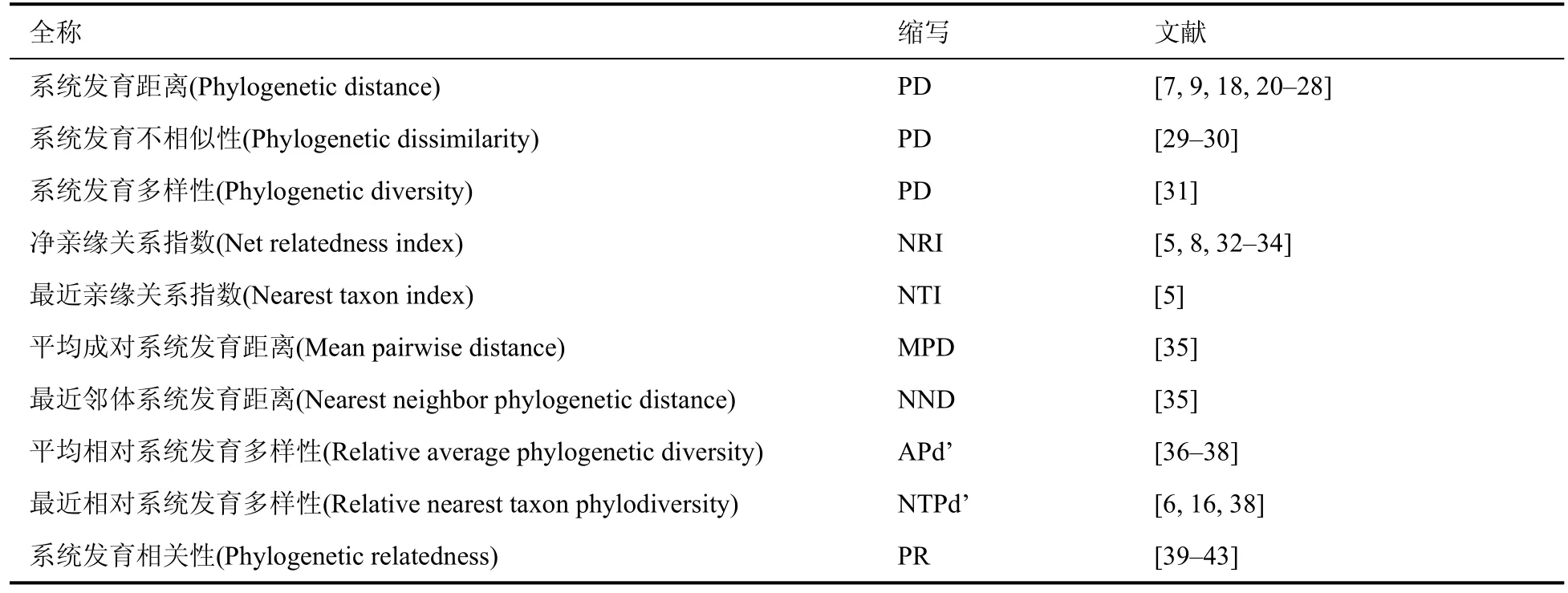
表1 邻体间系统发育指数Table 1 Phylogenetic indices of neighborhood
NRI、NTI、APd’ 和NTPd’ 的数值越大,代表邻体间的系统发育距离越近。其在模型中的参数为正时,说明邻体间系统发育距离越近,越不利于物种的存活、生长或更新,为系统发育负密度制约。若其在模型中的参数为负,则说明邻体间系统发育距离越近,越有利于物种的存活、生长或更新,为系统发育正密度制约。
PD、PR、MND 和NND 的单位为百万年(Ma),其数值越大表示邻体间的系统发育距离越远。在模型中的参数为正时,表明邻体间系统发育距离越远对物种间的相互作用越积极,为系统发育负密度效应;当参数为负时,表明邻体间系统发育距离越远越不利于物种的存活、生长或更新,为系统发育正密度制约。
2 结果与分析
2.1 研究地点分布
纳入分析的35 篇文献中涉及37 个研究地点(图2),主要分布在亚洲(15 篇)、美洲(11 篇)和欧洲(11 篇),且大部分研究地点分布在热带(16 篇)、亚热带(10 篇)和温带(6 篇)森林。

图2 研究地点地理分布Fig. 2 Geographical distribution of study sites
2.2 邻体间系统发育关系对密度制约的影响
检索到的文献中,最多的结果是PNDD (25 篇),其次是PPDD (10 篇),最少的是邻体间系统发育关系对邻体效应无显著作用(7 篇)(表2)。29 篇是在森林中进行野外实验,通过选定样地,建立样方,对幼苗或成树长期监测。有6 篇是室内实验条件下进行的[21—22,2527,43—44],且有两篇是同时进行野外和室内实验[27,43],其中一篇室内和野外的结果存在差异[27],在野外的结果为PNDD,在室内的结果则为PPDD,并且用于实验的对象为草本植物。仅有7 篇是无系统发育密度制约[18,21,25,28,33,42,44],包括3 篇室内实验[21,25,44]和4 篇野外实验[18,28—29,33]。

表2 邻体间系统发育关系对密度制约的影响Table 2 Effects of Phylogenetic relatedness of neighborhood on density dependence
在大尺度区域范围内,目标物种为成树时,邻体效应受到天敌、病原体和资源竞争因素影响。大尺度下热带幼苗表现出PNDD 主要由天敌、资源竞争和有害的隐性等位基因引起,而小尺度下幼苗只受到病原菌的影响。亚热带大尺度下的幼苗受到病原体、专一性天敌和资源竞争的影响,种子期受到天敌的影响;小尺度下的幼苗主要受天敌和资源竞争影响。在温带大尺度和小尺度下的幼苗分别受到病原体和资源竞争的制约(表3)。综上所述,导致PNDD 形成的原因与研究尺度和研究对象之间都没有明显规律。天敌、病原体、资源竞争和有害的隐性等位基因是导致PNDD 的重要原因。

表3 引起系统发育负密度制约的原因Table 3 Causes of phylogenetic negative density dependence (PNDD)
在大尺度和小尺度下生境过滤作用于热带、亚热带和温带地区的幼苗,同时大尺度下亚热带和温带地区的成树也受到该机制的制约;而生境异质性小和共生真菌具有特异性,只作用于大尺度下低纬度的幼苗(表4)。其中研究对象与导致PPDD 形成的原因表现出明显规律,即成树只受到生境过滤作用,而幼苗受到三种因子影响。除此之外,气候带和研究尺度与具体原因也存在一定的规律,但不明显。

表4 引起系统发育正密度制约的原因Table 4 Causes of phylogenetic positive density dependence (PPDD)
在亚热带和温带均检测到邻体间无系统发育密度制约的结果。由于部分文献没有解释具体原因,因此导致该结果的限制因子主要为性状无系统发育信号和早期物种间不会有直接的相互作用。当研究对象为大尺度空间的成树和幼苗时,主要是性状无系统发育信号;为小尺度种子时,主要是早期种子间没有直接相互作用(表5)。
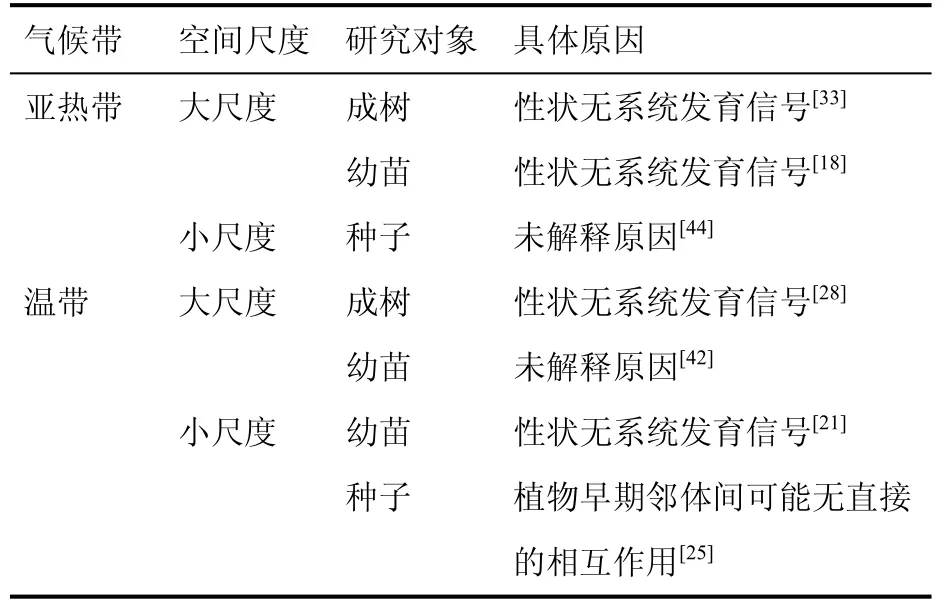
表5 邻体间无系统发育密度制约Table 5 Causes of not phylogenetic density dependence on neighborhood
在相同地点不同结果的文献中,研究地点分布在国内(黑石顶、天童山和长白山)和国外(Sepilok Forest Reserve、BCI 和Davis Bodega Marine Reserve)(表6)。在SFR 研究对象都为幼苗,部分幼苗存在有害的隐性等位基因导致PNDD,另一部分幼苗由于生长在生境异质性小的环境中进而出现PPDD。在BCI 研究结果不同于研究对象的生活史阶段,成树为PNDD,主要限制因子有资源竞争、天敌、病原菌和近亲不利繁殖;幼苗影响因素为生境过滤,表现出PPDD。黑石顶成树和幼苗阶段都检测到PNDD,主要由专一性天敌和病原体引起;但幼苗阶段还出现PPDD,其原因是与菌根真菌共生。在天童山,小树和灌木分布于下层受到资源竞争的影响,为PNDD;上层的成树由于环境过滤作用,结果为PPDD。长白山和Davis Bodega Marine Reserve不同的原因都与研究对象无关,其中长白山成树和幼苗都检测到PNDD 和PPDD 两种结果,成树和幼苗的PNDD 分别由天敌和病原体影响,而PPDD 都受到生境过滤的制约。在 Davis Bodega Marine Reserve 小尺度下研究对象都为幼苗,PNDD 和PPDD 分别由资源竞争和生境过滤导致。整体上,导致相同地点不同结果与研究对象总体不相关,在幼苗和成树阶段都检测到PNDD 和PPDD。
3 讨论
3.1 邻体间系统发育负密度制约
尽管许多相互并存的机制可能有助于森林中的植物物种共存,但对PNDD 稳定机制的研究主要集中在那些作为病原体的微生物和昆虫,种子捕食者和食草动物介导的负效应上[48]。从文献分析来看,形成PNDD 的三种主要因素在大尺度的成树中均有体现,可能在成树阶段能为其他生物提供生存环境,进而吸引各种生物来影响幼苗更新、幼树生长和树木死亡。而种子阶段可能受到“物种群体保护”,即高密度的异种苗可能会减少捕食者对种子的察觉,并增加种子到幼苗过渡的可能性[49]。低纬度带生物相互作用的强度作为物种聚集的驱动因素,使亲缘关系较远的物种聚集在一起[32]。已知异种邻体会减少宿主与其物种特异性害虫和病原体之间的接触,这种情况会减少天敌的传播,提高存活率[50]。在热带短时间和小的空间尺度上,幼苗物种多样性可能通过与病原体的相互作用,进而有利于周围其他幼苗生长[6]。而亚热带和温带的幼苗和种子阶段的PNDD 效应中没有受到病原菌影响,可能是其他因素的负效应强度超过病原菌。此外,性状和谱系的发散也可能有助于种间生态位分化,提高根系对病原菌的抵抗力,促进根系群落物种的稳定共存[37]。
因此,研究尺度的大小对研究结果影响不显著。但Hubbell 等[51]认为在大的空间尺度上,局部密度制约效应不影响森林群落多样物种的共存,然而小尺度上效应的积累有可能转移到大尺度上,从而影响群落结构,有助于多物种共存。在一些特殊环境中,空间资源的异质性可能掩盖潜在的PNDD 现象,即使在适合种群增长的环境中也难以检测到PNDD,而且还会产生PPDD 假象。
3.2 邻体间系统发育正密度制约
一开始PPDD 是一种尚未被认识和独立存在的现象,但近年来,这种现象不断在热带、亚热带和温带森林的各个生活史阶段被验证和认可,PPDD是真实存在的,即PPDD 也是影响植物生长和存活的一种因素[8—9,16,38]。
PPDD 主要通过较强的生境过滤[8—9,16]、环境异质性小[17]和共生真菌[43]三种机制促进邻体间近缘物种共存。生境异质性与物种生境偏好共同作用导致的生境过滤效应是物种生态位分化的一种形式[52],环境异质性有助于树木生长和发育的差异,包括获取资源、分散和承受压力的能力,还可以解释更多的密度制约机制。其中地形可影响光照、温度和水分分布,是为生态群落提供生物多样性的最重要的环境梯度之一[53];土壤是植物生存和生长的物质基础,土壤养分的空间异质性对生态群落有重要影响[54]。实际上,导致水和养分可用性差异的异质环境在自然界较常见,树木的生长和死亡与地形梯度的土壤资源相关[55]。例如,西双版纳大样地的生境因子主要是地形和土壤[16];台湾莲花池年均降水量超过2229 mm,关键的生境过滤限制因素不是土壤湿度,而是光照[38]。长白山地处高纬度和高海拔地区,温度和水分相对于热带和亚热带的森林较低,土壤湿度是该地带的限制因子[46]。在检索到的文献中,受到生境因子土壤理化性质[9,16—17,27,32,43,46]、地形[9,16—17,38]和光因子[8,41]影响的篇数依次递减,源于土壤理化性质是决定植物能否在其区域存活和生长的主要条件之一,还可能与部分植物对地形和光照不太敏感的特异性有关;光因子制约的文献较少,可能部分研究在实验过程未考虑光照的影响。
形成PPDD 的机制还与共生真菌有关,研究表明同属或同科的植物往往在植物各个器官里拥有共生微生物种群[56—58],其中外生菌根可能使邻近幼苗的亲缘关系增加[38]。菌根真菌外部的菌丝能够扩大宿主植物根的吸收面积,增强对营养的吸收,还能增强植物对病原菌的抗性[59]。
气候变化会影响树种的分布,导致生物多样性和生态系统功能的不同[60],进而促成局域群落间相互作用的差异。
3.3 邻体间无系统发育密度制约
邻体间系统发育关系无密度制约效应,即物种的生存与邻体的亲缘关系无关联,这与生态位理论相悖。最近研究表明,植物生长潜力而非系统发育关系能有效预测荒废农田土壤生物对外来和本土植物的作用[61],即无系统发育信号。邻体间缺乏强烈的系统发育密度制约可能反映检测的性状无系统发育信号。例如,在草本植物中,功能性状缺乏系统发育信号是相当常见的[62]。最大株高、叶磷含量、叶氮含量、种子平均质量和比叶面积都不是高寒草甸植物系统发育的保守性状[28]。一部分研究在室内做种子萌发实验,选择的目标物种处于种子阶段,而该阶段植物间可能没有直接的相互作用[25],进而导致邻体间无系统发育密度制约效应。此外,某些实验条件可能会模糊系统发育相关性、功能相似性和邻体间共存方式,如单独的系统发育距离并没有显著影响竞争,然而系统发育距离和胁迫条件之间的相互作用改变了植物性状和竞争[25]。
3.4 同一样地不同的邻体效应
同一样地的系统发育关系有不同的邻体效应,体现了生态位理论的两个相反过程(生境过滤和竞争排斥)同时存在。一方面,相同地点幼苗与幼苗邻体效应不同。首先,这是由幼苗的特性(稀有种和常见种)决定的,土壤微生物群对稀有种的影响具有显著的系统发育保守性[43],可能是由强病原体和弱菌根真菌共同感染造成的负作用[63]。此外,土壤有害生物更倾向于聚集在稀有物种附近,并产生不成比例的限制[64]。与稀有种相比,常见种通常被认为与菌根真菌有更强的关联,通过与微生物的互惠作用来促进生长[65]。其次,还与幼苗邻体的大小相关。大树遮蔽了较小的邻体,占据生根区和空间区的主导地位,并吸引大量的病原体和食草动物[66]对幼苗产生不利影响;幼苗与幼苗间获取的各种资源相当,受到环境过滤作用强度更大,有利于幼苗的存活[46]。另一方面,相同地点幼苗与成树的邻体效应不同。生活史阶段(成树与幼苗的生长差异),成树主要受到特定天敌(例如病原体、植食性昆虫等)的影响[5],以及种间对光和地下资源的竞争[8];而幼苗主要受到生境过滤的作用,系统发育距离较近的树种聚集分布[8—9]。此外,同一样地的不同邻体效应也源于物种的分布不同。在森林群落中林冠上下层邻体效应的差异归因于冠层结构驱动的森林群落地面以上的微气候差异,如林下光照强度、空气湿度等能随距离地面的距离而发生变化[67];上层大树能改变下层幼树或灌木的光照分布,进而改变林下物种多样性[68]。
4 结论
在不同研究中,邻体间系统发育关系对邻体效应的影响不一致,主要有三种结论:系统发育负密度制约效应、系统发育正密度制约效应和无系统发育密度制约。导致不同结论的原因可能在于不同的气候带、研究方法(野外或室内)、目标物种类型(稀有种和常见种)、植物生活史阶段(种子、幼苗、幼树和成树)以及环境异质性。因此,今后研究邻体间系统发育密度制约时,应尽可能考虑以上因素的影响,以确保研究结果的准确性和可靠性。
