水牛ACSL基因家族全基因组及表达分析
2023-08-16梁莎莎于农淇鄢胜飞卢瑛李厅厅黄健谭正准李辉黄荣春潘伟军覃广胜
梁莎莎 于农淇 鄢胜飞 卢瑛 李厅厅 黄健 谭正准 李辉 黄荣春 潘伟军 覃广胜
摘要 为鉴定水牛长链脂酞辅酶A合成酶(long-chain fatty Acyl-CoA synthetases,ACSLs)的家族成员并探究该基因家族在泌乳方面的功能,通过生物信息学技术对水牛ACSL基因家族进行了motif、基因结构分析、共线性分析、不同物种系统进化分析,运用qPCR技术检测ACSLs在不同组织的mRNA表达量。结果表明:水牛ACSL基因家族共有5个家族成员并分别命名为bbu.ACSL1、bbu.ACSL3、bbu.ACSL4、bbu.ACSL5和bbu.ACSL6。染色體分布情况结果显示:bbu.ACSL1位于1号染色体,bbu.ACSL3位于2号染色体,bbu.ACSL4位于25号染色体,bbu.ACSL5位于23号染色体,bbu.ACSL6位于9号染色体。这5个ACSL氨基酸数在699~722 aa,所有蛋白的等电点除ACSL6外均大于7.00。Motif分析发现,除ACSL3和ACSL4缺少motif8外,其余均含有预测出的Motif 1~10,且排序相同。物种内共线性分析结果显示,水牛和普通牛中均不存在串联重复和片段重复,种间共线性分析结果显示,水牛ACSL与普通牛ACSL存在4对片段重复基因。不同物种ACSL系统发育分析显示,ACSL基因家族具有较高的保守性,其中水牛与普通牛和牦牛的聚类更为相近。不同组织表达量分析发现ACSL1、ACSL3在乳腺组织的mRNA表达量高于其他组织,ACSL6在大脑的mRNA表达量高于其他组织,ACSL4在肺部的mRNA表达量高于其他组织,ACSL5在肝的mRNA表达量高于其他组织。根据RNA-seq 测序结果分析不同泌乳时期表达量结果显示,ACSL1在整个泌乳期均有高表达量,尤其是在泌乳第50、140和280天。ACSL3和ACSL5在泌乳第7天和第50天高表达,但随着泌乳天数增加,表达量降低。ACSL4和ACSL6在泌乳第7天表达量最高,但在整个泌乳期的表达量均远低于该家族的其他3个成员。
关键词 水牛; ACSL; 基因结构; 共线性; 系统发育;乳腺组织;表达量
中图分类号 S823.8+3 文献标识码 A
文章编号 0517-6611(2023)14-0079-07
作者简介 梁莎莎(1992—),女,广西南宁人,助理研究员,从事水牛分子遗传学研究。*通信作者,研究员,从事动物育种与繁殖研究。
收稿日期 2022-08-08
水牛以耐粗饲而著称,具有适应性强、耐高温高湿、抗病力强和易饲养等特点[ 1]。除了作为役畜外,水牛还可提供奶类和肉类,具有重要的经济价值[ 2]。水牛奶乳汁浓厚,奶质优良,营养丰富,具有较高的乳脂肪(8.0%)、乳蛋白(4.5%)、不饱和脂肪酸比例和较低的磷脂和胆固醇水平[ 3],有“奶中之王”之称。然而,水牛平均产奶量远低于荷斯坦普通牛,奶产量仅占世界牛奶产量的13%[ 4]。因此,提高水牛泌乳性能至关重要,而挖掘与产奶性状相关的候选基因有助于改善水牛泌乳性能。
长链脂酞辅酶A合成酶家族(longchain fatty Acyl-CoA synthetases,ACSLs)是哺乳动物利用脂肪酸活化催化生成酯酰辅酶A过程中所必需的酶[ 5]。目前已发现,哺乳动物ACSLs基因有5种亚型,分别为ACSL1、ACSL3、ACSL4、ACSL5和ACSL6[ 6-7]。研究表明,ACSLs基因在甘油三酯、磷脂和胆固醇合成及脂肪酸代谢中具有重要作用[ 8]。ACSL1基因可以提高甘油三酯中油酸的沉积,增加脂肪在细胞内的沉积量[ 5],对牛脂肪细胞中的多不饱和脂肪酸合成具有积极调控作用[ 9]。在中国红草原牛中,脂肪细胞分化过程诱导ACSL3,其过表达可促进脂滴甘油三酯含量的增加[ 10]。ACSL4被发现的功能包括细胞内脂质储存[ 11],胆固醇从内质网运输到线粒体[ 12],有研究发现,ACSL4多态性与不同猪品种的IMF含量和脂肪酸组成有关[ 13]。ACSL5存在于各种组织中,包括棕色脂肪组织、骨骼肌、肝脏和大脑,有发挥促进脂肪酸在合成代谢脂类和分解代谢β-氧化通路之间的通道作用[ 14]。ACSL6最初在大鼠大脑中被发现[ 15],ACSL6缺乏症会破坏正常的脑功能和精子发生,小鼠缺乏ACSL6会表现出运动功能障碍和星形胶质细胞增生增加[ 16]。
大量研究表明,ACSLs对哺乳动物的许多代谢过程中发挥着不可或缺的作用,特别是脂质代谢方面,然而目前关于水牛ACSL基因家族的研究较少。笔者以水牛基因组为参考,从全基因组水平鉴定水牛的ACSL家族成员,分析该家族成员的蛋白序列特征、motif分布、外显子-内含子结构、染色体定位、共线性关系、系统进化关系,运用qPCR和RNA-seq技术检测水牛ACSL基因家族在不同组织和不同泌乳时期的表达量差异,以期为后续深入挖掘水牛ACSL基因家族的功能提供理论基础。
1 材料与方法
1.1 材料
试验水牛组织cDNA(心、肝、肺、肾、胃、乳腺和大脑)均来自广西水牛研究所,品种为尼里水牛,月龄约48个月,体重约500 kg,雌性,泌乳期3头。
1.2 主要试剂和仪器设备 主要试剂 PerfectStart Green qPCR SuperMix购自北京全式金公司。主要仪器 NanoDrop2000紫外分光光度计(Gene,USA)和Roche Lightcycler 480实时荧光定量PCR仪。
1.3 试验数据 以水牛及其相关物种的全基因组序列为基础进行分析。全基因组数据包括全基因组的基因序列、蛋白质序列和基因注释文件。全基因组数据源自NCBI基因组数据库(表1)。
1.4 水牛ACSL基因家族成员蛋白序列的获得
登陆UniProt(https:∥www.uniprot.org/)搜索ACSL基因家族的蛋白质序列,勾选所有该家族不同物种的可靠蛋白序列,下载保存并使用MEGA 7.0软件比对其同源性。登陆NCBI下载水牛完整蛋白序列,再使用hmmbuild和hmmsearch软件构建HMM模型,并搜索序列库找到水牛ACSL基因家族所有的蛋白序列,最后使用TBtools软件将序列提取出来。
1.5 水牛ACSLs蛋白序列特征分析
使用ProtScale(https:∥web.expasy.org/protscale/)計算水牛ACSL家族蛋白质分子量和等电点;使用TBtools软件中的Table Row Extract or Filter插件从水牛和普通牛的基因注释文件中提取ACSLs染色体分布信息。
1.6 水牛ACSLs蛋白质保守基序分析及基因结构预测
使用MEME(http:∥memesuite.org/tools/meme)和GSDS(http:∥gsds.cbi.pku.edu.cn/)分析水牛ACSL家族蛋白质保守基序(motif)和基因结构;使用Pfam(http:∥pfam.xfam.org/)搜索每个motif所属的结构域;使用MEGA 7.0构建水牛ACSL家族系统进化树;最后使用TBtools软件将水牛ACSLs系统进化树和motif分析结果整合到一起进行可视化。
1.7 水牛和普通牛ACSLs共线性分析
为了探讨ACSL基因家族的进化进展,使用TBtools软件中的one step MCscanX插件对水牛和普通牛的种内及种间进行共线性分析,使用Advanced Circos和Dual Systeny Plot插件绘制物种内及种间共线性图。
1.8 不同物种ACSLs系统发育分析
为揭示水牛与其近缘物种ACSLs之间的进化关系,使用MEGA 7.0构建水牛、普通牛、牦牛、山羊、绵羊、马和骆驼的ACSL家族系统进化树,最后使用Adobe Illustrator CS6对系统进化树进行美化。
1.9 水牛ACSLs表达分析
1.9.1 不同组织表达量检测及分析。
使用Primer 3.0设计水牛ACSL基因家族qPCR引物,引物序列见表2。将水牛组织(心、肝、肺、肾、胃、乳腺和大脑)的cDNA 稀释至70~80 ng/μL,使用实时荧光定量PCR检测ACSL基因家族在不同组织中的表达情况,以GAPDH作为内参。反应体系为20 μL,其中2 x PerfectStart Green qPCR SuperMix 10.0 μL,Rnase water 8.2 μL,上下游引物各0.4 μL,cDNA 1.0 μL;反应条件:94 ℃ 预变性30 s,94 ℃ 5 s,58 ℃ 30 s,72 ℃ 10 s,40个循环。运用2-ΔΔCt法计算目的基因的相对表达量,采用Duncans新复极差法对不同组织表达量进行多重比较,P<0.05表示差异显著,P<0.01表示差异极显著。
1.9.2 基于RNA-seq进行不同泌乳时期表达分析。
为揭示ACSLs不同成员在水牛乳腺组织中不同泌乳时期的表达量差异,笔者选用4个泌乳时期:泌乳第7天(7 d)、第50天(50 d)、第140天(140 d)和第280天(280 d),每个时期2头,活体采集共8头摩拉水牛乳腺组织进行转录组测序,测序结果已上传到NCBI数据库,登录号:PRJNA480718(SRR7523532~SRR7523538)。
ACSLs表达量分析:使用sratoolkit将所有RNA-seq原始sra文件转换为fastq格式。运用kallisto构建水牛ACSL基因家族index库,并分别比对8个RNA-seq结果,进行量化分析,计算ACSLs在不同RNA-seq中的TPM(Transcripts Per Million)值,由TPM值来表示基因的表达量,计算每组的平均值后,使用TBtools软件中的Heatmap插件对其进行可视化,绘制表达量热图。
2 结果与分析
2.1 水牛ACSL蛋白序列的获得及特征分析
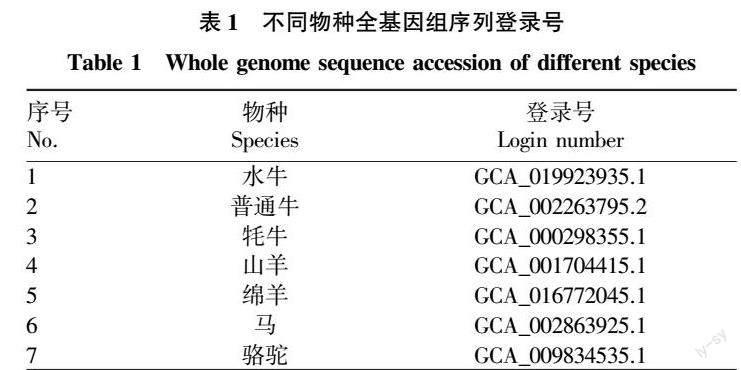
对水牛基因组的蛋白序列进行搜索,共鉴定得到5条水牛ACSL蛋白的编码基因,命名为bbu.ACSL1、bbu.ACSL3、bbu.ACSL4、bbu.ACSL5和bbu.ACSL6。染色体分布情况结果显示,bbu.ACSL1位于1号染色体,bbu.ACSL3位于2号染色体,bbu.ACSL4位于25号染色体,bbu.ACSL5位于23号染色体,bbu.ACSL6位于9号染色体。bbu.ACSL6的氨基酸数为722 aa,最短的是bbu.ACSL1,为699 aa。所有蛋白的等电点除了bbu.ACSL6,均大于7.00(表3)。
2.2 水牛ACSLs motif分析和基因结构预测
根据水牛ACSLs系统进化树结果,可将水牛ACSL家族基因分为3组,即ACSL1和ACSL6为一组,ACSL5单独为一组,ACSL3和ACSL4为一组。
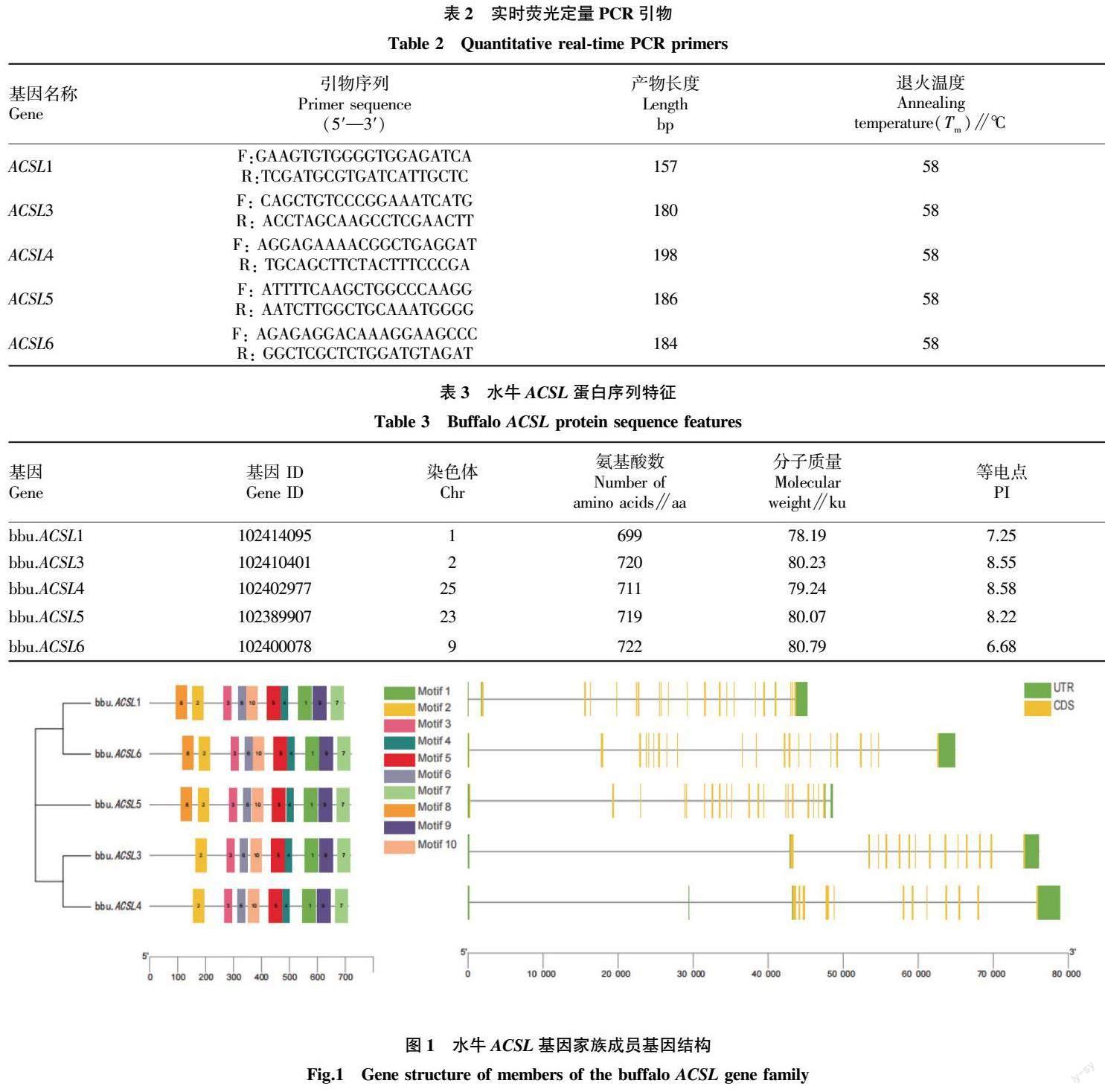
蛋白保守基序结果显示,共预测出10个保守的motifs(命名为Motif 1~10)。这10个保守motifs的分布见图1,氨基酸序列信息见表4。由图1可知,水牛ACSL家族成员中,除了ACSL3和ACSL4外缺少motif8外,其余均含有预测出的Motif 1~10且排序相同。使用Pfam分析每个motif后发现,除了motif7和motif8外,均含有AMP-binding结构域。基因结构结果显示,水牛ACSL1和ACSL5基因有23个外显子,22个内含子,ACSL3有15个外显子,14个内含子,ACSL4有18个外显子,17个内含子,ACSL6有24个外显子和23个内含子。
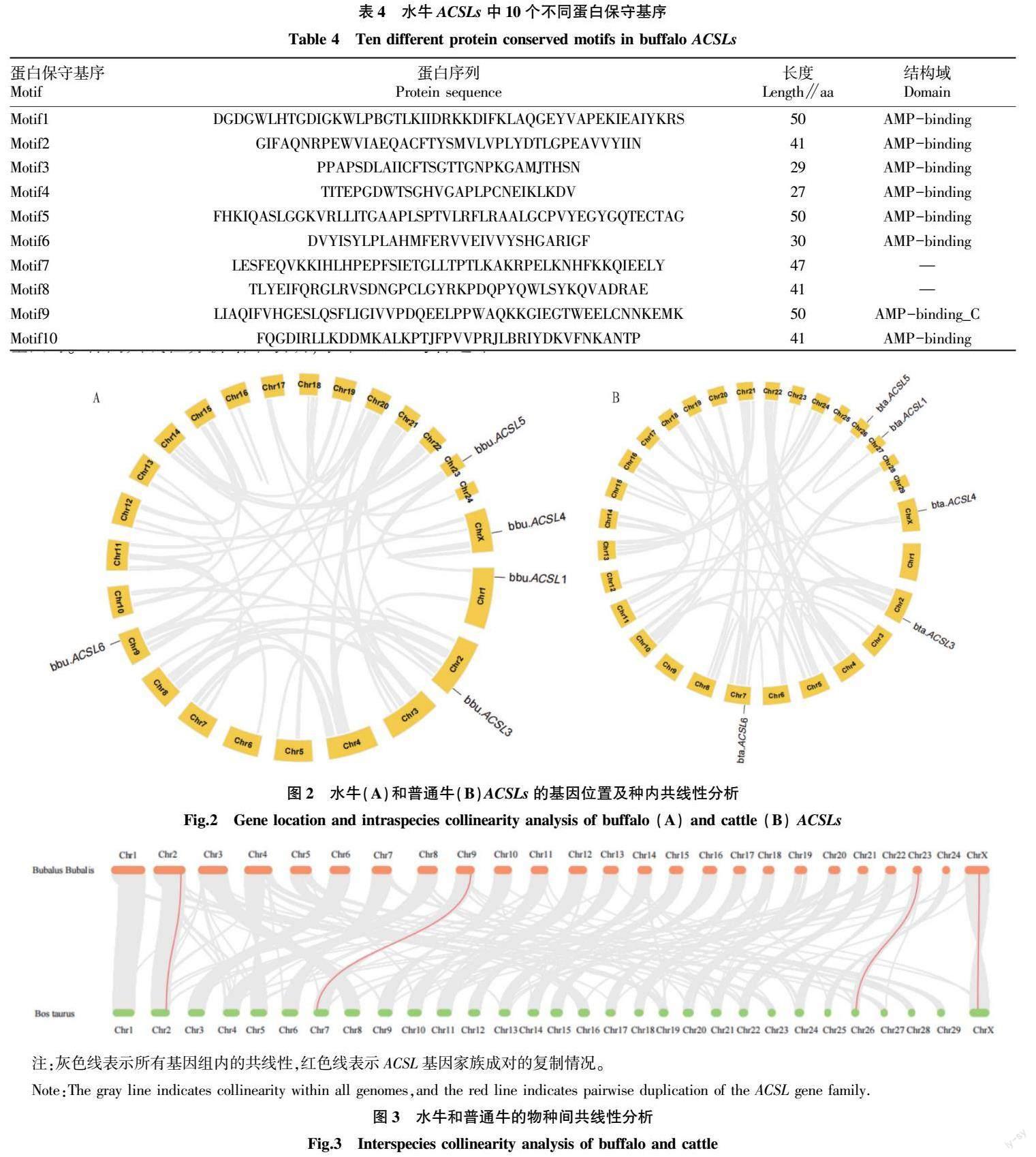
2.3 水牛和普通牛ACSLs共线性分析
为了研究ACSL家族的进化进展,对水牛和普通牛的种内及种间进行共线性分析(图2)。结果发现,在水牛和普通牛中均未发现串联重复基因对。种间共线性分析结果表明,水牛ACSL与普通牛ACSL存在4对片段重复基因,分别是bbu.ACSL3-bta.ACSL3、bbu.ACSL6-bta.ACSL6、bbu.ACSL5-bta.ACSL5和bbu.ACSL4-bta.ACSL4(图2、3)。
2.4 不同物种ACSLs系统发育分析
为揭示水牛与水牛近缘物种ACSLs之间的进化关系,分别获取了普通牛(Bos taurus)、牦牛(Bos mutus)、山羊(Capra hircas)、绵羊(Ovis aries)、马(Equus caballus)和骆驼(Camelus ferus)的ACSL家族蛋白序列,并构建了进化树(图4)。根据进化树信息,发现所有的基因划分为大类,分别是Group Ⅰ、Group Ⅱ和Group Ⅲ。Group Ⅰ 包括所有上述物种的ACSL1和ACSL6,Group Ⅱ只包括ACSL5,GroupⅢ包括ACSL3和ACSL4,其中水牛ACSL家族基因与普通牛和牦牛的聚类更为相近。
2.5 水牛ACSLs不同组织表达量分析
为了探究水牛ACSLs的组织特异性,利用qPCR技术检测了ACSLs在心、肝、肺、肾、胃、乳腺和大脑7个组织中的mRNA表达量,结果显示(图5),这5个ACSL基因在这7个组织中均有表达,其中ACSL1、ACSL3在乳腺组织中的mRNA表达量极显著高于其他组织(P<0.01),ACSL6在乳腺组织中的mRNA表达量极显著高于肺、肝、肾、胃、心、脂肪,ACSL3和ACSL6在大脑的mRNA表达量极显著高于肝、心、胃、心、脂肪组织(P<0.01),ACSL4在肺部的mRNA表达量极显著高于其他组织(P<0.01),ACSL5在肝的mRNA表达量极显著高于其他组织(P<0.01)。
2.6 水牛ACSLs不同泌乳时期表达分析
在组织表达分析中,ACSL基因家族在水牛泌乳期组织中均有表达,为了进一步探索ACSLs对水牛乳房发育及泌乳性能的作用,该试验选用4个泌乳时期:泌乳第7天(7 d)、第50天(50 d)、第140天(140 d)和第280天(280 d),每个时期2头,活体采集共8头摩拉水牛乳腺组织进行转录组测序,分析了水牛ACSLs不同泌乳时期的表达量差异。TPM计算结果显示,这5个ACSL基因在4个泌乳时期的乳腺组织中均有表达,其中ACSL1在整个泌乳期均有高表达量,尤其是在泌乳第50、140和280天。ACSL3和ACSL5在泌乳第7天和第50天高表达,但随着泌乳天数增加,表达量降低。ACSL4和ACSL6在泌乳第7天表达量最高,但在整个泌乳期的表达量均远低于该家族的其他3个成员(表5、圖6)。
3 讨论
该研究利用生物信息学技术在水牛全基因组序列中鉴定出5个ACSL家族成员,根据它们的进化关系分成了3组。这5个ACSL蛋白长度在699~722 aa,所有蛋白的等电点除ACSL6外均大于7。说明除了ACSL6外,其余家族成员均为碱性蛋白质。Motif分析发现,除ACSL3和ACSL4外缺少motif8外,其余均含有预测出的Motif 1~10且排序相同,说明水牛ACSL基因家族成员蛋白功能相似。
在遗传进化过程中,串联复制和片段复制有助于加速基因家族的扩增和基因组进化机制,为获取新的基因功能提供了可能[ 17],而在进化过程中,亲缘关系越近,共线性越好。该次对水牛和奶牛的种内共线性分析中,均未发现串联重复和片段重复,但是水牛和奶牛种间共线性关系很高并且发现了4对片段重复ACSL基因,说明两者ACSL基因家族的亲缘关系接近[ 18]。
不同物种ACSL系统发育分析结果显示,不同物种的直系同源ACSL基因聚类在一起,可以看出ACSL基因家族具有较高的保守性,其中水牛与奶牛和牦牛的聚类更为接近。在进化中的这种高度保守,也暗示了其在哺乳动物中具有重要的生物学功能。
ACSL家族包括ACSL1、ACSL3、ACSL4、ACSL5和ACSL6 5种不同的异构体,这些蛋白质具有不同的亚细胞定位、脂肪酸底物和组织特异性。长链脂肪酸在脑内的代谢及其与信号分子(如甘油二酰基和LPA)和细胞膜结构成分(包括髓鞘)的结合,需要ACSLs的激活,其中ACSL3和ACSL6是大脑中主要的ACSL亚型[ 19]。除了大脑,BU等[ 20]研究发现,ACSL3是介导肝脏脂肪合成的转录调控的新角色。DONG等[ 21]研究发现,高果糖饮食的仓鼠肝脏中ACSL3 的mRNA和蛋白表达明显减少。ACSL4被发现与人的肺部功能相关,铁凋亡和ACSL4的抑制可通过减少脂质过氧化和增加谷胱甘肽和GPX4水平来减轻肺缺血再灌注(IR)诱导的肺损伤中的铁质损伤[ 22]。Lewin等[ 23]研究发现,大鼠禁食48 h后,肝脏线粒体ACSL5活性增加。尼里水牛不同组织mRNA表达量结果显示,ACSL3和ACSL6在大脑的mRNA表达量极显著高于其他组织(P<0.01)(除了ACSL3大脑低于乳腺组织),ACSL4在肺部的mRNA表达量极显著高于其他组织(P<0.01),ACSL5在肝脏的mRNA表达量显著高于其他组织(P<0.01)。表明在尼里水牛中,ACSL3和ACSL6可能在调节大脑,ACSL4调节肺部,ACSL5调节肝脏的脂肪酸代谢方面发挥重要作用。而肝脏是脂质和脂肪酸代谢的重要部位,游离脂肪酸在肝脏中主要有3种代谢途径,分别是完全氧化合成H2O和CO2;不完全氧化生成酮体及再酯化合成甘油三酯。因此,进入肝脏的游离脂肪酸超过其氧化代谢速率时,代谢发生异常,游离脂肪酸部分再酯化成甘油三酯,从而诱发脂肪积累,使肝脏发生病变[ 24]。因此,研究ACSL3和ACSL5基因对水牛的脂质和脂肪酸代谢具有重要意义。然而ACSLs在摩拉水牛乳腺组织不同泌乳时期的表达量差异与使用尼里水牛组织进行的表达量检测结果有所不同。在摩拉水牛中,ACSL5在泌乳第7天和第50天高表达,表达量随着泌乳天数增加而降低,但ACSL4和ACSL6在整个泌乳期的表达量均远低于该家族的其他3个成员。这表明在不同水牛品种中,即使是相同的ACSL基因家族成员,对其泌乳性能也可能存在着不同的作用效果。
乳脂是一種高品质的脂肪,由甘油三酯(TAG)、二酰甘油、磷脂和丁酸等组成,其中甘油三酯的含量能达到总乳脂的98%。甘油三酯在细胞内会聚集成大脂滴,以乳脂球的形式分泌到牛奶中,因此通常以甘油三酯的含量来研究牛奶中的乳脂含量[ 25]。脂肪酸在乳腺中是被用来合成乳脂的底物,短链脂肪酸(C4-C8)和中链脂肪酸(C8-C14)几乎完全是在乳腺上皮细胞中从头合成的,而长链脂肪酸(>C16)是从血液中直接摄取的,Cl6脂肪酸在2种来源中各占50%左右。大部分长链脂肪酸(LCFA)主要通过ACSLl活化成长链酰基辅酶A穿过细胞膜,然后长链酰基辅酶A将LCFA带入细胞后活化再利用[ 26]。ACSL3能促进卵磷脂的合成和细胞内脂滴的形成,有助于维持脂质平衡[ 27]。ACSL6通过将CoA与脂肪酸连接来激活脂肪酸,对具有C16-C20骨架的饱和脂肪酸以及多不饱和脂肪酸具有相同作用,ACSL6还能参与小脂质滴的生物发生过程及TAG和胆固醇酯的形成过程[ 28-29],这些对乳脂的形成至关重要。此外,LIANG等[ 30]在荷斯坦奶牛中发现了6个ACSL1基因的SNPs,它们在一定程度上与产奶量、乳脂含量、乳蛋白含量和体细胞评分(SCS)有关。Li等[ 31]研究发现,ACSL3是奶牛乳脂肪酸性状相关的20个新的候选基因中的关键基因之一。吕艳涛[ 32]研究发现,泌乳期母猪乳腺中ACSL3在家族成员中基因表达量最高,且随着泌乳的进行mRNA表达量显著升高。王梦岩等[ 33]研究发现,ACSL5基因可能与中国荷斯坦奶牛的乳蛋白代谢相关,这些研究均表明ACSLs与哺乳动物乳脂合成密切相关。而深入挖掘ACSL基因家族的功能也将有助于改善水牛的产奶性状,优化水牛乳制品的品质,对今后提高水牛乳业的经济效益具有重要意义。
参考文献
[1]
黄加祥,黄锋,覃广胜,等.中国奶水牛产业发展综述:国家概况及科技创新[J].中国奶牛,2019(11):1-8.
[2] 庞春英,梁莎莎,马小娅,等.河流型水牛TGF-β基因家族鉴定及其胚胎发育早期的表达分析[J].中国畜牧兽医,2020,47(5):1472-1480.
[3] ZICARELLI L.Buffalo milk:Its properties,dairy yield and mozzarella production[J].Vet Res Commun,2004,28(S1):127-135.
[4] 李辉.Stat5a对山羊乳腺上皮细胞凋亡的调控研究[D].杨凌:西北农林科技大学,2014.
[5] 曹阳.绵羊ACSL1基因克隆和表达及其对脂肪含量的影响[D].长春:吉林农业大学,2016.
[6] KORNBERG A,PRICER W E,Jr.Enzymatic synthesis of the coenzyme A derivatives of long chain fatty acids[J].J Biol Chem,1953,204(1):329-343.
[7] 潘志雄,吕佳,卢立志,等.鹅ACSL1基因克隆及其在鹅肥肝形成中作用的初步研究[J].畜牧兽医学报,2010,41(11):1407-1413.
[8] PAUL D S,GREVENGOED T J,PASCUAL F,et al.Deficiency of cardiac AcylCoA synthetase1 induces diastolic dysfunction,but pathologic hypertrophy is reversed by rapamycin[J].Biochim Biophys Acta,2014,1841(6):880-887.
[9] ZHAO Z D,ABBAS RAZA S H,TIAN H S,et al.Effects of overexpression of ACSL1 gene on the synthesis of unsaturated fatty acids in adipocytes of bovine[J/OL].Arch Biochem Biophys,2020,695[2022-03-17].https://doi.org/10.106/j.abb.2020.108648.
[10] LV Y,CAO Y,GAO Y,et al.Effect of ACSL3 expression levels on preadipocyte differentiation in Chinese red steppe cattle[J].DNA Cell Biol,2019,38(9):945-954.
[11] XU X,GOPALACHARYULU P,SEPPNENLAAKSO T,et al.Insulin signaling regulates fatty acid catabolism at the level of CoA activation[J].PLoS Genet,2012,8(1):1-14.
[12] DUARTE A,PODEROSO C,COOKE M,et al.Mitochondrial fusion is essential for steroid biosynthesis[J].PLoS One,2012,7(9):1-12.
[13] CHEN J N,JIANG Y Z,CEN W M,et al.Distribution of HFABP and ACSL4 gene polymorphisms and their associations with intramuscular fat content and backfat thickness in different pig populations[J].Genet Mol Res,2014,13(3):6759-6772.
[14] MASHEK D G,LI L O,COLEMAN R A.Rat longchain acylCoA synthetase mRNA,protein,and activity vary in tissue distribution and in response to diet[J].J Lipid Res,2006,47(9):2004-2010.
[15] FUJINO T,YAMAMOTO T.Cloning and functional expression of a novel longchain acylCoA synthetase expressed in brain[J].J Biochem,1992,111(2):197-203.
[16] FERNANDEZ R F,KIM S Q,ZHAO Y W,et al.AcylCoA synthetase 6 enriches the neuroprotective omega3 fatty acid DHA in the brain[J].Proc Natl Acad Sci USA,2018,115(49):12525-12530.
[17] LU X R,DUAN A Q,LIANG S S,et al.Genomic identification,evolution,and expression analysis of collagen genes family in water buffalo during lactation[J].Genes,2020,11(5):1-12.
[18] 赵永斌,刘强,刘剑锋,等.杨树和柳树SBP基因的鉴定与进化分析[J].东北林业大学学报,2019,47(12):33-40.
[19] VAN HORN C G,CAVIGLIA J M,LI L O,et al.Characterization of recombinant longchain rat acylCoA synthetase isoforms 3 and 6:Identification of a novel variant of isoform 6[J].Biochemistry,2005,44(5):1635-1642.
[20] BU S Y,MASHEK M T,MASHEK D G.Suppression of long chain acylCoA synthetase 3 decreases hepatic de novo fatty acid synthesis through decreased transcriptional activity[J].J Biol Chem,2009,284(44):30474-30483.
[21] DONG B,KAN C F,SINGH A B,et al.Highfructose diet downregulates longchain acylCoA synthetase 3 expression in liver of hamsters via impairing LXR/RXR signaling pathway[J].J Lipid Res,2013,54(5):1241-1254.
[22] XU Y X,LI X H,CHENG Y,et al.Inhibition of ACSL4 attenuates ferroptotic damage after pulmonary ischemiareperfusion[J].FASEB J,2020,34(12):16262-16275.
[23] LEWIN T M,KIM J H,GRANGER D A,et al.AcylCoA synthetase isoforms 1,4,and 5 are present in different subcellular membranes in rat liver and can be inhibited independently[J].J Biol Chem,2001,276(27):24674-24679.
[24]陈小翠.长时间禁食对大鼠肝脏脂肪酸代谢的影响及其机制研究[D].湘潭:湖南科技大学,2020.
[25] 宋书媛.ACSL6对奶牛乳腺上皮细胞乳脂合成的调控作用研究[D].哈尔滨:东北农业大学,2020.
[26] 张航.不同日粮模式及长链脂肪酸对奶牛乳腺乳脂合成的影响及其机理研究[D].呼和浩特:内蒙古农业大学,2014.
[27] 吕阳,曹阳,张国梁.ACSL3基因研究现状[J].黑龙江畜牧兽医,2018(13):48-51.
[28] FUJIMOTO Y,ITABE H,KINOSHITA T,et al.Involvement of ACSL in local synthesis of neutral lipids in cytoplasmic lipid droplets in human hepatocyte HuH7[J].J Lipid Res,2007,48(6):1280-1292.
[29] GREVENGOED T J,KLETT E L,COLEMAN R A.AcylCoA metabolism and partitioning[J].Annu Rev Nutr,2014,34:1-30.
[30] LIANG Y,GAO Q S,ZHANG Q,et al.Polymorphisms of the ACSL1 gene influence milk production traits and somatic cell score in Chinese Holstein cows[J].Animals (Basel),2020,10(12):1-11.
[31] LI C,SUN D X,ZHANG S L,et al.Genome wide association study identifies 20 novel promising genes associated with milk fatty acid traits in Chinese Holstein[J].PLoS One,2014,9(5):1-21.
[32] 呂艳涛.母猪乳脂合成关键基因筛选及长链脂肪酸对乳脂合成的调控研究[D].广州:华南农业大学,2016.
[33] 王梦岩,姜平,余湘,等.奶牛乳腺组织中ACSL5基因的表达量差异分析[J].中国兽医学报,2019,39(1):100-104.
