一株侵染棕榈象甲幼虫的白僵菌的分离鉴定及其毒力
2023-08-05薛锐陈杰付玉飞等
薛锐 陈杰 付玉飞等

关键词 球孢白僵菌;红棕象甲;昆虫病原真菌;生物学特性;毒力
中图分类号:S 476.12 文献标识码:A DOI:10.16688/j.zwbh.2022417
红棕象甲Rhynchophorus ferrugineus Oliver属鞘翅目Coleoptera象甲科Curculionidae棕榈象属Rhynchophorus,别名锈色棕榈象、椰子隐喙象、椰子甲虫等,于2005年被国家林业局列入林业检疫性有害生物名录[1]。红棕象甲起源于亚洲南部及西太平洋上美拉尼西亚群岛[2],于20世纪80年代通过苗木调运传人地中海沿岸、大洋洲和拉丁美洲等40余个国家和地区,国内在海南、广东、福建、云南等长江以南地区发生为害[3-4]。红棕象甲寄主包括加纳利海枣、椰子、槟榔、大王棕、霸王棕、油棕和美丽针葵等30余种棕榈科植物[5]。红棕象甲成虫在寄主植物叶柄基部的伤痕、裂口处产卵,卵孵化后,幼虫首先蛀人幼嫩的叶柄基部,后蛀人茎干内,形成隧道并在隧道内留下植物纤维和排泄物,其取食幼嫩组织,破坏植株生长点最终导致寄主死亡[6]。因其隐蔽取食特点及生活习性,防控难度大。现主要应用化学手段对其进行防治,但易造成环境污染并可能导致人畜中毒等问题。目前国内采用生物防治手段防治红棕象甲的研究主要集中于金龟子绿僵菌Metarhizium anisopliae、黏质沙雷氏菌Serratia marcescens、淡紫拟青霉Paecilomyces lilacinus及昆蟲病原线虫[7-9]。国内尚未见有自然侵染红棕象甲的白僵菌的相关研究报道。
白僵菌Beauveria是一类常见的昆虫病原真菌,因其寄主范围广、易于培养、对环境友好等特点,具有很强的开发潜力[10-12]。自1956年我国首次报道关于白僵菌防治甘薯象甲Cylas formzcarzus的研究后,对此方向的研究愈发深入,白僵菌现已广泛被应用于防治松墨天牛Monochamus alternatus、西花蓟马Frankliniella occidentalis、亚洲玉米螟Os-trinia furnacalis、草地贪夜蛾Spodoptera frugi-perda等害虫[13-17]。近年来,我国利用昆虫病原真菌防治象甲科类害虫的报道逐年增加。朱晓敏等[18]发现,球孢白僵菌Beauveria bassiana与苦参碱复配后对稻水象甲Lissorhoptrus oryzophilus表现出加成以及增效作用;张强等[19]发现田间施用250亿孢子/g白僵菌可湿性粉剂对稻水象甲的防治效果可达65%以上;陈建军等[20]利用400亿孢子/g白僵菌可湿性粉剂对稻水象甲成虫防治效果理想,施药后7d防治效果高于10%醚菊酯悬浮剂以及200g/L氯虫苯甲酰胺悬浮剂。由于寄主种类不同以及外界环境的多样性,不同白僵菌在形态特征及致病力方面存在差异[21],为白僵菌的分类鉴定带来了难度。目前,将形态学特征和核酸序列分析相结合已成功用于白僵菌[22-23]和绿僵菌[24-26]的分类鉴定。
本研究自昆明市盘龙区云南农业大学采集的罹病红棕象甲幼虫上分离纯化获得1株病原真菌,采用形态学特征及ITS-rDNA序列对该菌株进行鉴定,同时测定了该菌株的生物学特性以及对红棕象甲不同龄期幼虫的致病力,旨在为生物防治红棕象甲提供白僵菌生防药剂的备选菌株。
1材料与方法
1.1菌株培养与分离纯化
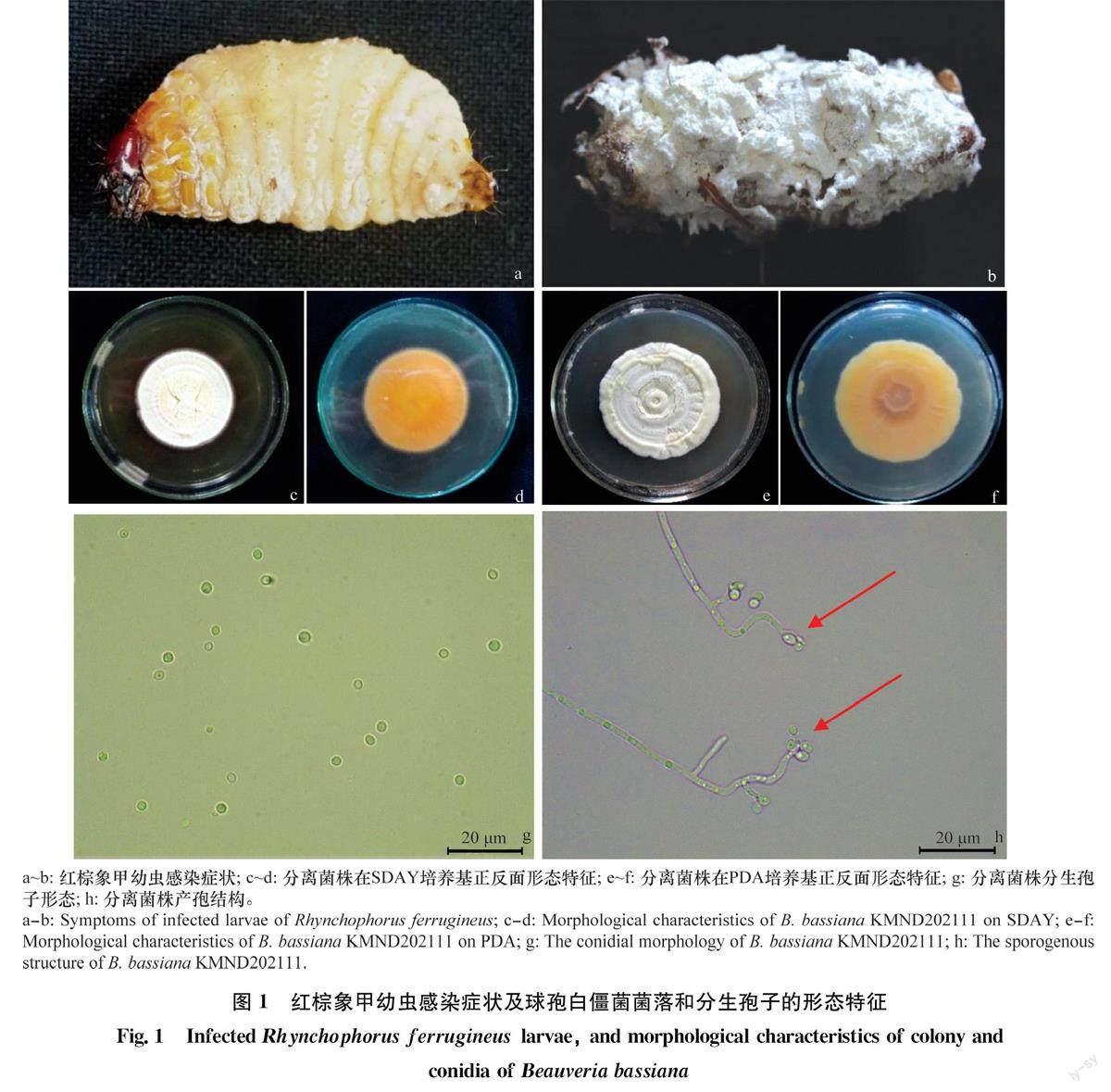
将采自云南省昆明市云南农业大学(25°07′N,102°44′E,海拔1950 m)校园内棕榈树上的罹病红棕象甲幼虫保湿培养,待虫体表面出现菌丝或分生孢子后,在超净工作台内用接种针挑取少量分生孢子,采用点接法接种于马铃薯葡萄糖琼脂(PDA)培养基上[26-27],置于(25±1)℃,相对湿度(75±5)%,L∥D=12h∥12h的恒温培养箱中培养获得菌株。菌株经单孢纯化后命名为KMND202111,转入PDA斜面培养基生长5d后置于40C冰箱内保存备用。
1.2供试培养基
马铃薯葡萄糖琼脂培养基(PDA):去皮马铃薯块200 g煮汁过滤,葡萄糖20 g,琼脂20 g,蒸馏水1000mL[27]。马铃薯蔗糖琼脂培养基(PSA):葡萄糖替换为等量蔗糖,其他同PDA[27]。萨氏葡萄糖琼脂培养基(SDAY):葡萄糖40 g,蛋白胨10 g,酵母浸粉10g,琼脂20 g,蒸馏水1000mL[27]。萨氏麦芽糖酵母培养基( SMAY):葡萄糖替换为等量麦芽糖,其他同SDAY[27]。
1.3菌株鉴定
1.3.1形态鉴定
将菌株分生孢子点接于PDA培养基(直径90 mm)中央,在(25±1)℃,相对湿度(75±5)%,L∥D=12h∥12h的恒温培养箱中倒置培养,每日观察菌落形态及颜色。接种后5d,挑取新鲜菌丝于灭菌载玻片上,在显微镜下观察分生孢子梗形态特征。7d后挑取分生孢子制作玻片每组随机观测20个分生孢子的形状与大小,设置5次重复。
1.3.2分子生物学鉴定
用1mL无菌枪头刮取菌株KMND202111长势良好的菌丝体,置于无菌研钵中,用液氮快速研磨成粉末,转入1.5 mL离心管中,采用改良的CTAB法[28]并稍作修改(本试验将65℃恒温水浴时间由1h替换为30min,将TE缓冲液替换为ddHO,其他步骤不做更改)提取DNA。以菌株基因组DNA为模板,采用真菌通用引物ITS1(5′-TCCGTAG-GTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCT-TATTGATATGC-3′)扩增其rDNA-ITS-ITS列[29]。PCR反应体系为25μL,其中ITS1和ITS4各1μL,PCR MasterMix 12.5μL,模板DNA 1μL,超纯水9.5μL。PCR反应条件:95℃预变性3 min;940C变性1 min,55℃退火1 min,72℃延伸1.5 min,35个循环;72℃延伸10 min。1%琼脂糖凝胶电泳检测PCR产物,在凝胶成像系统下观察扩增结果[30]。扩增产物送生工生物工程(上海)股份有限公司测序。
去除所得序列两端质量不好的碱基,将优化后的序列提交至NCBI网站进行BLAST比对,下载与目标序列具有高同源性的序列,使用MEGA X软件通过邻接法(neighbour-joining method,NJ),运行1000次bootstrap验证,构建菌株的系统发育树[31]。
1.4分离菌株的生物学特性测定
1.4.1培养基对菌株生长和产孢的影响
分别测定菌株在SDAY、SMAY、PDA及PSA培养基上的生长和产孢情况。采用点接法将分生孢子接种于上述培养基平板中央,置于25℃,光周期L∥D=0h∥24h培养箱内(因采集到的罹病红棕象甲幼虫处于预蛹阶段,因此将初始光周期设定为全黑暗),定时观察并记录菌落色泽、形态及产孢时间等。培养至第12天,采用十字交叉法测量菌落直径,并计算菌落生长速率。用灭菌打孔器(直径10 mm)自菌落中心点至边缘距离的1/2处打取菌饼,每处理打取3个菌饼,置于含有20 mL无菌的0.05% Tween-80溶液的三角瓶中,磁力搅拌10min获得孢子悬浮液,用血球计数板测定孢子浓度,并计算菌落单位面积的产孢量[32]。每处理重复5次。
1.4.2光周期对菌株生长和产孢的影响
将菌株分生孢子接种于1.4.1选出的最佳培养基上,置于25℃,光周期分别为L∥D=24h∥0h、L∥D=12h∥12h、L∥D=0h∥24h条件下培养12d,测定菌落直径,计算生长速率及产孢量。每处理重复5次。
1.4.3温度对菌株生长和产孢的影响
采用1.4.1和1.4.2筛选出的适宜培养基及光周期条件,温度分别设置为20、25、30℃,培养12 d后测定菌落直径。按1.4.1方法测定菌落生长速率及产孢量。每处理重复5次。
1.5分离菌株的致病力测定
采用浸渍法[33]测定菌株致病力。将红棕象甲3龄及6龄幼虫在浓度为108个/mL的孢子悬浮液中浸渍15s后挑出,用灭菌滤纸吸干虫体表面多余水分后移至皿底垫有湿润滤纸片的培养皿(直径20 cm)内单头饲养,每日更换新鲜甘蔗供其取食,以0.05% Tween-80浸渍为对照。将处理后的幼虫置于温度(25±1)℃,相对湿度(75±5)%,光周期L∥D=0h∥24h的人工气候箱中饲养。每处理设置3个重复,每重复24头幼虫,接种后每天定时观察,连续观察14d,并记录幼虫死亡数量,计算累计死亡率及校正死亡率。对死亡后的幼虫进行单头保湿培养观察,待死亡虫体表面长出白色菌丝或白色分生孢子时则视为被白僵菌侵染致死。
1.6数据分析
用统计软件SPSS 22.0对菌落直径及单位面积产孢量间的差异显著性进行LSD(least significantdifference)检验,采用Probit方法计算致死中时(LT)。利用Origin 2020绘图。
菌落直径(mm)=(菌落横径+菌落纵径)/2;
菌落生长速率(mm/d)=平均菌落直径/培养天数;
菌落单位面积产孢量(个/mm)=平均每小格孢子数×400×104×稀释倍数/菌饼面积;
累计死亡率=(处理组累计死亡总虫数/处理总虫数)×100%;
校正累计死亡率=(处理组累计死亡率-对照组累计死亡率)/(1-对照死亡率)×100%。
2结果与分析
2.1菌株KMND202111的形態特征及分子生物学鉴定
菌株KMND202111在培养基上生长情况如图1c~f。培养第3天,菌落平展,边缘整齐呈白色绒毛状,呈扩环状生长;第5天开始产孢,菌落变为淡黄色,中间略凸起,孢子层呈粉状,长势均匀,菌落背面淡黄色并出现褶皱。在光学显微镜下,分生孢子无色、光滑、圆球形,少数卵圆形,直径2. 1~2.4μm。营养菌丝细长无色;分生孢子梗在顶部轴式产孢,形成穗状产孢结构。
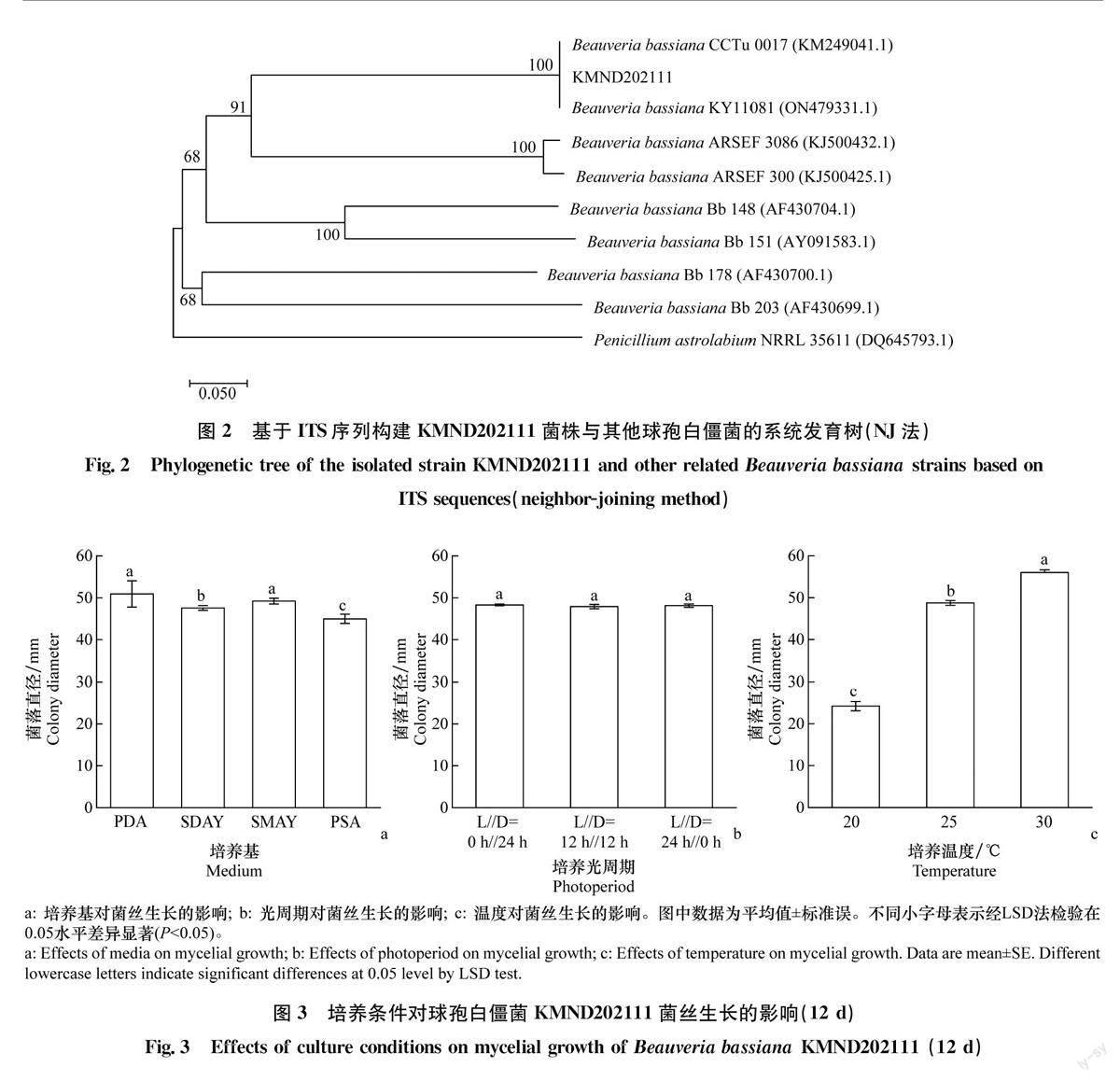
使用PCR扩增分离菌株rDNA-ITS序列片段,测序结果显示扩增片段为543bp,去除两端低质量序列后在GenBank数据库中进行BLAST比对,发现目的菌株ITS序列与已知球孢白僵菌对应序列的最高相似性可达到99.44%,选取相关序列,使用MEGA 11.0构建系统发育树(图2),其中KMND202111菌株与Beauveria bassiana CCTu0017(KM249041.1)和Beauveria bassiana KY11081(ON479331.1)处于进化树最小分支,亲缘关系最近,说明该菌株与NCBI数据库中球孢白僵菌具有高度相似性。综合形态特征鉴定和ITS序列相似性分析,确定该菌株为球孢白僵菌。
2.2不同培养条件对菌株KMND202111生长的影响
由图3a可知,球孢白僵菌菌株KMND202111在供试培养基上均可生长,但生长至第12天时菌落直径及生长速率均存在显著差异(菌落直径:F=6.493,P<0.05;生长速率:F=4.312,P<0.05)。菌株在PDA及SMAY培养基上生长最快,第12天时菌落直径分别为50.94mm和49.23 mm;菌落生长速率分别为4.24mm/d和4.10mm/d,显著高于在PSA(3.75mm/d)和SDAY(3.97mm/d)培养基上的生长速率。
球孢白僵菌菌株KMND202111在24h连续黑暗、12h光暗交替、24h连续光照条件下均可生长(图3b),生长至第12天各处理间菌落直径及生长速率未见显著差异(菌落直径:F=1.053,P>0.05;生长速率:F=1.144,P>0.05)。
3个温度下球孢白僵菌菌株KMND202111生长12 d的菌落直径以及生长速率存在显著差异(菌落直径:F=1701. 135,P<0. 05;生长速率:F=799.51,P<0.05)。在20、25℃和30℃条件下,菌落生长速率依次为2.02、4.06、4.67mm/d,其中30℃条件下生长最快,第12天菌落直径为56.00mm,显著高于其余处理(图3c)。
2.3不同培养条件对菌株株KMND202111产孢量的影响
球孢白僵菌KMND202111在不同培养基上单位面积产孢量存在极显著差异(F=64.463,P<0.01)。其中在PDA培养基上的产孢量最高,达到6.38×10个/mm,极显著高于其他处理;在SDAY培养基上产孢量为3.20×10个/mm,极显著低于其余处理(图4a)。综上所述,球孢白僵菌KMND202111菌株适宜在PDA培养基上生长。
球孢白僵菌KMND202111于24 h连续黑暗、12 h光暗交替、24 h连续光照条件下培养12 d,产孢量没有显著差异(F=0.383,P>0.05),由此表明,该菌株对光周期的变化不敏感(图4b)。
3个培养温度下球孢白僵菌KMND202111的产孢量存在极显著差异(F=428.20,P<0.01),在30℃下培养至第12天菌落单位面积产孢量为9.09×10个/mm,极显著高于其他温度处理;在20 0C下,产孢量为2.47×10个/mm,极显著低于其余处理(图4c)。
2.4菌株KMND202111对红棕象甲幼虫的毒力
毒力测定结果表明,球孢白僵菌KMND202111对红棕象甲具有侵染作用(图1a~b)。浓度10个/mL的孢子悬浮液接种处理后第3天,3龄幼虫累计死亡率明显大于对照;接种处理后第4天,6龄幼虫累计死亡率明显大于对照。将死亡虫体置于光照培养箱中保湿培养,第3天时死亡虫体的首末端开始有菌丝长出,第6天时幼虫虫体长满菌丝。第10天,球孢白僵菌KMND202111菌株对红棕象甲3龄幼虫的累计校正死亡率达到100%(图5a),毒力回归方程为y=4.12x-2.67(R=0.917),LT为4.88d,95%置信区间为3.611~5.793 d;第10天,球孢白僵菌KMND202111對红棕象甲6龄幼虫的累计校正死亡率为76.4%,第14天达到90.3%(图5b),毒力回归方程为y=3.07x=2.55(R=0.902),LT为7.64 d,95%置信区间为5.903~8.446d。
3结论与讨论
红棕象甲是一种重要的园林植物害虫,国内外许多学者十分关注昆虫病原体例如球孢白僵菌[34]、金龟子绿僵菌[35]、病原线虫[36]以及苏云金芽胞杆菌[37]等对其毒力作用及防控应用。国内对于该虫防治主要集中于金龟子绿僵菌[26]、淡紫拟青霉[3]及黏质沙雷氏菌[3]的利用,尚未见侵染红棕象甲的白僵菌相关研究报道。本研究从自然罹病红棕象甲幼虫上分离获得1株病原真菌KMND202111,根据该菌株形态特征以及ITS序列比对分析,鉴定该菌株为球孢白僵菌。
球孢白僵菌是目前广泛应用的杀虫真菌,其不同菌株的生物学特性及致病力随其寄主的不同而存在差异。黄鹏等[38]发现,分离自南洋臀纹粉蚧Pl-anococcus lilacinus和石蒜绵粉蚧Phenacoccus sola-ni的球孢白僵菌BB-T02在28℃,光周期L∥D=8h∥16h条件下生长发育良好。温绍海等[22]发现,分离自蠼螋成虫的球孢白僵菌HNC-1及YY-1在25℃下生长速率、产孢量以及萌发率最高;陈春艳等[39]发现,分离自栗实象Curculio davidi的球孢白僵菌在25~26℃,光周期为L∥D=14 h∥10 h条件下生长速率以及产孢量最佳;张磊等[40]研究发现,分离自大豆高隆象Ergania doriae yunnanus的球孢白僵菌BEdyl菌株在26℃下生长速率最高,在22℃下产孢量最大;王定锋等[41]研究发现,分离自茶丽纹象甲Myllocerinus aurolineatus的布氏白僵菌Beauveria brongniartii Bbr1552在蛋白胨马铃薯葡萄糖琼脂培养基(PPDA)上25℃时生长发育及产孢量最高。本研究发现,分离自红棕象甲幼虫的球孢白僵菌KMND202111菌株适宜生长的温度为30℃,而光周期对其生长发育无明显影响。由此表明,分离自红棕象甲幼虫的球孢白僵菌KMND202111具有较强的耐热能力,适合在30℃左右环境中使用。
本研究发现菌株KMND202111对红棕象甲3龄及6龄幼虫有较强毒力,其LT分别为4.88 d和7.64 d。相较于国内外其他用于防治红棕象甲幼虫的生防真菌,该菌株毒力更强,致死中时更短[42-45]。球孢白僵菌对寄主昆虫的侵染是附着胞产生的机械压力以及分泌酶共同作用的结果[46],分生孢子附着于虫体壁后诱导分泌胞外蛋白酶,使其可以牢固附着于寄主昆虫虫体表面[47],并通过蛋白质酶、几丁质酶、脂酶以及淀粉酶等进一步降解昆虫表皮完成侵染。不同昆虫病原真菌的致病力与某些酶(如蛋白质酶和几丁质酶)的含量及活性有关[48-49]。据报道,黄粉虫Tenebrio molitor、褐飞虱Nilaparvata lugens、马铃薯甲虫Leptinotarsa decemlineata不同发育阶段对虫生真菌敏感性存在差异[50-52],李浩等[53]和Yu等[54-55]研究认为,老熟幼虫体内的解毒酶更容易被诱导,且活性较高,随着昆虫逐渐老熟,其表皮结构也更为完善,因而对生防真菌的敏感性降低。本研究分离的菌株对棕榈象甲幼虫表现出较强的致病力,但其侵染致病作用是否与蛋白质酶和几丁质酶含量与活性相关还需要深入探究。
