赤霉素和磷肥配合施用对胡麻光合酶活性及其产量形成的影响
2023-08-04王瑛泽高玉红王一帆崔政军
王瑛泽,曹 智,高玉红,,吴 兵,王一帆,剡 斌,,崔政军,徐 鹏
(1.甘肃农业大学农学院,甘肃 兰州 730070;2.甘肃省干旱生境作物学国家重点实验室,甘肃 兰州 730070;3.甘肃农业大学生命科学技术学院,甘肃 兰州 730070)
光合作用是决定作物生长发育和产量形成的重要因素,也是植物生命活动所需物质和能量的主要来源。脱落酸、生长素和赤霉素等激素参与调控植物光合器官的发育与功能维持[1],其中赤霉素(Gibberellic acid,GA)作为调控信号参与植物生长发育的全过程[2]。施用外源GA3可以提高作物光合酶和氮代谢酶活性,增加作物株高、叶面积和生长速率,促进光合同化物的积累[3],提高小麦千粒重[4]和马铃薯单薯重[5],从而获得高产。磷作为作物生长的必需元素之一,是植物体内核酸、磷脂等生物大分子的结构组分,同时参与光合磷酸化过程和碳水化合物的合成与运转过程,在作物生长发育和产量形成方面发挥着重要作用[6]。缺磷不仅导致植株叶片的叶绿素含量[7]、PEP羧化酶活性、RuBP羧化酶活性降低[8],还会抑制叶片扩展,造成叶面积减小、光截获量减少,从而降低光合同化物的积累[9];而磷肥过量时,作物呼吸作用增强,容易徒长,造成养分过量消耗,严重时影响作物生长发育,导致其产量下降[10]。因此,合理的磷肥运筹是改善作物光合特性、提高作物产量的重要调控措施。有研究发现,叶面喷施GA3与有机、无机肥配施可促进辣椒生长发育,增加其干物质积累[11];赤霉素与钾肥配合施用可以提高烟叶碳氮代谢酶活性,增强烟株的碳氮代谢强度,增加植株中钾的积累[12];在氮、磷肥处理试验中,GA3的喷施在作物养分吸收方面发挥了重要作用,其不仅提高了养分利用率,促进作物高产,同时最大限度地减少了农田氮和磷的输入和损失[13]。可见,外源GA3与肥料互作可以改善作物光合特性,促进其生长发育和产量形成。
胡麻(LinumusitatissimumL.)是我国重要的油料作物和经济作物,多种植于温带地区[14]。胡麻油因富含α-亚麻酸、木酚素等营养成分和对预防高脂血症、结肠肿瘤和动脉硬化等多种疾病有积极作用而受医用、食品、农业等诸多领域的关注[14]。研究表明,增施磷肥可以增强胡麻叶片的光合作用,协调植株各器官内养分吸收、同化与转运三者之间的关系,促进更多的光合产物向籽粒转运,从而提高籽粒产量[15]。然而,现阶段施入农田的磷只有10%~25%被植物吸收,其余均被释放至环境,这不仅增加了农业生产成本,浪费了有限的磷资源,还不利于保护环境和维持生态平衡[16]。目前,外源激素已经被普遍应用于提高肥料利用率、调节作物生长发育、增加产量等方面[13, 17],关于赤霉素与有机肥、钾肥互作的研究已有报道[11-12],而其与磷肥配合施用对胡麻等作物光合酶活性及其产量的影响机理尚不明确。本文通过研究外源GA3在不同磷肥水平下对胡麻光合酶活性及其产量的调控效应,以期为胡麻高产栽培提供理论依据与技术支撑。
1 材料与方法
1.1 试验区概况
试验于2019年3月—2020年8月在甘肃省定西市农业科学院油料所试验站(34°26′ N,103°52′ E)进行,试验区位于陇中黄土高原半干旱丘陵沟壑区,属于中温带干旱半干旱区,平均海拔2 050 m,年均气温6.43℃,≥10℃积温2 239.1℃,多年平均降水390.9 mm,年均日照时数2 476.6 h,无霜期约140 d。试验地为梯田,土质为黄绵土,肥力均等,0~30 cm土层土壤基础养分如下:有机质17.51 g·kg-1、全氮0.81 g·kg-1、全磷0.69 g·kg-1、速效磷27.43 mg·kg-1、速效钾108.30 mg·kg-1,pH 8.14。
1.2 试验设计
试验采用田间二因素裂区设计,以磷肥(纯P2O5)施用量为主处理,设3个水平[18]:P0:0 kg·hm-2,P1:67.5 kg·hm-2,P2:135 kg·hm-2;以GA3喷施浓度为副处理,设3个水平:G0:0 mg·L-1,G1:15 mg·L-1,G2:30 mg·L-1。共9个处理,3次重复,27个小区。主区面积18 m2(6 m×3 m),副区面积6 m2(3 m×2 m),小区和重复间均间隔80 cm,四周设1 m保护行。
供试材料为‘轮选2号’胡麻品种,由内蒙古自治区农牧业科学院提供。播种量750万粒·hm-2,播深3 cm,行距20 cm,保苗375万株·hm-2。除依据不同水平施用磷肥外,各处理均施用氮肥150 kg N·hm-2、钾肥52.5 kg K2O·hm-2,供试氮肥为尿素(N 46%)、磷肥为过磷酸钙(P2O516%)、钾肥为硫酸钾(K2O 52%),所有肥料全部做基肥施入。赤霉素分别在胡麻现蕾期、盛花期喷施,各处理每次喷施1 L,G0喷施等量清水。胡麻于2019年4月10日播种,8月31日收获;2020年4月9日播种,8月19日收获。其他管理方式同当地大田。
1.3 测定项目与方法
植株干物质积累量测定:2019年和2020年于胡麻苗期、现蕾期(喷GA3前)、盛花期(喷GA3前)、青果期和成熟期分别在各小区采集具有代表性且长势基本一致的植株样品30株,采用烘干法[19]测定植株地上部干物质积累量。
酶活性测定:2019年和2020年胡麻现蕾期和盛花期喷施GA3前、喷施GA3后1、3、5、10 d分别在各小区采集具有代表性且长势基本一致的植株样品30株,取中上部叶片放入液氮中,带回实验室储藏于-80℃冰箱,统一采用苏州科铭技术有限公司生产的试剂盒提取叶片中核酮糖二磷酸羧化酶(RuBP羧化酶)、磷酸烯醇式丙酮糖羧化酶(PEP羧化酶),用酶标仪测定其活性。
籽粒产量测定:2019年和2020年胡麻成熟收获时,按小区单打单收,晾晒至胡麻籽粒水分含量低于10%后称取籽粒重量,测得各处理实际籽粒产量。
1.4 数据处理
采用Excel 2019进行数据整理和作图,使用SPSS 21.0统计分析软件进行相关性分析以及主效应检验,运用LSD法进行显著性检验。
2 结果与分析
2.1 磷肥和外源GA3对胡麻叶片RuBP羧化酶活性的影响
2.1.1 对胡麻现蕾期叶片RuBP羧化酶活性的影响 增施磷肥和喷施外源GA3均可显著增加胡麻现蕾期叶片RuBP羧化酶活性(P<0.05,表1、图1)。由表1可知,不同施磷量之间,喷施GA3前,胡麻叶片RuBP羧化酶活性随施磷量的增加而逐渐提高,其中,P2水平较P0和P1水平分别显著提高44.20%和9.82%;喷施GA3后,不同喷后天数均表现为P1、P2>P0,喷后1、3 d和10 d,P1分别较P0显著提高121.78%、18.84%和17.78%,P2分别较P0显著提高80.86%、12.09%和15.31%。各GA3浓度之间,不同喷后天数叶片RuBP羧化酶活性变化规律有所不同,喷后1、3 d和5 d均表现为G1>G2、G0,G1分别较G0显著增加27.01%、11.76%和20.91%,喷后10 d,G1与G2之间差异不大,但均显著高于G0。如图1所示,施磷水平与外源GA3交互作用均对喷后1~10 d胡麻叶片RuBP羧化酶活性有极显著影响(P<0.01),其中,P1G2处理在喷后1、3 d和10 d叶片RuBP羧化酶活性均达最高,P2G1处理则在喷后5 d最高,二者较P0G0处理显著提高29.95%~258.56%。可见,施磷67.5 kg·hm-2配合喷施GA315 mg·L-1和施磷135 kg·hm-2配合喷施GA330 mg·L-1均可显著提高胡麻现蕾期叶片RuBP羧化酶活性。
2.1.2 对胡麻盛花期叶片RuBP羧化酶活性的影响 增施磷肥和喷施外源GA3均可显著增加胡麻盛花期叶片RuBP羧化酶活性(表2、图2)。由表2可知,不同施磷量之间,喷施GA3前,胡麻盛花期叶片RuBP羧化酶活性随着施磷量的增加显著增加;喷后1 d,P1和P2分别较P0显著提高76.30%和68.54%;喷后3 d和10 d,变化规律均表现为P1>P2>P0,P1较P0分别显著提高236.48%和184.46%,较P2分别显著提高154.82%和55.22%。除喷后3 d外,P0和P1水平下喷施GA3后胡麻叶片RuBP羧化酶活性均显著提高。不同GA3浓度之间,喷后1 d和5 d均表现为G1>G2>G0,G1分别较G0显著提高46.65%和15.31%;喷后3 d和10 d则为G2最高,分别较G0显著提高45.91%和46.47%。

表1 2020年不同磷肥和GA3水平对胡麻现蕾期叶片RuBP羧化酶活性的影响/(nmol·min-1·g-1)
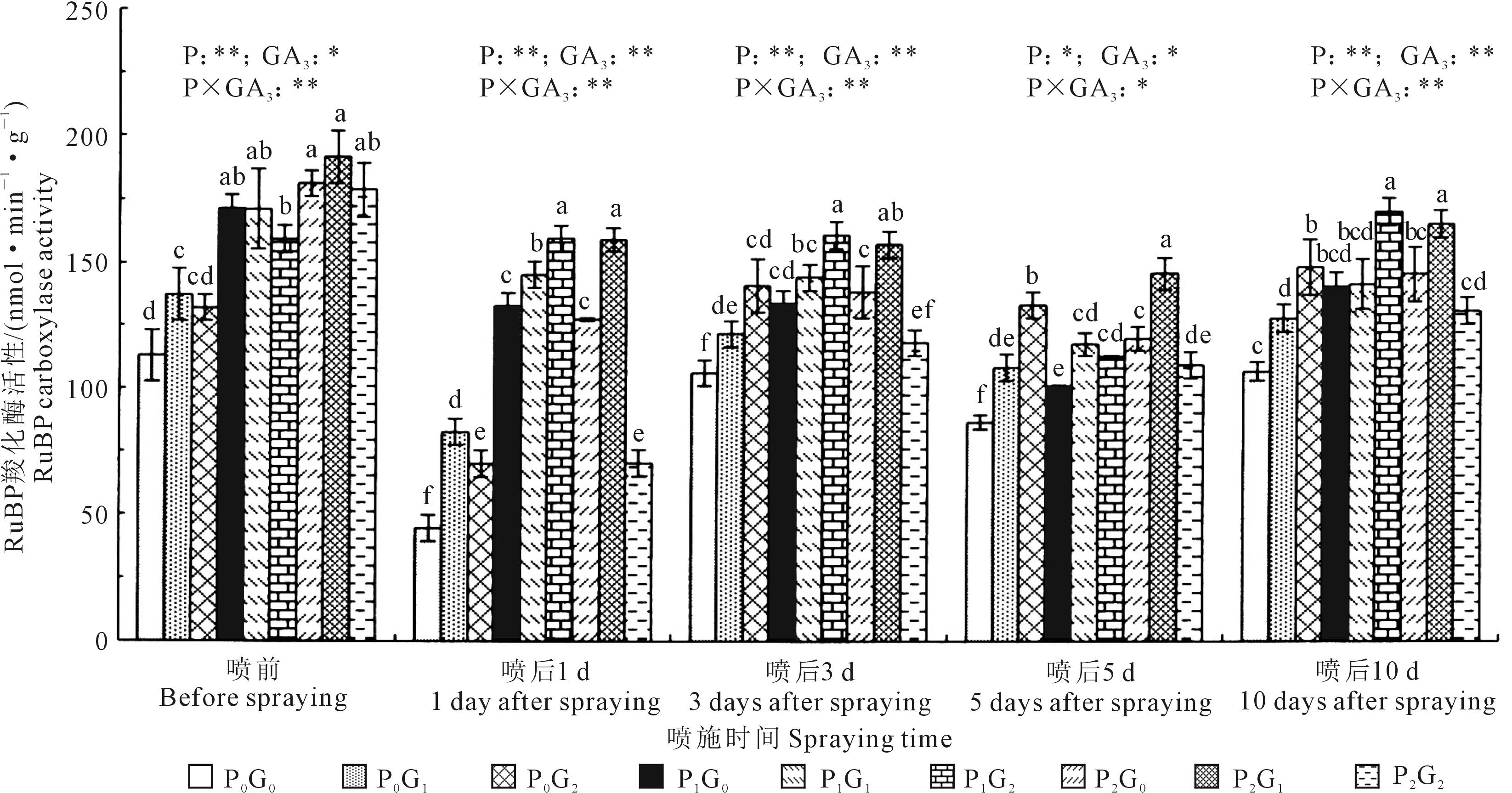
注:不同小写字母表示同一喷施时间下处理之间差异显著(P<0.05);*表示在P<0.05水平下差异显著,**表示在P<0.01水平下差异显著,“-”表示尚未喷施GA3。下同。

表2 2020年不同磷肥和GA3水平对胡麻盛花期叶片RuBP羧化酶活性的影响/(nmol·min-1·g-1)

图2 2020年不同施磷量和GA3处理下盛花期胡麻叶片RuBP羧化酶活性
由图2可以看出,不同施磷量下,RuBP羧化酶活性因GA3喷施浓度不同差异显著。P0和P1水平下,叶片RuBP羧化酶活性均以G2处理最高;P2水平下则以G1处理最高,较G0显著提高45.98%~73.80%。在不同喷施时间,磷肥与GA3之间均存在显著的交互效应,喷后1 d和喷后5 d均表现为P2G1处理最高,分别较P0G0处理显著提高229.14%和100.00%;喷后3 d和10 d则以P1G2处理表现最优,分别较P0G0显著提高11.88倍和4.29倍。施磷量、GA3喷施浓度及二者互作均显著影响胡麻盛花期叶片RuBP羧化酶活性。结果表明,喷施15~30 mg·L-1GA3配施67.5~135 kg·hm-2磷肥可显著提高胡麻盛花期叶片RuBP羧化酶活性。
2.2 磷肥和外源GA3对胡麻叶片PEP羧化酶活性的影响
2.2.1 对胡麻现蕾期叶片PEP羧化酶活性的影响 如表3所示,除2020年GA3喷后10 d外,增施磷肥在其余年份和喷施时间均显著提高了胡麻现蕾期叶片PEP羧化酶活性。不同施磷量之间,2个生长季节内,喷前、喷后1 d,P1和P2叶片PEP羧化酶活性均显著高于P0,增幅分别为126.72%~161.50%(喷前)和14.46%~23.90%(喷后1 d);喷后3 d和5 d,P2显著高于其他处理,较P0和P1分别显著提高8.68%~61.86%(3 d)和25.44%~174.85%(5 d)。不同GA3喷施浓度之间,2个生长季胡麻叶片PEP羧化酶活性在喷后1 d均表现为G1>G2>G0,各水平间差异显著;喷后3、5 d和10 d,基本表现为G2最高,分别较G0显著提高44.03%~52.63%、39.20%~86.10%和15.11%~28.54%。表明GA3对PEP羧化酶的促进作用随时间的推移呈先升高后降低的趋势,在喷后5 d时达最大。
由图3可知,施磷量对GA3喷前及喷后1~10 d胡麻现蕾期叶片PEP羧化酶活性影响显著(2020年喷后10 d除外),GA3浓度极显著影响喷后1~10 d叶片PEP羧化酶活性,二者对喷后1~10 d叶片PEP羧化酶活性的互作效应均达极显著水平(2020年喷后10 d除外)。其中,P2G1处理叶片PEP羧化酶活性在喷后1、3 d和5 d均较高,分别较P0G0处理显著提高183.74%~214.73%、92.6%~106.92%和290.60%~359.65%。结果表明,施磷135 kg·hm-2配合现蕾期喷施GA315 mg·L-1可显著提高胡麻现蕾期叶片PEP羧化酶活性,有利于光合同化物的转化。

表3 不同磷肥和GA3水平对胡麻现蕾期叶片PEP羧化酶活性的影响/(nmol·min-1·g-1)
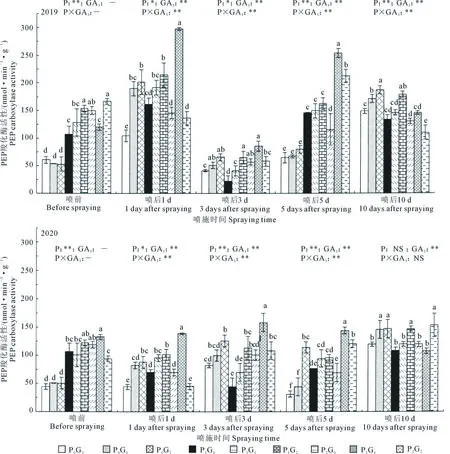
图3 不同施磷量和GA3处理下现蕾期胡麻叶片PEP羧化酶活性
2.2.2 对胡麻盛花期叶片PEP羧化酶活性的影响 施磷量和GA3喷施浓度均对胡麻盛花期叶片PEP羧化酶活性影响显著(表4、图4)。由表4可知,2个生长季节内,不同施磷量之间,喷施GA3前后胡麻盛花期叶片PEP羧化酶均表现为P1>P2>P0,P1较P0分别显著提高82.03%~87.23%(喷前)、72.07%~106.46%(喷后1 d)、113.68%~126.28%(喷后3 d)、132.82%~166.64%(喷后5 d)和43.13%~85.43%(喷后10 d)。

表4 不同磷肥和GA3水平对胡麻盛花期叶片PEP羧化酶活性的影响/(nmol·min-1·g-1)

图4 不同施磷量和GA3处理下盛花期胡麻叶片PEP羧化酶活性
不同GA3喷施浓度之间,喷后1 d,2个生长季内G1和G2分别较不喷施GA3(G0)平均显著提高26.78%和33.90%;喷后3、5 d和10 d,PEP羧化酶活性均为G1最高,分别较G0显著提高13.41%~43.80%、17.42%~45.91%和38.57%~47.58%。
如图4所示,GA3喷施浓度对2个生长季不同喷施时期叶片PEP羧化酶活性均有极显著影响,不同施磷量下PEP羧化酶活性随GA3喷施浓度变化规律有所不同,除2020年喷前、喷后3 d、喷后5 d外,P1水平下胡麻叶片PEP羧化酶活性随GA3喷施浓度的增加而提高;P2水平下则均为G1最高。同一GA3浓度下,G1水平PEP羧化酶活性表现为P2>P1>P0(2019年喷后5 d和2020年喷后3 d除外);G2水平则为P1最高。互作效应分析表明,P×GA3对2个生长季节内叶片PEP羧化酶活性的影响均达极显著水平,其中P1G2和P2G1处理下胡麻叶片PEP羧化酶活性较P0G0显著提高147.44%~265.87%。综上可知,增施磷肥后喷施GA3能够发挥二者的协同作用,提高胡麻叶片PEP羧化酶活性,可为进一步提高胡麻籽粒产量奠定基础。
2.3 磷肥和外源GA3对胡麻干物质积累量的影响
由图5可知,2个生长季节内,随着胡麻生育进程的推进,全生育期干物质积累量先增加后降低,在青果期达到最大。不同施磷量之间,苗期和现蕾期的干物质积累量均表现为P2>P1>P0,苗期P2和P1分别较P0显著提高16.07%~28.35%和14.29%~23.04%,现蕾期分别显著增加16.86%~73.25%和12.96%~41.60%。不同GA3喷施浓度之间,盛花期~成熟期的干物质积累量均表现为G2、G1显著高于G0,G2、G1之间差异不大(表5)。

图5 不同施磷量和GA3处理下胡麻干物质积累量

表5 不同磷肥和GA3水平对胡麻干物质积累量的影响/(kg·hm-2)
如图5所示,2个生长季节内,GA3喷施浓度对现蕾~成熟期干物质积累量影响极显著(2019年现蕾期除外)。盛花期~成熟期,P0和P1水平下胡麻干物质积累量均随GA3浓度的增加而增加,其中,G2较G0分别提高10.38%~23.63%(盛花期)、11.25%~21.31%(青果期)和8.08%~27.32%(成熟期);P2水平下则为G1最大,各时期分别较G0显著提高8.90%~15.22%、12.73%~15.95%和12.87%~13.28%。增施磷肥显著影响苗期~成熟期干物质积累量,喷施GA3后(盛花期~成熟期),同一GA3水平下,P1和P2分别较P0增加8.36%~41.19%和2.39%~19.73%。互作效应分析表明,磷肥和GA3配合施用对干物质积累量的影响在2019年青果期和成熟期达显著水平,在2020年现蕾期~成熟期达极显著水平,其中,P2G1处理在2个生长季盛花期~成熟期均表现良好,较P0G0处理显著提高4.00%~58.69%。结果表明施用磷肥135 kg·hm-2配合现蕾期和盛花期喷施15 mg·L-1浓度GA3有利于促进胡麻干物质积累量,为胡麻高产奠定了基础。
2.4 磷肥和外源GA3对胡麻产量的影响
由表6可知,不同施磷量之间,2个生长季胡麻籽粒产量均随施磷量的增加而增加,P1和P2分别较不施磷(P0)显著增加9.20%~10.13%和10.33%~11.27%。不同施磷水平下籽粒产量对GA3喷施浓度的响应不同,P1水平下籽粒产量随GA3喷施浓度的增大而升高,且G2较G0显著提高7.64%~9.04%;P2水平下则为G1较G0显著提高3.68%~8.66%。磷肥与GA3互作对胡麻籽粒产量的影响达极显著水平,其中,P2G1处理增产能力最强(与P1G2处理差异不显著),较P0G0显著提高16.88%~17.64%。表明施磷67.5 kg·hm-2配合喷施30 mg·L-1GA3和施磷135 kg·hm-2配合喷施15 mg·L-1GA3均可作为试区胡麻适宜的高产施肥措施。

表6 不同施磷量和GA3处理对胡麻籽粒产量的影响
2.5 磷肥和外源GA3处理下旱地胡麻产量及干物质积累量与光合酶的相关性分析
由表7可知,除现蕾期喷施GA3后3 d和10 d外,胡麻籽粒产量、干物质积累量与现蕾期、盛花期喷施赤霉素前后的PEP羧化酶活性均呈显著正相关关系;喷施GA3前后,胡麻籽粒产量与现蕾期和盛花期的RuBP羧化酶活性均呈显著正相关关系;除盛花期喷施GA3后3 d外,胡麻干物质积累量与现蕾期、盛花期喷施GA3前后的RuBP酶活性亦呈极显著正相关关系。结果表明,胡麻叶片RuBP羧化酶、PEP羧化酶活性的提高促进了干物质积累量的增加和高产的形成。

表7 PEP羧化酶和RuBP羧化酶与干物质积累量及产量间相关系数
3 讨 论
核酮糖-1,5-二磷酸羧化酶/氧化酶(RuBP)和磷酸烯醇式丙酮酸羧化酶(PEPC)是光合作用的关键酶,其活性高低与光合作用密切相关。其中,RuBP羧化酶既控制着CO2的固定,又制约着碳素向Calvin循环和光呼吸循环分流,其活性直接影响作物光合速率[20];PEP羧化酶作为C4作物的光合关键酶,起着“CO2泵”的作用,其活性直接决定了CO2固定能力的强弱[21]。RuBP羧化酶和PEP羧化酶对磷肥营养具有较高的应激效应,土壤中无机磷被植物吸收后不仅可充当光合作用的底物,还影响RuBP羧化酶的形成[22]。有研究指出,外源激素能显著提高玉米叶片PEP羧化酶活性,增强其固定CO2的能力[23]。本研究中,增施磷肥显著提高了胡麻现蕾期和盛花期叶片RuBP羧化酶和PEP羧化酶活性,但单独施磷67.5 kg·hm-2和135 kg·hm-2处理间差异不显著,这与杨晴等[24]的研究结果一致。本研究还发现,施磷135 kg·hm-2配施15 mg·L-1GA3处理和施磷67.5 kg·hm-2配施30 mg·L-1GA3处理均显著提高了胡麻叶片光合羧化酶活性。究其原因,可能是施磷配合喷施GA3能显著提高植株叶片气孔导度和胞间CO2浓度,促进光能的吸收,从而提高了RuBP羧化酶和PEP羧化酶活性[25]。这说明在基础土壤磷素含量较高条件下继续增施磷肥,配合喷施一定浓度的赤霉素有利于进一步提高胡麻光合酶活性[13,26]。
干物质是作物光合产物的最高形式,干物质积累量与产量呈正相关关系[27]。研究表明,优化磷肥用量是实现胡麻高产的重要途径之一,但缺磷和施磷过多均会抑制胡麻生长,不利于其干物质的累积[28]。本研究中,施用磷肥显著增加了胡麻各生育时期干物质积累量,这与前人研究结果一致[28],但盛花期和成熟期施用67.5 kg·hm-2和135 kg·hm-2磷肥,植株干物质积累量之间差异不显著,可能是因为作物地上部干物质的积累还受外源GA3的影响[29]。外源GA3不仅可以促进植株的生长发育,提高作物功能叶片的光合强度和同化物的运输能力,还能加快干物质在籽粒中的积累,从而提高生物产量和籽粒产量[30]。本研究中,施磷67.5 kg·hm-2水平下盛花期~成熟期干物质积累量均随GA3喷施浓度的增加而增加;磷肥和GA3配合施用处理中,施磷135 kg·hm-2配施15 mg·L-1GA3处理的胡麻干物质积累量较高。这可能是因为增施磷肥配合施用外源GA3提高了胡麻的光合和磷代谢酶活性,进而提高了植株对磷养分的吸收和运输效率,抑制生长素氧化酶活性,促进IAA的生物合成,有利于光合同化物的积累,且作物对GA3的响应因养分供应不同而存在差异[13,31-32]。
有关施磷量和外源激素对作物产量的影响研究已有报道[33-34]。研究表明,施用磷肥能有效地促进胡麻籽粒产量的提高[34],在小黑麦开花期叶面喷施20 mg·L-1GA3显著提高了其千粒重和产量[35]。Khan等[13]研究指出,GA3、N和P的联合施用提高了胡麻千粒重和籽粒产量。本研究中,磷肥配施GA3显著增加了胡麻籽粒产量,且在2个生长季节内籽粒产量均以施磷135 kg·hm-2配施15 mg·L-1GA3处理最高,这与前人研究结果基本一致[13]。相关性分析发现,胡麻现蕾期和盛花期叶片RuBP羧化酶和PEP羧化酶的活性与其籽粒产量均呈极显著正相关关系,表明施磷配合喷施GA3可进一步提高胡麻叶片RuBP羧化酶和PEP羧化酶活性,进而增加籽粒产量。
4 结 论
磷肥和GA3的施用对胡麻叶片光合酶活性、干物质积累量和籽粒产量影响显著。其中,施磷67.5 kg·hm-2配合喷施30 mg·L-1GA3显著提高了胡麻叶片RuBP羧化酶活性、盛花期叶片PEP羧化酶活性和籽粒产量;施磷135 kg·hm-2配合喷施15 mg·L-1GA3显著提高了现蕾期和盛花期叶片RuBP羧化酶和PEP羧化酶活性,促进了全生育期干物质积累,进而增加了籽粒产量。可见,施磷67.5 kg·hm-2配合喷施30 mg·L-1GA3和施磷135 kg·hm-2配合喷施15 mg·L-1GA3均可作为试区适宜胡麻的高产施肥管理模式。
