环境内分泌干扰物暴露影响神经元突触可塑性的研究进展
2023-08-02喻华雯
喻华雯,付 娜,许 洁
(遵义医科大学 公共卫生学院营养与食品卫生学教研室,贵州 遵义 563099)
突触是神经元之间发生功能连接和信息传导的关键部位。突触可塑性(synaptic plasticity)指突触在一定条件下变化功能和形态的能力。突触可塑性的改变主要表现为突触结构性和功能性失调。突触是由突触前膜、突触间隙、突触后膜组成的,其中包含多种超微结构。突触结构性失调主要是指突触的面积、数量、体积以及各种超微结构等发生的变化,突触的主要功能是连接细胞、传递信息,当突触可塑性的改变发生在动物的海马等与学习记忆有关的脑区时,突触功能性失调是指动物的学习记忆功能发生障碍。突触可塑性是发挥正常学习记忆功能的前提,两者相互影响,学习记忆功能的改变又会反过来引起突触可塑性的改变,从而形成新的神经回路[1]。有研究表明,突触形态和功能的改变可以导致神经行为异常[2]。有研究表明,突触可塑性的改变会引起大脑中特定部位的损伤从而增加阿尔兹海默病(alzheimer’s disease,AD)、亨廷顿病(huntington’s disease,HD)以及帕金森病(parkinson’s disease,PD)等神经退行性疾病发生的风险[3]。目前已明确,环境内分泌干扰物的暴露是诱导突触可塑性改变的诱因之一。
环境内分泌干扰物(endocrine disrupting chemicals, EDCs)是指可以通过干扰生物或人体内保持自身平衡和调节发育过程的天然激素的运输、结合、反应和代谢等过程从而对生物或人体的生殖、免疫、神经系统等功能产生影响的外源性化学物质[4]。生活中常见的环境内分泌干扰物有:双酚类、邻苯二甲酸酯类(PAEs)、农药、金属及金属混合物、壬基酚(nonylphenol,NP)等。既往研究[5-7]证实EDCs 暴露会引起神经退行性疾病的发生,其病理基础之一是突触可塑性的改变。突触相关蛋白的表达、NMDA受体、膜上动作电位及电流阈值、炎症因子的表达、葡萄糖转运蛋白等的变化都有可能造成突触可塑性的改变(图1)。

图1 EDCs对突触可塑性的改变及机制
EDCs 暴露引起突触可塑性的改变,突触结构性失调可表现为突触的数量减少、突触的面积密度和体积密度降低、突触前膜、后膜以及间隙变得模糊不清,突触间隙增宽、突触小泡数量减少且分布无序,突触活性带长度缩短,突触后膜上的致密物分布散漫、厚度降低及长度缩短,突触界面曲率降低;突触功能性失调的表现有正常的突触传递受限、联系细胞异常、认知功能障碍。通常采用电生理实验、行为学实验、透射电镜、免疫组化实验等观察突触可塑性的改变。突触可塑性的改变影响学习记忆功能和认知功能、影响神经元兴奋性与抑制性之间的平衡、影响神经递质的释放与回收,进而在诱发神经退行性疾病中发挥重要作用(图2)。本文就不同类型的EDCs 从不同途径对突触可塑性的改变进行综述,旨在为神经退行性疾病发生发展的机制提供研究方向,并探讨预防和治疗神经退行性疾病的新靶点。
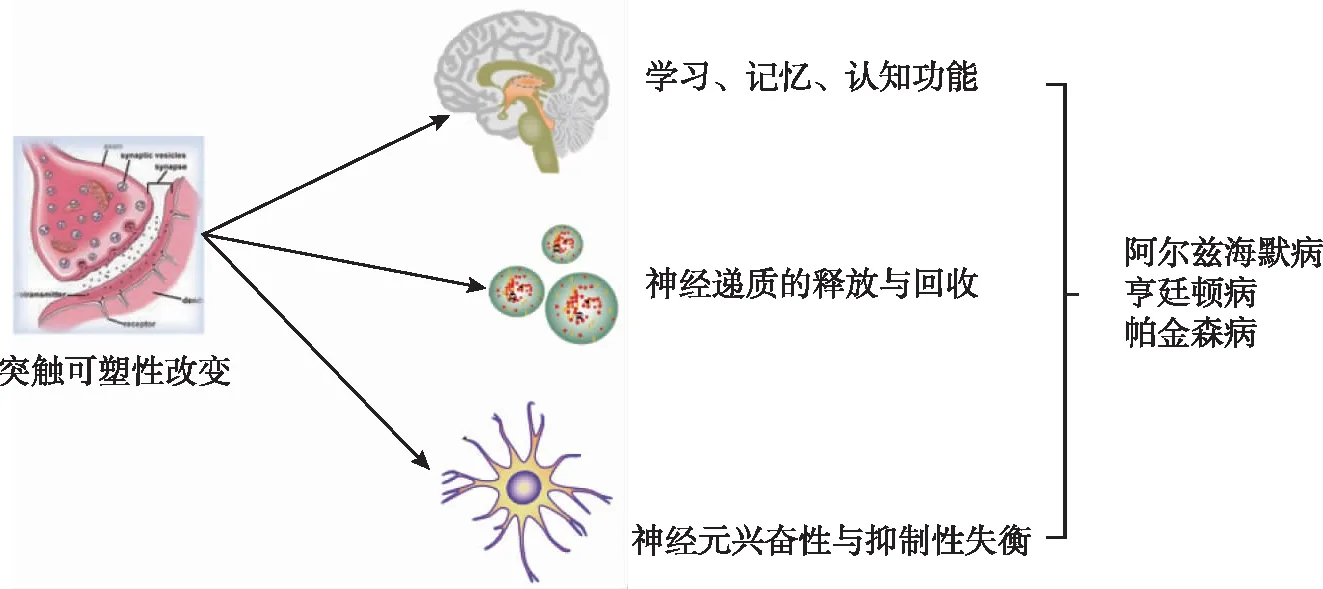
图2 突触可塑性改变对神经系统功能的影响
1 EDCs 对突触可塑性的作用机制
1.1 双酚类 双酚类物质广泛地应用于各行各业中,其典型代表-双酚A(bisphenol A, BPA)作为增塑剂主要用于食品饮料包装、医疗器械包装、牙科材料等[8]。BPA 普遍存在各种环境介质中,在人体的器官和组织中都有积累,既往研究证实 BPA 具有多种毒性。暴露于 BPA 青春期大鼠有空间学习记忆功能障碍以及海马区树突棘密度的降低[9]。在黑腹果蝇的研究中发现,BPA 暴露增加了轴突分支的数量,下调了与突触发育、学习记忆相关的基因表达,同样会引起认知功能障碍[10]。Lee等[11]发现BPA 暴露会减少神经元数量,抑制突触形成,阻碍突触传递,同时孕鼠暴露 BPA 会引起成年后代的神经行为异常。Wu等[12]研究发现,围产期BPA暴露的后代的空间记忆和识别记忆功能均受损,仔代海马 CA1 区的锥体细胞的数量和层数减少且神经元排列松散和不规则,此外仔代海马 PKC/ERK/CREB 信号通路相关蛋白表达降低。Zhang等[13]研究学者发现,围产期暴露 BPAF 会引起雄性后代海马 CA1 区和 DG 区神经元的树突状分枝和复杂性均损伤,以性别依赖的方式诱导雄性子代小鼠的突触可塑性损伤以及认知功能障碍。Hyun等[14]研究在 BPA 染毒皮质锥体神经元的研究中发现,BPA 暴露引起突触的大小、数量、密度的降低,同时正常的突触传递和认知行为遭到破坏。BPA 暴露会通过多种途径引起突触结构性和功能性失调,诱导突触可塑性减弱,进而损害正常的认知功能及神经行为。
突触后密度蛋白95(postsynaptic density 95,PSD-95)和突触素(synapsin,SYP)是典型的突触相关蛋白,突触可塑性的改变不光有形态、数量上的变化,还有分子、蛋白层面的改变。PSD-95 是突触后膜上的一类蛋白,其形成和活动依赖性调节在突触可塑性的改变中发挥重要作用[15]。SYP是突触囊泡膜上的特异性蛋白,其密度和分布可间接反应体内突触的数量和分布情况[16]。BPA[(5、50 mg/(kg·d)]暴露后发现可通过 NMDAR/PSD-95/nNOS-NO-cGMP途径下调PSD-95 蛋白和 mRNA 的水平,突触后密度降低,从而引起空间学习记忆的障碍[17]。Yin等[18]通过 BPA 染毒Neuro-2a 细胞发现,BPA 暴露不仅会导致突触数量的降低、突触完整性受到破坏,同时 SYP 表达上调。Wang等[19]通过不同浓度的 BPA(0、0.1、1.0、10 μM)长期暴露于人类胚胎干细胞后发现,人类胚胎干细胞以浓度依赖性方式抑制神经的生长发育,同时BPA 暴露上调PSD-95 及SYP mRNA及蛋白的表达,以及造成突触棘的退化,从而引发人类胚胎干细胞的神经毒性作用,此外这些 BAP 介导的神经毒性可能是通过增加一氧化氮合酶(iNOS)、神经元一氧化氮合酶(nNOS)3、活性氮(RNS)和活性氧(ROS)的产生以及硝基酪氨酸表达和 Ca2+涌入导致的。突触相关蛋白表达的异常会引起突触结构性和功能性的改变,进而对突触可塑性发挥正常功能产生影响。
N-甲基-D-天冬氨酸受体(NMDA受体)是一种亚型谷氨酸受体,在神经系统的发育过程扮演重要角色,比如:参与调节神经元存活、影响神经元结构,参与形成神经回路,以及影响突触可塑性的变化等;NMDA 受体依赖的突触可塑性对发挥正常学习记忆功能具有重要意义[20]。NMDA 受体通常分布于神经元的突触后膜上,结合谷氨酸后打开离子通路,进而完成 NMDA 受体介导的信息传递。一旦 NMDA 受体的表达改变,突触可塑性受到影响,正常的突触传递受到抑制。暴露于 BPA 的大鼠显著性下调 NMDA 受体[21]。Yu等[17]的研究中同样证实了暴露于 BPA 会下调 NMDA受体(NR1、NR2A)和突触相关蛋白的表达,进而影响突触可塑性。在 Wang等[22]的研究中发现,母体暴露于 BPA 会引起子代 NMDA 受体表达的改变,以及造成子代学习记忆功能障碍。NMDA 受体表达的下降导致突触可塑性的降低,从而损害学习记忆功能。
1.2 邻苯二甲酸酯类 邻苯二甲酸酯类可对神经系统造成损害,其原因可能是突触数量、形态等的改变从而引起的神经疾病的发生。邻苯二甲酸脂二(2-乙基己基)酯(Di-2-ethylhexyl phthalate,DEHP)是一种典型的邻苯二甲酸酯类,DEHP 暴露损害雄性子代海马的突触可塑性,破坏突触超微结构,降低突触相关蛋白水平,说明暴露于DEHP会导致神经发育障碍和学习记忆障碍;DEHP 暴露下调 Rac1/PAK/LIMK1/cofilin信号通路,该通路可能是通过改变细胞骨架结构来参与构建突触可塑性[23]。通过灌胃 DEHP 妊娠期SD 大鼠发现F1 子代大鼠脑组织中 NMDA 受体以及突触素蛋白表达降低[24]。此外,邻苯二甲酸酯类还可以通过改变突触膜上的动作电位、电流阈值以及影响炎症因子表达或释放对突触可塑性产生影响。
突触在传递信息的过程中,突触前膜率先发生电位变化,进而打开离子通道、释放神经递质,神经递质作用于突触后膜并在突触后膜形成突出后电位,即完成突触传递。在 Ran等[25]的研究中发现,DEHP 暴露不仅会显著降低 CA3 锥体细胞的微兴奋性突触后电流(mEPSC)频率,还有CA1 锥体细胞的电流阈值以及动作电位;此外,DEHP 能抑制CA1锥体细胞的电压门控钾通道,进而诱导突触可塑性的抑制。邻苯二甲酸单(2-乙基己基)酯(MEHP)是DEHP的主要活性代谢物之一。在 Lu等[26]的研究中发现,MEHP暴露降低了 CA3神经元sAP的振幅和mEPSCs的频率;此外,MEHP(100 μM和300 μM)显著降低了Ca2+通道的峰值电流密度,仅在300 μM的浓度下降低了Na+和 K+通道的峰值电流密度,进而降低大鼠的突触可塑性及影响神经元兴奋性,从而影响突触之间正常的功能连接和信号传递。
炎症因子在突触可塑性中的作用逐渐被认识,在Khalifa等[27]的研究中发现,DEHP(50、300、750 mg/kg)暴露激活 TLR4/NF-κB 信号通路并增加了炎症因子(IL-1β、IL-6、TNF-α)的表达,且炎症因子的表达随着暴露剂量的增加而增加;此外中剂量组和高剂量组的学习记忆能力显著低于对照组。
1.3 金属及金属混合物 金属在生活中的生产和应用非常广泛,其对人们产生的危害逐渐成为人们关注的热点问题之一。有研究表明,在人类环境接触相关的水平上,铅、镉、汞的混合物暴露会直接损伤大鼠正常的神经行为;此外,随着暴露剂量的增加大鼠海马CA1、DG、CA3区域树突棘密度降低,且树突棘类型改变,成熟蘑菇状棘减少、不成熟的粗短状棘增多[28]。在Pb暴露海马神经元的研究中发现,Pb暴露增加mIPSC频率,降低mEPSC频率,抑制性突触前斑点数量显着增加,而兴奋性突触前末梢数量减少;此外,加强 GABA 的合成并增强海马神经元中突触后 GABAAR 簇的表面分布,进而破坏突触兴奋性和抑制性之间的平衡[29]。在郭克楚等[30]的研究中发现,低剂量Pb、Cd和Hg 混合物暴露即可破坏大鼠正常的学习记忆功能以及下调 NMDA 受体的表达;同时,NR1和NR2A的蛋白表达水平随Pb、Cd和Hg 混合物暴露剂量的增加呈现浓度依赖性降低。此外,还可以通过影响炎症因子表达或释放以及细胞膜葡萄糖转运蛋白来影响突触可塑性。
促炎细胞因子肿瘤坏死因子(TNF)是炎症因子的典型代表,有研究证实[31],高浓度的TNF(1 μg/mL)会造成小鼠CA1海马神经元的损伤,从而造成突触可塑性的改变,这一改变依赖于细胞内钙储存。锌混合物暴露会降低TNF-α 的表达,以及上调PSD-95 和 SYP 的水平,造成突触可塑性的改变,进而损伤大鼠学习记忆功能[32]。
葡萄糖转运蛋白(GLUT)是细胞转运葡萄糖的载体,其中包含 GLUT1、GLUT2、GLUT3、GLUT4 等多种蛋白,GLUT4 在胰岛素的刺激易位到细胞膜上并转运葡萄糖。细胞在分解葡萄糖时会产生大量的 ATP,而 ATP 是突触发生功能连接和信号传递的重要能量来源。有研究证实[33],Pb暴露会引起葡萄糖代谢紊乱,导致脑能量代谢紊乱,进而增加 AD 的患病风险。在 Zhao等[34]研究中发现,妊娠期低水平Pb暴露会引起子代大鼠空间学习及记忆障碍,降低海马 CA1 区的树突棘密度;铅暴露通过磷脂酰肌醇 3 激酶-蛋白激酶 B(PI3K-Akt)通路降低细胞膜中的 GLUT4 水平,从而降低海马葡萄糖代谢,影响突触之间的信号传递。
1.4 农药 农药在生活中使用广泛,其对环境的污染问题已成为现在人们关注的热点。既往研究表明[35-38],接触农药不仅会对环境产生影响,还会对呼吸、心血管、神经系统造成损害。鱼藤酮作为一种常见的杀虫剂在生活中得到广泛地应用。有研究表明,鱼藤酮暴露会造成小鼠记忆损伤,导致小鼠的 GABA 电流减少,从而降低抑制性突触传递[39]。鱼藤酮暴露引起α-突触核蛋白聚集,增加小鼠中脑的细胞因子的表达,破坏单胺类神经递质稳态,进而造成突触可塑性的改变[40]。此外,鱼藤酮暴露会诱导大鼠记忆缺陷,增加大鼠纹状体、前额叶皮层、海马体的TNF-α 和 IL-6 的水平[41]。围产期暴露杀虫剂会引发子代的神经发育障碍及神经行为异常[42]。百草枯曾作为世界上使用最广泛的除草剂之一,但其对人体的危害极大,现已被大多国家禁用。通过对大鼠暴露于百草枯发现,大鼠脑干的多巴胺降低以及突触核蛋白的改变[43]。多巴胺作为一种神经递质与突触核蛋白作为一种典型的突触相关蛋白在调节突触可塑性中发挥重要作用,百草枯的暴露会影响突触可塑性的调节,从而引发突触结构性和功能性失调。
1.5 壬基酚 NP作为一种常见的 EDCs 广泛应用于建材、造纸、洗涤剂、农药、涂料等行业。NP具有持久性和生物蓄积性,一旦 NP 排入环境中会对人体健康造成长时间持续性伤害[44]。有研究发现,NP 暴露大鼠的神经元发生皱缩、胞核固缩,数量减少,突触棘密度降低,突触间隙增宽等变化,且随着 NP 暴露剂量和时间的增加,PSD-95 和synapsin-1的表达愈发降低,突触结构性失调越显著[45]。在 Fu等[46]的研究中发现,NP 染毒引起树突排列紊乱、树突棘密度减少,突触后密度松散且厚度与长度缩短,突触间隙的长度与宽度增加,同时还发现 NMDA受体表达的降低;此外NP暴露可造成大鼠学习记忆功能和空间记忆功能损伤。在NP 染毒原代神经元的研究中发现,NP 暴露引发轴突长度缩短和功能损伤;此外,发育期 NP 暴露可激活子代海马中的Wnt-Dvl-GSK-3β信号通路,损伤子代的轴突发育[47]。NP 暴露不光影响正常的突触结构性和功能性,还会对子代产生神经毒性作用。
2 EDCs通过调节突触可塑性参与认知功能障碍发生的可能机制
2.1 突触可塑性改变对学习记忆功能和认知功能的影响 突触可塑性是维持正常学习记忆功能和认知功能的重要基础[48]。EDCs 暴露可以通过多种途径改变突触可塑性,即导致突触的结构性和功能性失调。在 Dong等[23]的研究中同样证实了这一观点,在突触传递的过程中,树突棘的微小变化将改变突触的功能,从而影响学习和记忆;与对照组相比,DEHP 处理组[300、750 mg/(kg·d)]仔代海马 CA1 区锥体神经元的树突棘密度明显降低,同时DEHP处理组的成熟蘑菇状棘更少,未成熟丝状棘更多;说明DEHP暴露影响了雄性后代的树突状棘发育。在Hyun等[14]的研究中发现类似报道,与对照组相比,BPA 处理组的树突棘的数量减少、长度缩短,同时丝状伪足样刺增加,细刺、蘑菇刺和短刺的数量减少,进而影响突触的形态与功能。既往研究中有很多类似报道,EDCs 的暴露引起突触的结构、数量、面积、体积等变化,进而影响正常的突触传递、改变突触功能,减弱突触可塑性。突触的结构性或功能性失调发生在海马等与学习记忆相关的脑区会影响正常的学习记忆功能和认知功能,从而引发 AD 等神经退行性疾病[49]。
2.2 突触可塑性改变导致神经元兴奋性与抑制性失衡 EDCs 的暴露会引起突触可塑性结构性和功能性失调。突触可塑性的改变会调节神经元兴奋性[50]。在Ran等[25]的研究中发现,暴露于 DEHP[100、300 mg/(kg·d)]可以通过抑制电压门控钾通道抑制神经元的兴奋性。在Zou等[29]的研究中发现Pb暴露会改变突触可塑性,破坏神经元兴奋性与抑制性之间的平衡。保持神经元兴奋性和抑制性之间的平衡是维持正常脑功能的前提,一旦失衡,会增加癫痫、PD 等多种神经疾病患病风险,而突触可塑性在保持这种平衡中扮演重要角色[51]。有研究指出[52],突触可塑性是神经网络建立神经元兴奋性和抑制性平衡的有效机制。突触可塑性失调引起神经元兴奋性和抑制性之间的平衡受到破坏,使正常发挥脑功能受限,发生认知功能障碍。
2.3 突触可塑性改变影响神经递质的释放与回收 现在越来越多的研究表明神经递质与学习记忆有关,与突触可塑性有关。谷氨酸是常见的神经递质,暴露于BPA的大鼠发现谷氨酸受体,其中 AMPA受体1 显著下调,且 Ca2+浓度随着 BPA暴露浓度的增加而增加[19]。在 Hu 等[21]的研究中同样发现 BPA 暴露后GIuR1的表达降低,海马组织中谷氨酸水平降低,海马 CA1 区的突触前递质合成与释放减少。此外,鱼藤酮暴露可引起单胺类神经递质释放与回收紊乱[40]。突触可塑性改变导致突触之间的联系、信息传递减少,神经递质受体减少会导致神经递质的释放与回收发生障碍,神经递质的是释放与回收是突触在完成正常信息传递和功能连接中的关键一环,其异常又会反过来对突触可塑性产生影响。
3 展望
随着科学技术的发展,研究学者对 EDCs 的危害性的研究逐渐增多。EDCs 暴露会引起神经行为功能紊乱,其病理基础之一是神经元突触可塑性的改变。EDCs暴露可以通过改变突触相关蛋白的表达及 NMDA 受体的表达、改变膜上动作电位和电流阈值、影响炎症因子的表达与释放以及降低 GLUT 的活性等多种途径对突触可塑性造成影响,从而造成认知功能障碍,进而增加 AD、PD、HD 等神经退行性疾病的患病风险。突触在发挥正常功能时,需要多个环节共同参与,例如,突触膜上的受体含量、突触膜内外的离子浓度、ATP的释放与回收、神经递质的释放与回收等,且不同类别不同位置上的突触可塑性的改变是不同的,各突触之间的连接方式也不尽相同,所以诱导突触的数量、面积、体积、超微结构等发生改变,以及神经元连接异常和信息传导异常的因素很多,这些改变是通过Rac1/PAK/LIMK1/cofilin信号通路,或者PI3K-Akt 信号通路,亦或者其他通路来介导的,有待进一步研究。这对探讨神经退行性疾病发生发展的机制提供参考价值,也为以后探讨EDCs 诱导神经疾病的发病机制展开更加深入的研究奠定了基础。
