泗洪地区克氏原螯虾病原维氏气单胞菌鉴定及致病性分析
2023-07-31汤环宇钱且奇徐静雯朱玉洁许文静姬进宝张晓君
汤环宇,钱且奇,陈 圳,徐静雯,朱玉洁,许文静,姬进宝,高 策,张晓君
(扬州大学动物科学与技术学院,江苏扬州 225009)
克氏原螯虾(Procambarusclarkii),俗称小龙虾,隶属甲壳纲十足目螯虾科,具有食性杂、养殖周期短、环境适应能力强等特点,具有较高的经济价值,是我国常见的水产养殖品种之一[1]。自1929年从日本引入国内后,克氏原螯虾养殖产业进入快速发展期,已成为一些地区的支柱性产业及重要经济来源[2]。同时,随着养殖规模的不断扩大及水环境的急剧恶化,小龙虾病害问题日趋频繁和严重。近年来,细菌性病害已成为克氏原螯虾产业发展的主要制约因素,且病原细菌呈现种类多、致病性强、发病率高和耐药性增强等特点。目前已有报道可引起克氏原螯虾暴发性死亡的病原菌主要包括豚鼠气单胞菌(Aeromonascaviae)[3]、嗜水气单胞菌(A.hydrophila)[4]、维氏气单胞菌(A.veronii)[5]、弗氏柠檬酸杆菌(Citrobacterfreundii)[6]等,病虾表现为食欲减退、行动迟缓、活力减弱,鳃部附着大量污物等。
气单胞菌属是我国淡水养殖业常见的致病源之一,易引起多种水生养殖动物的大规模死亡[7]。维氏气单胞菌隶属于气单胞菌科气单胞菌属,是一种广泛存在于淡水、海水、淤泥和土壤中的条件致病菌,感染鱼类后主要引发细菌性败血症,以引起皮肤溃烂及坏死为典型症状[8]。已有报道该菌可导致草鱼(Ctenopharyngodonidella)[9]、鲫(Carassiusauratus)[10]、尼罗罗非鱼(Oreochromisniloticus)[11]、黄颡鱼(Pelteobagrusfulvidraco)[12]、黄鳝(Monopterusalbus)[13]、锦鲤(P.fulvidraco)[14]、大口黑鲈(Micropterussalmoides)[15]、斑点叉尾鮰(Ictaluruspunctatus)[16]等鱼类发病死亡。此外,维氏气单胞菌也会引起中华绒螯蟹(Eriocheirsinensis)[17]、凡纳滨对虾(Litopenaeusvannamei)[18]、青虾(Macrobrachiummipponensis)[19]等甲壳类水生养殖动物大量死亡。周慧华等[20]在2017年6月从患病中华绒螯蟹中分离出致病性维氏气单胞菌;姜光明等[21]报道,2016年也从江苏某地患菌血症的克氏原螯虾上分离到强致病性的维氏气单胞菌;胡骞等[22]报道维氏气单胞菌感染克氏原螯虾后可导致其反应迟钝,引起肝胰腺等组织损伤,且死亡数量大。 5-7月是克氏原螯虾养殖与出售的关键时期,而此时水温与维氏气单胞菌的最适温度区间相重合,正是维氏气单胞菌繁殖最为旺盛的时候,这使得维氏气单胞菌对克氏原螯虾养殖业的危害更甚于其他水产品。
已有研究表明,维氏气单胞菌致病性受到其毒力因子分泌、表达的控制,如外毒素、肠毒素、胞外蛋白酶和各种黏附因子等[23]。其中溶血素、胞外蛋白酶是维氏气单胞菌的重要毒力因子,在病原菌的致病过程中起到决定性的作用。罗志飞等[24]曾通过研究发现气单胞菌弱毒株培养上清均不溶血、不溶蛋白,而强毒株具有强溶血活性和溶蛋白活性。
维氏气单胞菌致病机制较为复杂,可能是由多个毒力因子共同作用所致。因此,检测分离菌所携带毒力因子,对揭示其潜在致病性至关重要。溶血素是维氏气单胞菌的重要毒力因子,它通过改变细胞膜的通透性,进而使红细胞破裂溶解来表现出溶血性和肠毒性,而且溶血素还会破坏其他细胞,且这种破坏是不可逆转的[25]。目前,关于维氏气单胞菌溶血素的研究较少,SREEDHARAN等[26]从濒死的鱼类分离出的维氏气单胞菌均含有aerA和hlyA基因,研究表明这些基因能对宿主细胞的损伤和死亡造成一定影响,而本次实验中也在分离菌CPA2020中发现了hly基因,证明这也是克氏原螯虾致病过程中维氏气单胞菌毒力的重要来源之一。此外,各种胞外酶活性也是气单胞菌毒力的重要因素[27]。胞外蛋白酶是一类广泛存在的酶,在细菌致病性、应激反应和细胞活力等方面发挥着重要作用。明胶酶是一种潜在的毒性因子,它缺乏明显的底物特异性,可水解酪蛋白、血红蛋白、胶原蛋白等多肽,造成广泛的组织损伤,并有助于细菌在高密度环境中的传播。同时,毒力基因检测从致病菌CPA2020中还发现脂肪酶基因lip,NAWAZ等[28]在对鲶鱼毒力因子的研究中发现,脂肪酶会破坏红细胞的细胞膜,致使细胞发生溶解。外膜蛋白(outer membrane proteins,Omps)是革兰氏阴性菌外膜的主要结构,与维持细胞形态、物质代谢、组织黏附及致病等密切相关[29]。王薇等[30]的研究证实了OmpA是拟态弧菌的一种重要黏附素,它通过黏附参与致病作用,是一种重要的毒力因子。Namba等[31]报道,从鲤肠道中分离出来的维氏气单胞菌CWP11菌株产生的外膜蛋白OMPA是一种黏附因子,经克隆和核苷酸测序发现OmpA存在2个串联的OmpAⅠ-OmpAⅡ同系物,利用同源重组技术缺失该菌株的OmpAⅠ,发现可降低对鲤鱼肠上皮细胞的黏附活性降低,说明OmpAⅠ对肠上皮细胞有较强的黏附作用。
近年来江苏省泗洪多家克氏原螯虾养殖场细菌性病害颇为严重,病虾往往呈现虾体发红、四肢无力、摄食减少、行动迟缓、对外界刺激反应迟钝、肝胰腺水肿等症状,并陆续出现暴发性死亡。为明确该病病原,本研究对江苏省泗洪螯虾主养区的多个典型发病养殖场进行调研与病原检测。通过对分离菌进行致病性、形态特征观察、理化特性鉴定,结合gyrB基因序列测定与系统发育学分析,结果表明引起克氏原螯虾暴发性死亡的病原为维氏气单胞菌。研究结果可为克氏原螯虾细菌性病害的防治及维氏气单胞菌致病机理的深入研究提供参考。
1 材料与方法
1.1 实验虾
发病克氏原螯虾[平均体长(7.0±1.0)cm]取自江苏省泗洪县某克氏原螯虾养殖场,症状为摄食减少、反应迟缓、腹部肌肉肿胀,肝胰腺水肿,颜色加深。用于回归感染实验的健康克氏原螯虾取自江苏省扬州市克氏原螯虾养殖场,本实验采用了封闭循环水养殖系统,养殖用水经曝气2 d,水温(24~25)℃,每日投喂人工饲料1次。
1.2 病原菌的分离与纯化
取患病克氏原螯虾肝胰腺划线接种于LB培养基平板,于28 ℃过夜培养。挑取优势单菌落划线接种于LB培养基上,进一步纯化,28 ℃培养18 h,选取单菌落(菌株编号为CPA2020)接种于 LB液体培养基培养,置于甘油管-40 ℃保存备用。
1.3 病原菌人工感染实验
将纯化的优势菌(CPA2020)接种于LB培养基并于28 ℃过夜培养18 h,用无菌生理盐水调至浓度为1.6×104、1.6×105、1.6×106、1.6×107和1.6×108CFU/mL。选用暂养5 d的健康克氏原螯虾,每个实验组分别注射感染20只健康克氏原螯虾,每只虾注射0.1 mL不同浓度的菌液,同时设立无菌生理盐水作对照,对照组虾注射等量无菌生理盐水。连续观察虾的死亡情况并记录,按Behreans and Karber方法[32]计算半数致死剂量LD50,对濒死虾进行细菌再分离和鉴定。
1.4 病原菌的鉴定
对分离菌CPA2020分别进行革兰氏染色、氧化酶、接触酶、糖(醇及苷)类代谢、硝酸盐还原、O-F实验、动力等理化特性测定,各项理化指标的测定主要参照《Bergey′s Manual of Systematic Bacteriology》[33]进行。细菌微量生化反应管等均购自杭州天和微生物试剂有限公司。
1.5 gyrB基因序列测定与系统发育学构建
1.5.1 PCR模板DNA的制备
取纯培养菌分别接种于含盐1%的营养LB肉汤中28 ℃培养16 h,按TIANGEN细菌基因组DNA提取试剂盒(上海赛百盛基因技术有限公司产)使用说明书提取DNA作为PCR模板DNA,-20 ℃保存待用。
1.5.2gyrB基因序列的PCR扩增与测序
gyrB基因PCR扩增的两个引物分别为:UP1(正向引物):5′-GAAGTCATCATGACCG TTCTGCAYGCNGGNGGNAARTTYGA-3′,UP2r(反向引物):5′-AGCAGGGTACGGATGT GCGAGCCRTCNACRTCNGCRTCNGTCAT-3′。在20 μL反应体系中含有:正反引物各0.5 μL,PCR Mix 10 μL,ddH2O 8 μL,细菌基因组模板1 μL。PCR反应条件为:94 ℃预变性4 min;94 ℃变性1 min,55 ℃退火1 min,72 ℃延伸2 min,共30个循环;72 ℃延伸6 min;16 ℃结束反应程序。扩增产物用1%的琼脂糖凝胶电泳检测,之后交由南京擎科生物技术有限公司进行测序。
1.5.3gyrB基因序列系统发育树构建
对分离菌的gyrB基因序列通过NCBI的Blast进行同源性检索,并使用Clustal X 2.0软件与从GenBank数据库下载相似性较高的菌株的序列进行多序列比对(multiple alignments),并对gyrB基因采用MEGA7.0软件构建系统发生树,选用邻接(neighbor joining method)法构建系统发育树。
1.6 克氏原螯虾的病理组织分析
分别取健康、自然患病及人工感染的克氏原螯虾各5只,将解剖出的克氏原螯虾肝胰腺用波恩氏液固定24 h后,使用70%酒精保存,克氏原螯虾肝胰腺组织经脱水、透明、浸蜡和石蜡包埋一系列操作后,使用切片机连续切片(3~6 μm),之后进行H-E染色,中性树脂胶封片,最后使用显微镜观察记录。
1.7 胞外酶与溶血活性检测
采用平板法测定分离菌CPA2020的卵磷脂酶、脂酶、明胶酶、脲酶以及溶血素活性。吸取10 μL分离菌CPA2020分别点种在含10%卵黄液、1%吐温80、1%明胶、2%尿素(含酚红指示剂)以及7%兔脱纤血的固体培养基上,于28 ℃恒温倒置培养24 h。实验均重复3次,根据是否出现透明圈判定酶活性。若有水解圈产生,则反应为阳性。
1.8 毒力基因检测
根据GenBank已登录的维氏气单胞菌的毒力基因,实验选择flaA、aer、ompA、aha、flaH等16个常见毒力基因,采用Primer Premier 5.0软件设计特异性引物,由上海生物工程技术公司合成(表1),以检测分离菌几种毒力基因的携带情况。
1.9 病原菌耐药性性测定
取供试分离菌,移接于普通营养肉汤中培养18 h,后将菌株悬液均匀涂布于LB琼脂平板上,待自然干燥后平贴各药敏纸片,每个平板均匀分布贴6张,置于28 ℃生化培养箱中培养24 h后观察并记录结果。参照杭州滨和微生物试剂有限公司药敏纸片抑菌圈标准,以测量的抑菌圈直径大小为指标判定其敏感与耐药性。
2 实验结果
2.1 克氏原螯虾疾病发生情况
人工感染后的病虾与自然发病虾的症状相同,表现为行动迟缓,对外界刺激反应迟钝,头胸部与腹部间肌肉松软,头胸甲和肌节容易分离(图1a),外观可见其心脏积液(图1b),腹部肌肉肿胀(图1c),肝胰腺水肿,颜色加深(图1d)。

图1 患病克氏原螯虾临床症状Fig.1 The clinical signs of diseased P.clarkiia:健康虾头部;b:病虾头部;c:健康虾心脏;d:病虾心脏;e:健康虾腹部;f:病虾腹部;g:健康虾肝胰腺;h:患病虾肝胰腺
2.2 病原菌的致病性
低浓度组在观察期内持续发病死亡;对照组克氏原螯虾在5 d的观察期内均未出现死亡(图2)。计算得分离菌对克氏原螯虾的半致死量为6.31×105CFU/mL。证明分离菌对克氏原螯虾具有较高致死率,并从人工感染死亡克氏原螯虾体内分离到大量优势原感染菌。
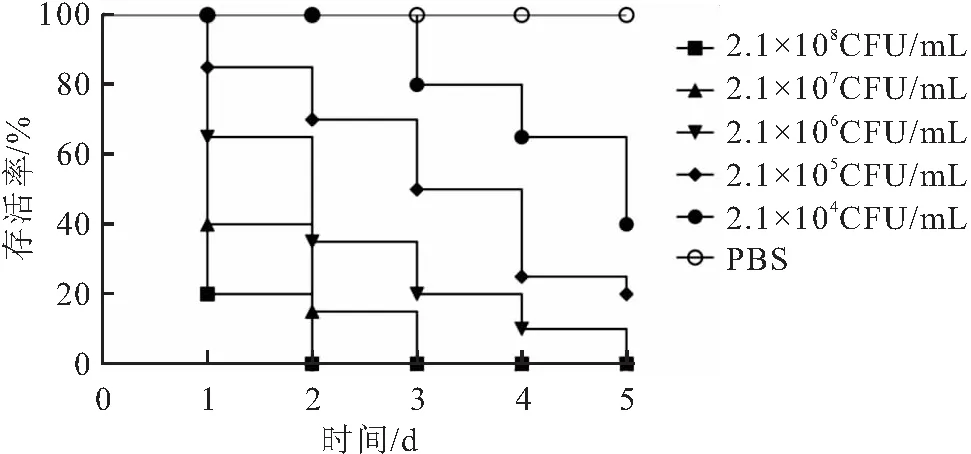
图2 菌株CPA2020对克氏原螯虾人工感染实验结果Fig.2 The artificial infection experiment results of strain CPA2020 on P.clarkii
2.3 病原菌的鉴定
菌株CPA2020的生理生化特性鉴定结果见表2,分离菌能够在37 ℃中生长,具有动力,能利用蔗糖、精氨酸双水解酶、甘露糖等,这与《Bergey′s Manual of Determinative Bacteriology》[33]中维氏气单胞菌的理化特性描述基本一致。初步鉴定菌株CPA2020为维氏气单胞菌。
对其gyrB基因序列进行扩增、测序,在NCBI中进行Blast同源性检索,发现其gyrB基因序列与维氏气单胞菌的同源性最高(100%)。选取不同气单胞菌属的代表株gyrB基因序列构建系统发育树(图3)。结果显示,CPA2020与维氏气单胞菌(R537457.1)聚为一支,综合生理生化检测结果,可以判定病原菌为维氏气单胞菌,并命名其为CPA2020。

图3 基于CPA2020 gyrB序列构建的气单胞菌的系统发育树Fig.3 Phylogenetic tree of Aeromonas based on CPA2020 gyrB sequence
2.3 毒力基因及胞外酶检测
PCR扩增结果图4所示,分离鉴定的病原维氏气单胞菌可扩增出lip、hly、flgN、ompA、flgM、flaH、aha、flgA 8种相关毒力基因,表明菌株CPA2020携带这些毒力因子。

图4 菌株CPA2020毒力基因PCR凝胶电泳结果Fig.4 Virulence genes of strain CPA2020 by PCR amplificationM:Marker;1:lip;2:hly;3:flgN;4:ompA;5:flgM;6:flaH;7:aha;8:flgA
病原菌CPA2020胞外酶检测结果表明,分离菌具有明胶酶、溶血素和卵磷脂酶活性,但不具有脂肪酶和脲酶活性。结果如图5所示。
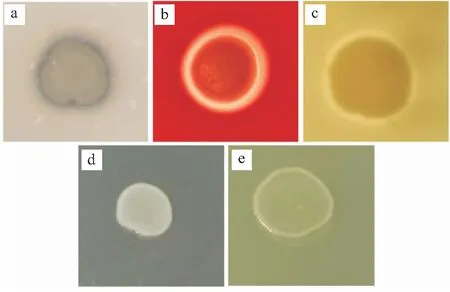
图5 胞外酶和溶血素的检测Fig.5 Determination of extracellular enzymes and hemolysin.a:明胶酶活性;b:溶血素活性;c:卵磷脂酶活性;d:脂肪酶的活性;e:脲酶的活性。
2.4 维氏气单胞菌感染克氏原螯虾的病理学分析
克氏原螯虾感染分离菌CPA2020后,肠道和肝胰腺组织出现明显的组织学改变,产生明显的炎症反应。健康虾的肠道结构形态完整,层次分明(图6A)。自然患病虾的肠道内各组织溶解在一起,肠道组织间出现较大裂隙,皱嵴减少(图6B),人工感染的病虾的肠道内也呈现出皱嵴减少的病理变化(图6C)。健康虾的肝小管形态完整,排列紧密(图6D),而自然患病和人工感染的病虾肝小管间均出现间隙,空泡化严重(图6E、F)。
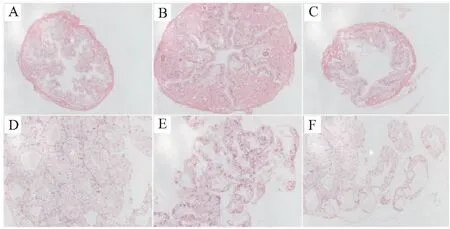
图6 维氏气单胞菌感染克氏原螯虾后肝胰腺与肠组织病理变化(放大倍数,400×)Fig.6 Tissue structure of hepatopancreas and intestine of P.clarkii infected by A.veronii(magnification,400×)A:正常的肠道组织;B:自然患病的肠道组织;C:人工感染的肠道组织;D:正常的肝胰腺组织;E:自然患病的肝胰腺组织;F:人工感染的肝胰腺组织。
2.5 药敏实验
药敏实验结果见表3,菌株CPA2020对链霉素、万古霉素、环丙沙星等12种药物耐药,对头孢唑林、克拉霉素等9种药物中介,对头孢呋辛、左氟沙星、氨曲南等13种药物敏感。
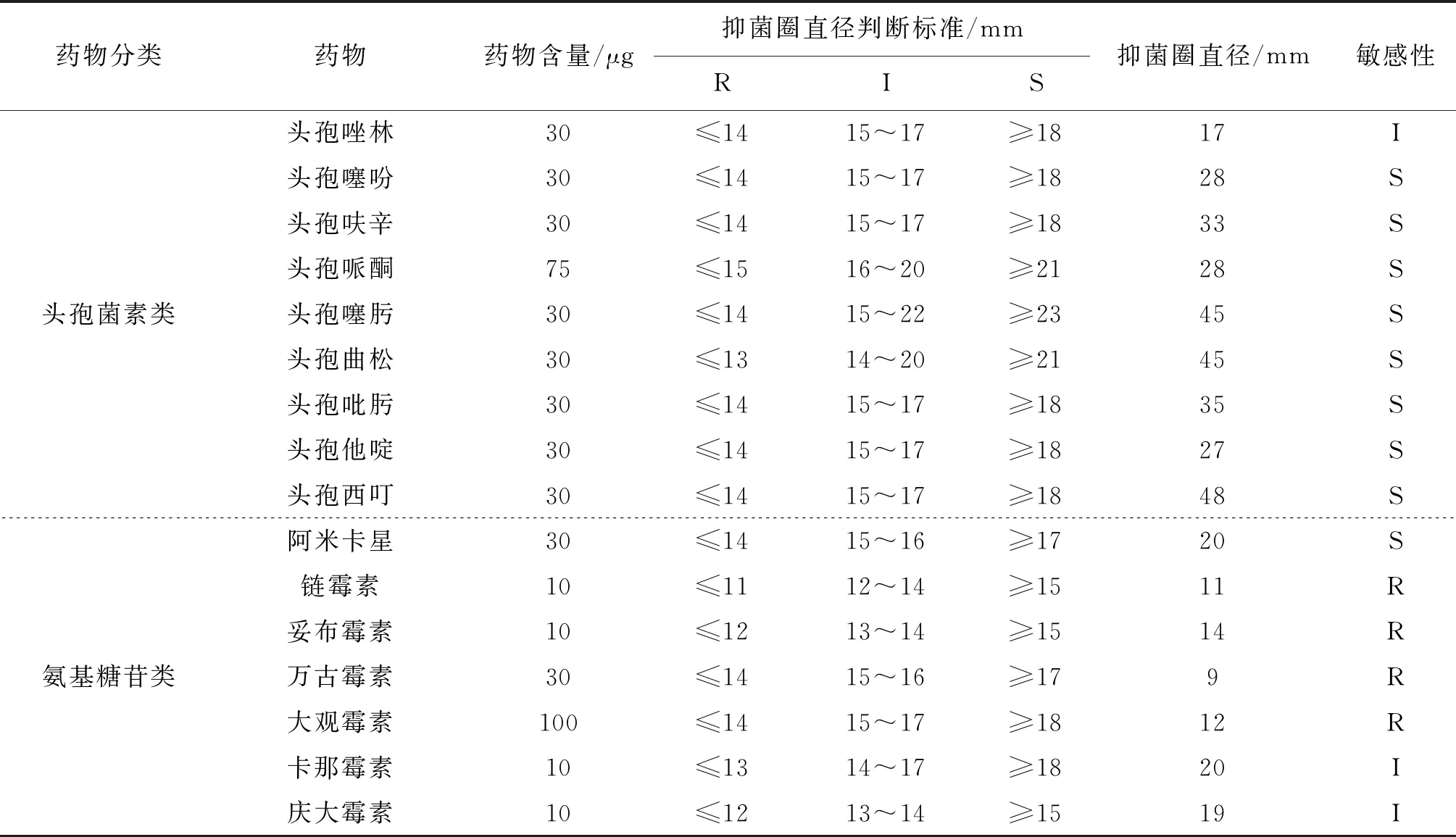
表3 菌株CPA2020的耐药性分析Tab.3 Analysis of drug resistance of strain CPA2020
3 讨论
3.1 维氏气单胞菌亲缘性分析
通过gyrB基因同源性分析,并且构建系统进化发育树,分离菌CPA2020与维氏气单胞菌聚为一支。因此,可鉴定分离菌CPA2020为维氏气单胞菌。此外,不同维氏气单胞菌之间的亲缘性也不同。由图3可知,CPA2020与一株从斑点叉尾鮰鱼上分离出的维氏气单胞菌(KR537457.1)同源性最高,达100%,与GAO等[34]分离出的一株引起青虾大量死亡的维氏气单胞菌(MK905236.1)同缘性很高,而与胡骞等[22]从患病克氏原螯虾肝胰腺中分离到的维氏气单胞菌相似度比较低(MN189983.1),说明不同来源的维氏气单胞菌,其亲缘性存在一定的差异性。
3.2 维氏气单胞菌致病性分析
人工感染实验结果表明,CPA2020可使健康的克氏原螯虾在5 d内全部死亡,说明该菌有较强的致病性。为探究其致病的重要因子,本研究对分离菌CPA2020进行胞外酶检测,结果表明,分离菌CPA2020可分泌卵磷脂酶、明胶酶和溶血素。为进一步了解CPA2020的致病机制,本研究对其进行了毒力基因检测。结果显示菌株CPA2020携带8种毒力因子,进一步揭示菌株CPA2020具有强致病性。同时,这些毒力基因可作为检测致病性类维氏气单胞菌的生物学标记,为分子流行病学研究奠定基础。
3.3 药敏实验结果分析
药敏实验结果显示菌株CPA2020对氨曲南、头孢呋辛、左氟沙星等13种药物敏感,但部分头孢类药物是违禁药品,目前不能广泛应用于临床。对庆大霉素、卡那霉素、克拉霉素等9种药物中度敏感,这与耿昕颖等[36]研究结果部分一致,其他不相同部分可能与菌株来源、用药差异等有关。对复方新诺明、万古霉素、链霉素等12种药物耐药,所以在药物使用时应该结合国家渔业用药准则,有选择地用敏感药物进行治疗,并轮换用药,防止耐药菌株的产生。由于抗菌药物在水产养殖生产中的长期使用,诱导维氏气单胞菌产生耐药性,并且不同地域分离株以及不同水产动物来源分离株的耐药性均有差异。本研究的耐药性分析表明,菌株CPA2020对氨基糖苷类、青霉素类药物耐药。这和蔺凌云等[37]、田甜等[38]的实验结果相似,说明维氏气单胞菌对氨基糖苷类、青霉素类药物的抗药性是较为普遍的。本实验结果显示只有头孢类药物可有效抑制维氏气单胞菌,但其已被禁用。维氏气单胞菌多重耐药现象严重,耐药株的出现是在抗生素选择压力下发生多位点突变所致。当前的养殖过程中因更多仅考虑疾病治愈效果而滥用抗菌药物,致使细菌发生耐药性变异。因此我们亟需研发不产生耐药性的新型药物,如抗细菌毒力药物,选择性地遏制病原菌的黏附、毒素表达、毒素传递和免疫逃避等,从而降低病原菌的毒力,达到有效预防和治疗细菌感染的目的。
综上所述,本研究对此次克氏原螯虾细菌性疾病病原进行分离鉴定及药敏实验,可为我国螯虾产业的疫病防控、合理用药、流行病学调查等研究提供参考。未来仍需进一步开展维氏气单胞菌的致病因子、耐药基因和螯虾免疫相关基因等方面的研究,以期揭示螯虾细菌性疾病的免疫应答规律,从而为其细菌疾病的防控与治疗提供科学依据,保证螯虾养殖产业的可持续发展。
