长吻苗种适宜圈养密度研究
2023-07-31张新福何盛亮黄路全贾智明徐聚臣宗巍巍吕亚兵何绪刚
张新福,杨 军,何盛亮,黄路全,贾智明,徐聚臣,宗巍巍,吕亚兵,侯 杰,3,何绪刚,3
(1.华中农业大学水产学院,武汉 430070;2.湖北省宜昌市水产技术推广站,湖北宜昌 443000;3.长江经济带大宗水生生物产业绿色发展教育部工程研究中心,武汉 430070)
绿色高效圈养模式(简称“圈养模式”)是由华中农业大学基于“时时打扫卫生”的理念提出,采用圈养装备实现高效养殖并及时收集和处理养殖废弃物的一种新型养殖模式[1],因其养殖效率高、“零排放”、产品优质、捕捞方便等突出优势,倍受产业关注,推广应用势头迅猛。长吻(Leiocassislongirostris),俗称江团、肥鱼、肥沱等,具有个体大、生长快、肉质细嫩、营养价值高、味道丰富鲜美、无肌间刺等诸多优点,是一种高价值的淡水名优品种[2]。当前长吻苗种培育以池塘培育为主,但池塘培育存在病害多发、饵料利用率低、存活率低等生产问题[3]。本研究在圈养模式下,探究3种不同养殖密度对长吻苗种生长性能、血清生化指标及非特异性免疫指标的影响,为长吻苗种适宜的圈养密度提供参考依据,同时为后期长吻成鱼的生态、高效、健康养殖提供借鉴。
1 材料与方法
1.1 实验条件
本养殖实验在湖北省枝江市渔丫头水产养殖专业合作社进行,养殖池塘面积为16 675 m2,平均水深为2.5 m,池塘内共架设60个圈养桶,其中9个圈养桶用于本试验。实验所用长吻苗种均为同一批人工繁育所得,规格整齐,生长良好。
1.2 实验设计与饲养管理
1.3 生长数据采集与计算
存活率(SR)=100%(N0-Nt)/N0
增重率(WGR)=100%(Wt-W0)/W0
特定生长率(SGR)=100%(lnWt-lnW0)/t
体质量变异系数(CVw)=100%Ws/Wt
饵料系数(FCR)=FI/(Wt-W0)
摄食率(FR)=100%FI/[0.5(Wt+W0)×t]
肥满度(CF)=100%Wb/L3
其中:W0(g)为鱼体初始均重;Wt(g)为鱼体终末均重;t为饲喂天数;FI(g)为尾均摄食量(湿重);Nt为死亡尾数;N0(g)为放养尾数;Wb(g)、L(cm)分别代表体重和体长,Ws(g)为体质量标准差。
1.4 血清样品采集与处理
在30、60、90 d于每个重复随机取5尾鱼,用MS-222溶液麻醉后,用2.5 mL注射器进行尾静脉取血,4 ℃下静置2 h后,在3 500 r/min,4 ℃条件下离心15 min,取血清放于-80 ℃保存,用于生化、抗氧化及免疫指标测定。
1.5 血清生化、抗氧化及免疫指标的测定
血糖、总蛋白、白蛋白、胆固醇、总甘油三酯、谷草转氨酶、谷丙转氨酶、碱性磷酸酶在Selectra-xl型全自动生化分析仪(荷兰威图公司)上测定,试剂盒购自中生北控生物技术股份有限公司。血清抗氧化指标超氧化物歧化酶、过氧化氢酶、丙二醛及血清免疫指标溶菌酶、补体3、补体4均采用南京建成生物科技研究生产的试剂盒测定,操作步骤严格按照说明书进行。
1.6 数据整理与分析
实验结果均采用平均值±标准差(Mean±SD)表示,用Excel对数据进行常规计算与统计,运用SPSS25.0软件对各组数据进行单因素方差分析处理,P<0.05为差异水平显著,并使用Origin2019绘图。
2 结果
2.1 养殖密度对长吻苗种生长性能的影响
表1 90 d后不同密度组长吻苗种生长情况Tab.1 Growth of the juveniles of L.longirostris in different density groups after 90 days
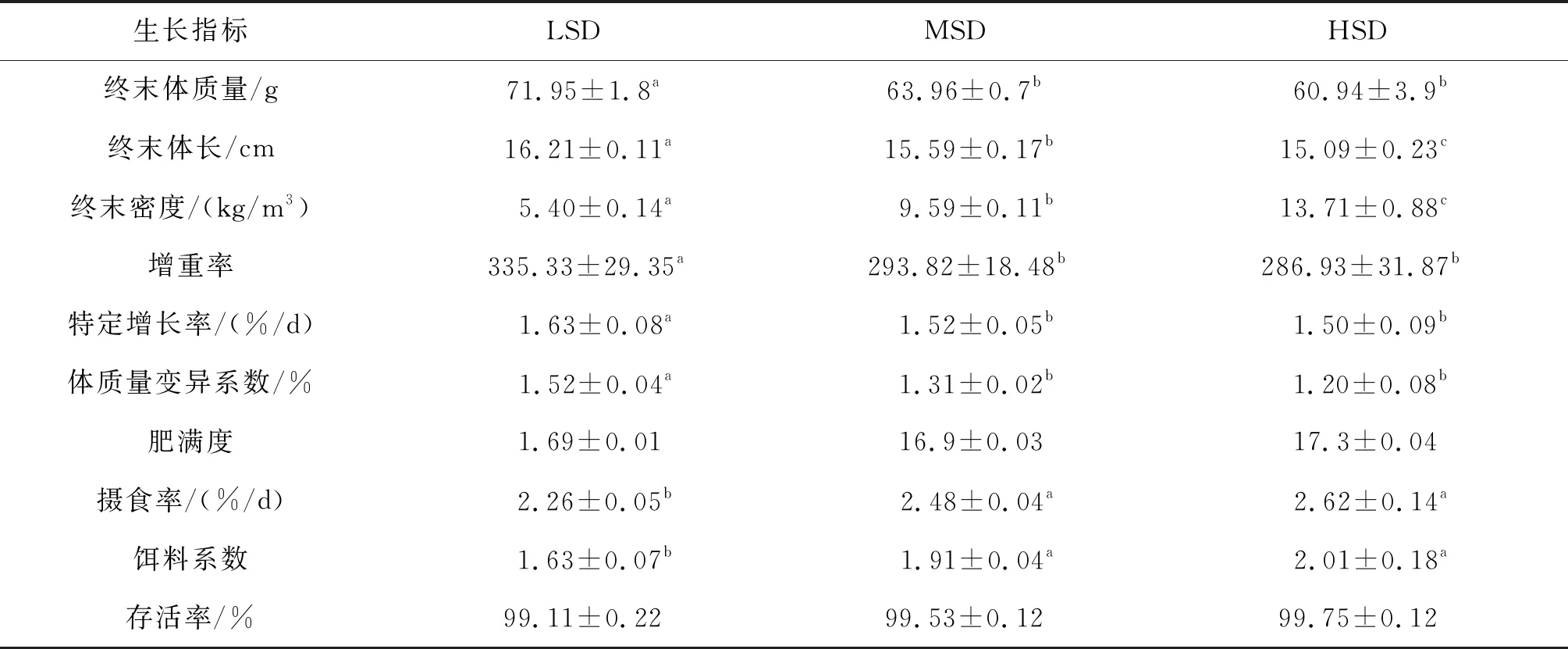
表1 90 d后不同密度组长吻苗种生长情况Tab.1 Growth of the juveniles of L.longirostris in different density groups after 90 days
生长指标LSDMSDHSD终末体质量/g71.95±1.8a 63.96±0.7b 60.94±3.9b终末体长/cm16.21±0.11a15.59±0.17b15.09±0.23c终末密度/(kg/m3)5.40±0.14a9.59±0.11b13.71±0.88c增重率335.33±29.35a293.82±18.48b286.93±31.87b特定增长率/(%/d)1.63±0.08a1.52±0.05b1.50±0.09b体质量变异系数/%1.52±0.04a1.31±0.02b1.20±0.08b肥满度1.69±0.01 16.9±0.03 17.3±0.04 摄食率/(%/d)2.26±0.05b2.48±0.04a2.62±0.14a饵料系数1.63±0.07b1.91±0.04a2.01±0.18a存活率/%99.11±0.22 99.53±0.12 99.75±0.12
注:表中同行不同的上标字母表示组间差异性显著(P<0.05)。下表同。

图1 不同密度组下长吻苗种体质量和体长Fig.1 Body weight and length of the juveniles of L.longirostris under different densities不同字母表示组间差异显著(P<0.05)。
2.2 养殖密度对长吻苗种血清生化指标的影响
血清总蛋白和白蛋白的含量均随着养殖时间进行而逐渐升高,在90 d时,HSD组血清白蛋白和总蛋白含量显著高于LSD、MSD组。血糖含量随着养殖时间推移呈先升高后降低的趋势,且各密度组间的差异不显著;总甘油三脂和胆固醇水平随着养殖时间而逐渐升高,且各密度组两者均差异不显著;血清中谷草转氨酶含量变化较小,且差异不显著;谷丙转氨酶含量随着养殖时间推进而升高,养殖后期其含量随着养殖密度升高而降低,但不呈显著性差异;LSD组碱性磷酸酶含量高于其余两组,但差异不显著,90 d各密度组碱性磷酸酶含量略有升高(表2)。
表2 各密度组长吻苗种血清生化指标Tab.2 Serum biochemical indexes of the juveniles of L.longirostris under different densities
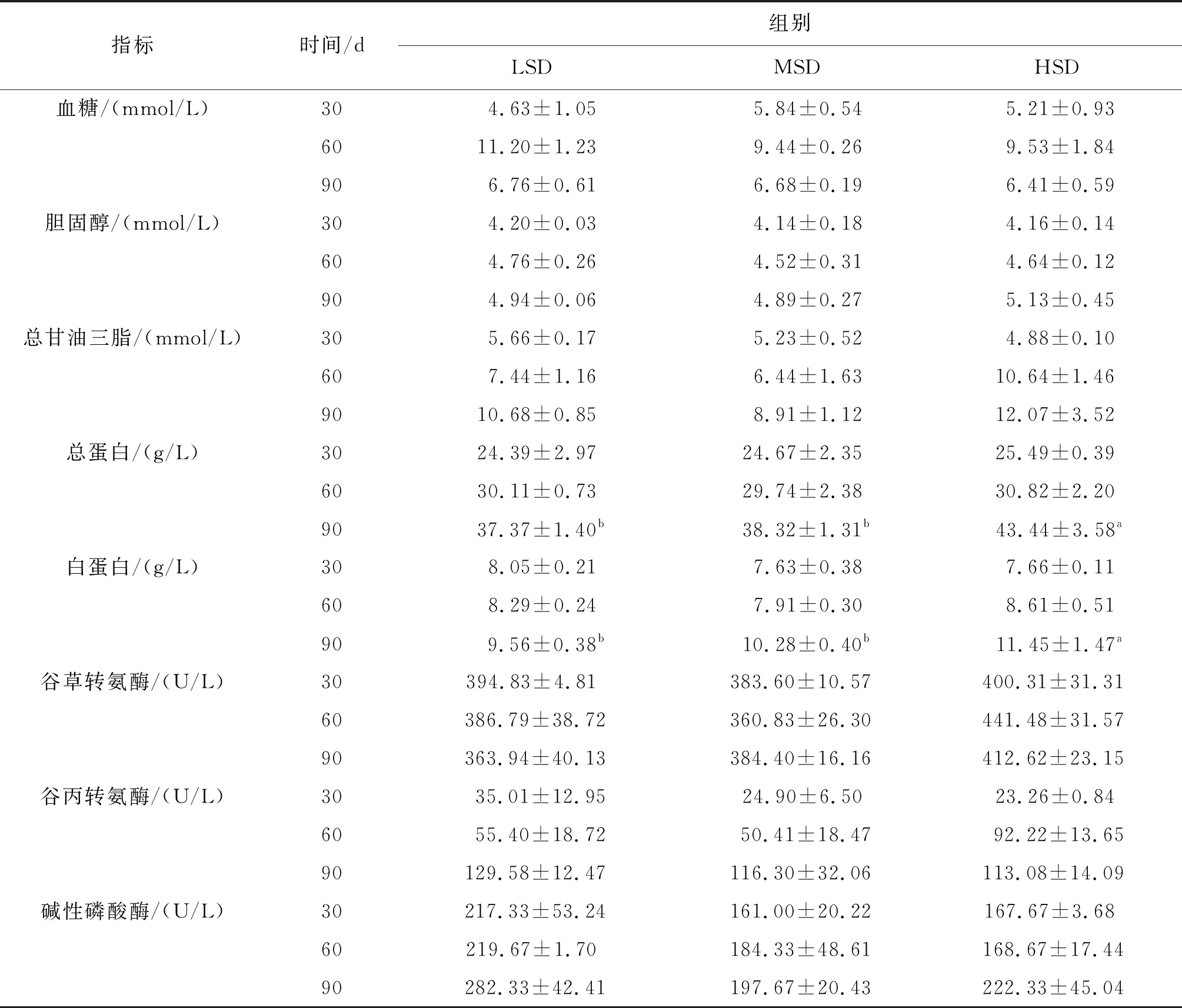
表2 各密度组长吻苗种血清生化指标Tab.2 Serum biochemical indexes of the juveniles of L.longirostris under different densities
指标时间/d组别LSDMSDHSD血糖/(mmol/L)304.63±1.055.84±0.545.21±0.936011.20±1.239.44±0.269.53±1.84906.76±0.616.68±0.196.41±0.59胆固醇/(mmol/L)304.20±0.034.14±0.184.16±0.14604.76±0.264.52±0.314.64±0.12904.94±0.064.89±0.275.13±0.45总甘油三脂/(mmol/L)305.66±0.175.23±0.524.88±0.10607.44±1.166.44±1.6310.64±1.469010.68±0.858.91±1.1212.07±3.52总蛋白/(g/L)3024.39±2.9724.67±2.3525.49±0.396030.11±0.7329.74±2.3830.82±2.209037.37±1.40b38.32±1.31b43.44±3.58a白蛋白/(g/L)308.05±0.217.63±0.387.66±0.11608.29±0.247.91±0.308.61±0.51909.56±0.38b10.28±0.40b11.45±1.47a谷草转氨酶/(U/L)30394.83±4.81383.60±10.57400.31±31.3160386.79±38.72360.83±26.30441.48±31.5790363.94±40.13384.40±16.16412.62±23.15谷丙转氨酶/(U/L)3035.01±12.9524.90±6.5023.26±0.846055.40±18.7250.41±18.4792.22±13.6590129.58±12.47116.30±32.06113.08±14.09碱性磷酸酶/(U/L)30217.33±53.24161.00±20.22167.67±3.6860219.67±1.70184.33±48.61168.67±17.4490282.33±42.41197.67±20.43222.33±45.04
2.3 养殖密度对长吻苗种血清抗氧化及免疫指标的影响
血清超氧化物歧化酶、过氧化氢酶含量变化范围较小,养殖密度对其影响也不显著;丙二醛含量随着养殖时间推进呈现先升高后降低的趋势,90 d时,HSD组丙二醛含量显著高于LSD、MSD组。溶菌酶含量随着时间推进先升高后降低,60 d时溶菌酶含量约2倍于90 d,但各密度组间含量差异不显著。补体3、补体4含量整体上随时间呈升高趋势,且随着密度的增大而升高,但各密度组间差异不显著(表3)。
表3 各密度组长吻苗种血清抗氧化及免疫指标Tab.3 Antioxidant and immune indexes in serum of the juveniles of L.longirostris under different densities
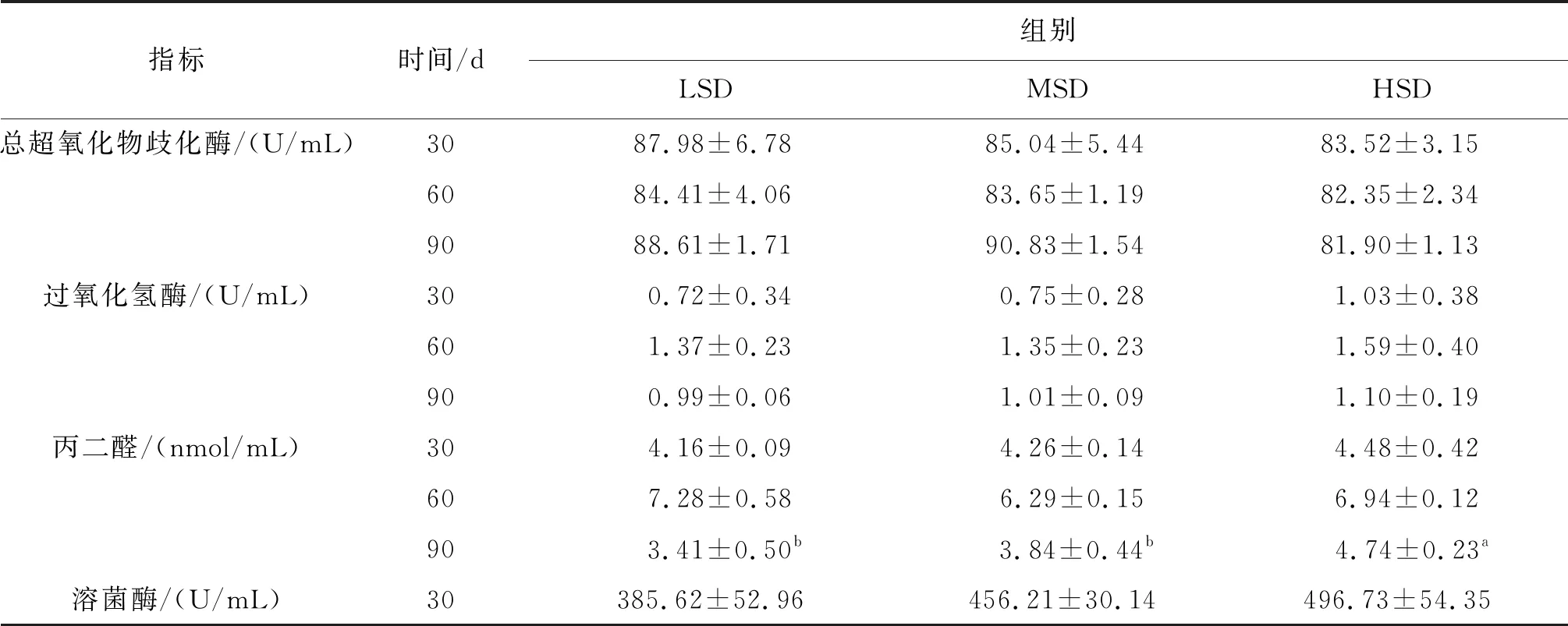
表3 各密度组长吻苗种血清抗氧化及免疫指标Tab.3 Antioxidant and immune indexes in serum of the juveniles of L.longirostris under different densities
指标时间/d组别LSDMSDHSD总超氧化物歧化酶/(U/mL)3087.98±6.7885.04±5.4483.52±3.156084.41±4.0683.65±1.1982.35±2.349088.61±1.7190.83±1.5481.90±1.13过氧化氢酶/(U/mL)300.72±0.340.75±0.281.03±0.38601.37±0.231.35±0.231.59±0.40900.99±0.061.01±0.091.10±0.19丙二醛/(nmol/mL)304.16±0.094.26±0.144.48±0.42607.28±0.586.29±0.156.94±0.12903.41±0.50b3.84±0.44b4.74±0.23a溶菌酶/(U/mL)30385.62±52.96456.21±30.14496.73±54.35
续表3
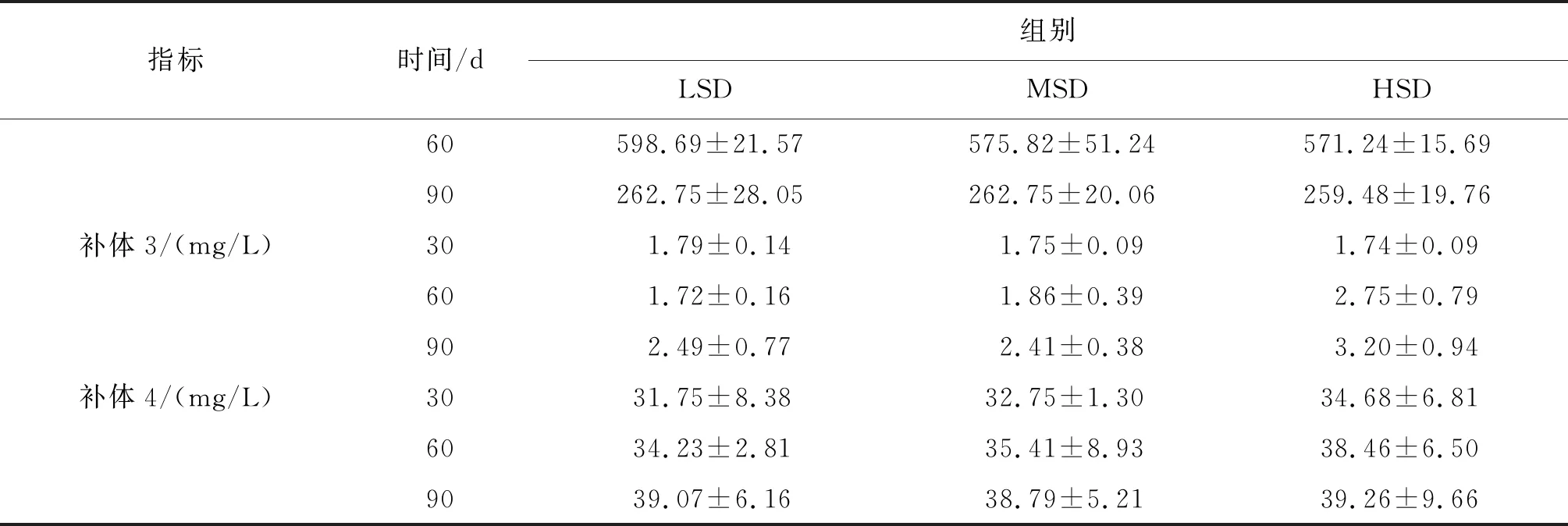
表3 各密度组长吻苗种血清抗氧化及免疫指标Tab.3 Antioxidant and immune indexes in serum of the juveniles of L.longirostris under different densities
指标时间/d组别LSDMSDHSD60598.69±21.57575.82±51.24571.24±15.6990262.75±28.05262.75±20.06259.48±19.76补体3/(mg/L)301.79±0.141.75±0.091.74±0.09601.72±0.161.86±0.392.75±0.79902.49±0.772.41±0.383.20±0.94补体4/(mg/L)3031.75±8.3832.75±1.3034.68±6.816034.23±2.8135.41±8.9338.46±6.509039.07±6.1638.79±5.2139.26±9.66
3 讨论
3.1 养殖密度对长吻苗种生长性能的影响
养殖密度升高可导致鱼类的生长性能降低,养殖密度越高对鱼类生长的抑制作用越强[4]。有学者认为,养殖鱼类为适应高密度的养殖环境,机体功能产生一系列的调节活动,一系列的调节活动需消耗大量能量,肝脏中的糖原等能源物质被大量分解,糖异生作用加强产生大量能量,导致用于生长的能量大幅减少抑制其生长[5]。在本试验中,整体上长吻苗种体质量和体长随着养殖时间进行而逐渐增大,且随着密度的增大而逐渐降低,45 d之前(体质量<50 g),各密度间差异不显著,45 d之后(体质量>50 g),LSD组显著大于MSD、HSD组,MSD和HSD差异不显著。增重率和特定生长率也与养殖密度呈负相关性,LSD组显著大于MSD、HSD组。有学者认为多数鱼类也存在社会行为,养殖密度是造成鱼类表现社会行为的关键原因,过高或过低的养殖密度使鱼类产生慢性胁迫,造成个体差异增加[6-8]。各密度组长吻苗种的体质量变异系数随养殖密度升高表现为负相关性,说明低密度组长吻苗种个体差异更明显。各密度组长吻苗种的摄食率和饲料系数均随养殖密度的升高而升高,该结果表明养殖密度的增大降低了养殖鱼类对饵料吸收和利用的效率;各密度组的苗种一直保持良好的食欲,且在短时间内吃完饵料,但高密度组的饲料系数大于低密度组,表明同样进食了相等量的饵料,但LSD组可以更好吸收和利用饲料。肥满度是一项常用的形体指标,反映的是鱼类生长情况、能量储存状态和鱼体肥瘦程度,肥满度的下降通常被解释为能量储备的消耗,如储存的肝糖原或体脂肪被消耗[9,10]。在本研究中,随着养殖密度的升高肥满度未表现出变化趋势,说明本实验设置的养殖密度未对长吻苗种的形体指标造成影响。各密度组死亡量相近,平均成活率均在99.11%以上,各密度组间差异不显著,说明长吻苗种可以较好地适应本实验的养殖密度范围,高密度组的养殖密度也并未达到该规格下长吻临界养殖密度。从生长性能方面看,为保证较高的生长速度,建议在养殖中后期(体质量约50 g)时降低养殖密度,控制在2 000~3 000 尾/圈为宜。
3.2 养殖密度对长吻苗种血清生化指标的影响
血液承担着体内多种物质的运输,同时也是机体代谢的主要调节途径,健康的血液也是鱼体行驶正常生理功能的前提条件,血液生化指标的变化一定程度上指示了机体组织器官的健康状态[11-13]。血糖、血脂及血液蛋白质均可作为能源物质为机体提供能量,其三者在不同情况时可以相互转化,被机体利用的顺序也因种类不同而变化[14,15]。在本研究中,养殖密度对血糖、血脂影响不显著,而血清总蛋白含量随着养殖密度的增大而显著升高,白蛋白含量变化趋势也与总蛋白相同,血清中的白蛋白对维持血液的渗透压平衡起着重要作用,是血液中含量最高的一种蛋白质,也是机体重要的免疫因子,抵御入侵机体的病原体和异物,也有修复受损组织的重要作用[16]。因此,持续作用的密度胁迫对长吻苗种产生了一定不利影响,高养殖密度胁迫作用对长吻苗种的组织器官造成了损伤,导致白蛋白得到相应的升高。血清酶的含量和活力变化趋势反映了机体物质转化和代谢情况,可以反映鱼体生理状态和健康状况[17]。在正常条件下,转氨酶主要存在于肝脏等的细胞内,一般血液中含量较低,当肝细胞损伤时,就被大量释放到血液中,导致其含量显著升高,因此两种转氨酶含量的高低指示了肝脏的健康状态及蛋白质的合成和代谢正常与否[18]。在本研究中,谷丙转氨酶含量随着养殖密度的增大而呈升高趋势,随着养殖时间进行谷草转氨酶含量变化不大势,不同时期各密度组间差异不显著,鱼体应对养殖密度的不同发生相应的调整机制,高密度组肝脏细胞可能受到一定程度的损伤。碱性磷酸酶通常是一种肝脏功能和骨损伤的标志酶,在肝脏损伤或者功能障碍时可引起其含量升高。同时,碱性磷酸酶也直接参与机体新陈代谢中磷酸基团的代谢和转移过程,在机体的物质代谢和免疫系统中发挥着关键功能[19]。在本研究中,碱性磷酸酶含量随着养殖时间而略有升高趋势,表明养殖后期高密度条件对机体肝脏功能产生了一定负面影响。从血清生化指标方面看,建议在养殖后期高密度组应降低养殖密度,控制在2 000~3 000 尾/圈为宜。
3.3 养殖密度对长吻苗种血清抗氧化及免疫指标的影响
高密度条件下鱼体长期处于不利环境胁迫中,此时机体会产生大量的强氧化性的活性氧自由基,它是鱼体内的重要的活性物质,完成体内各种生理生化的调节活动,但过量的氧自由基机体并不能及时清除,造成相应器官及机体的氧化损伤、机体对不良环境抵抗力降低,而活性氧自由基的消除则主要依赖于抗氧化系统发挥作用[20,21]。超氧化物歧化酶和过氧化氢酶是抗氧化物酶中两种关键酶[22],超氧化物歧化酶是去除活性氧自由基的关键酶,主要将细胞内氧自由基转化为过氧化氢和氧分子[23]。过氧化氢酶可将超氧化物歧化酶催化产生的过氧化氢催化分解为氧分子和水分子以降低过氧化氢的毒性[24]。在本试验中,随着养殖密度增加长吻苗种超氧化物歧化酶含量没有显著差异,高密度组过氧化氢酶含量在整个养殖周期均略高于中低密度组且差异不显著,表明该条件下中低密度下长吻苗种的抗氧化系统处于正常状态,机体没有受到活性氧自由基的侵害,高密度组过氧化氢酶活性升高表明氧自由基浓度升高,相应细胞组织一定程度上被氧化毒害,脂质过氧化产物丙二醛含量也有所反映,丙二醛含量随着养殖密度先升高后降低,30、60 d时各密度组间差异不显著,90 d时高密度组丙二醛含量显著大于低密度组,表明高密度胁迫使鱼体内活性氧自由基含量增多,脂质过氧化反应增强,过量的自由基并没有及时被清除,氧自由基就会直接攻击细胞膜中的不饱和脂肪酸发生脂质过氧化反应,其分解反应产物则为丙二醛,因此丙二醛是用来评价细胞氧化损伤及肝脏和胰腺损伤程度的重要指标。
鱼类非特异性免疫因子主要由溶菌酶、补体及干扰素等构成[25,26]。溶菌酶是存在于许多鱼类的体表黏液、肠道黏液、巨噬细胞和血清中的水解酶,是一种碱性蛋白质[27],其通过破坏细菌内外渗透压平衡导致细菌细胞膜破裂,从而达到免疫防御作用[28,29]。补体是鱼类抵抗外界不良生存环境的重要成分,具有溶菌、杀菌及免疫吸附等的作用[26,30],补体3是补体激活旁路中的关键成分,补体4在经典途径中发挥重要作用,两者含量可以一定程度上显示机体免疫病理性受损的情况[31,32]。本研究中,溶菌酶含量呈现先升高后降低趋势,补体3、补体4含量均是呈升高趋势,且各密度间差异不显著,表明养殖密度对溶菌酶、补体3、补体4水平影响较小,对非特异性免疫系统影响不显著,长吻苗种可以通过生理调节适应拥挤胁迫。从血清抗氧化指标方面看,建议在养殖后期高密度组应降低养殖密度,控制在2 000~3 000尾/圈为宜。
